
Abstract
Recent advancements in cancer research have led to major breakthroughs; however, the impact on overall cancer-related death rate remains unacceptable. Thus, further insights into tumor markers and subsequent development of targeted therapies are urgently needed. For decades the urokinase plasminogen activator (uPA) system has been thought to drive tumor progression by mediating directed extracellular proteolysis on the surface of migrating or invading cells. Intervention with this proteolysis by targeting of uPA receptor (uPAR) has been proposed to represent a novel approach for inhibiting tumor progression. Recent data have provided new insights into the role of uPAR in gastric cancer progression. In addition to mediating proteolysis, this receptor also appears to mediate cell signaling, proliferation, and survival, and these observations have revealed novel ways to target uPAR. In this review, we discuss uPAR expression in gastric cancer, the relationship between uPAR and Helicobacter pylori, and recent insights into uPAR-signaling mechanisms. The role of uPAR as a cancer target in gastric cancer is also summarized.
Introduction
Gastric cancer is the second most common carcinoma causing death worldwide. 1 The mechanisms of this clinically intractable problem have yet to be clearly elucidated. The factors explaining these variations remain unknown so far, given the complexity and multifactorial nature of the disease. Among the key events for cancer development and progression are tumor cell invasion into the adjacent normal tissue, and eventually metastasis. Because of local tissue invasion and metastasis, radio- or chemotherapy do not significantly enhance the length or quality of life of patients with advanced gastric cancers. An understanding of the detailed mechanisms of invasion and metastasis in gastric cancer would therefore be helpful in developing improved treatments. Extracellular proteolytic enzymes, such as serine proteinases and metalloproteinases, have been implicated in cancer metastasis. Degradation of the extracellular matrix (ECM) in the tissue surrounding the tumor is a critical event in the process of cancer invasion and metastasis. 2 The basic idea is that the release of proteolytic enzymes in tumors facilitates cancer cell invasion into the surrounding normal tissue by breaking down the basement membranes and ECM. 3 –5
The urokinase plasminogen activator (uPA) system is thought to play a role in several different processes important to tumor progression, including angiogenesis, 6 tumor growth, 7 and metastasis. 8 This system is composed of a serine protease, uPA, its glycolipid (glycosylphosphatidylinositol) anchored receptor (uPAR), and several serine protease inhibitors (serpins), including plasminogen activator inhibitors 1 and 2. uPA has the ability to convert plasminogen to active plasmin, and activates matrix metalloproteinases (MMPs) and growth factors, 9 which may play important roles in pericellular proteolysis and angiogenesis. 10 It binds to uPAR on the surface of the tumor cell, and it is this binding that elicits the activation of plasminogen into plasmin, which converts ECM to degradation products and also activates many intracellular signaling pathways (Fig. 1). Because uPA is secreted from tumor cells as well as normal fibroblasts, uPAR expression may better reflect tumor cell malignancy than the expression of uPA. 11 uPAR is predominantly seen on inflammatory cells and cancer cells in areas of cancer invasion in several types of cancer, including gastric, colon, breast, and oral cancer, 12 –17 suggesting that it is involved in both tumor cell invasion and metastasis. In gastric cancer lesions, uPAR expression has also been observed in macrophages, endothelial cells, and cancer cells located at the invasive front of the tumors. 12,15,16,18 uPAR can function in various pathways in the development of gastric cancer. Identification of uPAR signals provides a new pathway for anticancer treatment. Although this target has been known for many years, attempts at developing a therapeutic targeting uPAR have never advanced past the in vivo proof-of-principle stage. The mechanisms of uPAR signaling are still only partially understood, especially in gastric cancer. In this review, we concentrate on recent insights into uPAR-signaling mechanisms. We begin with an overview of uPAR expression in gastric cancer. The relationship between uPAR and Helicobacter pylori is then discussed. We also summarize the various mechanisms of uPAR signaling associated with gastric cancer.
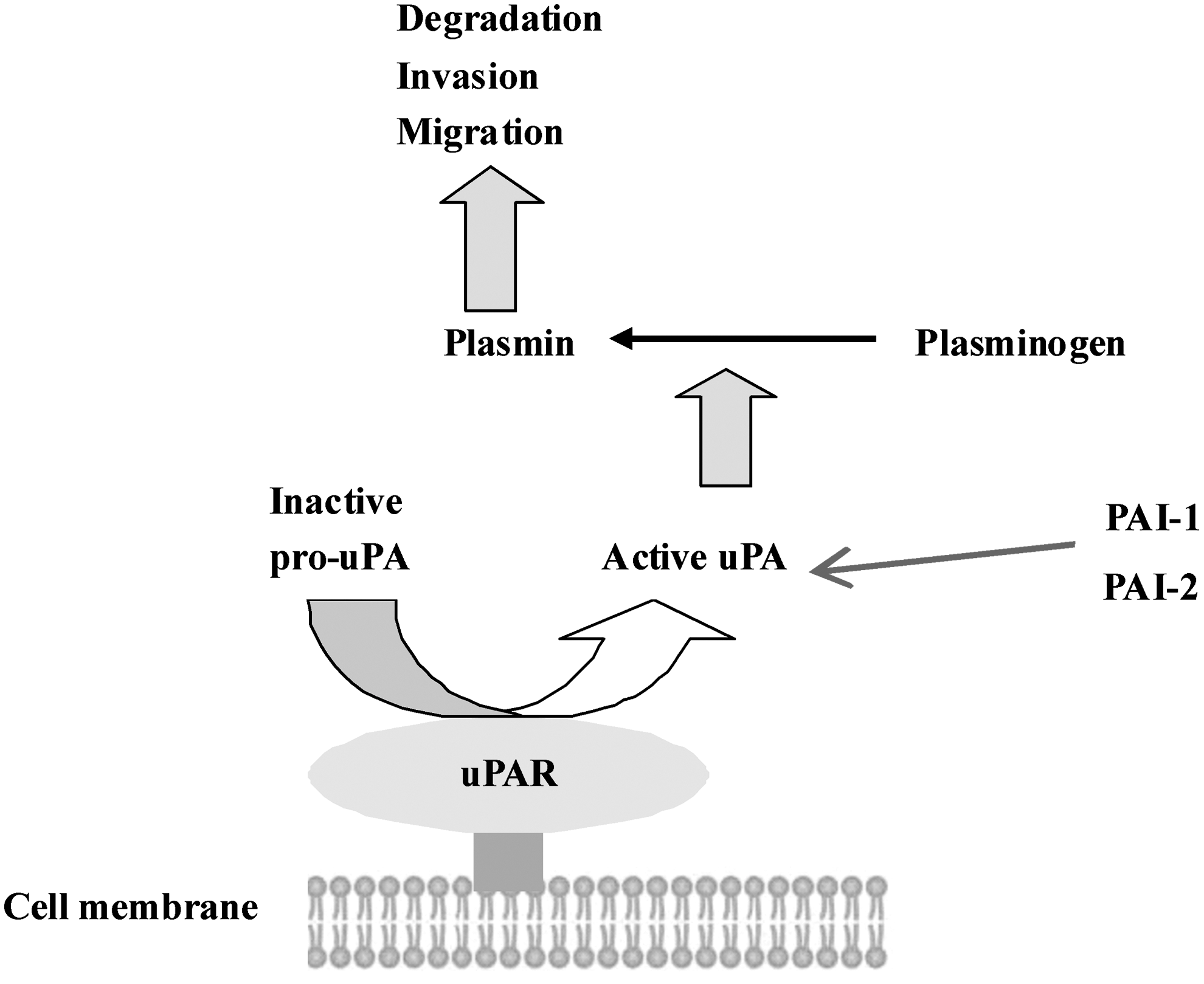
Schematic representation of the role of u-PAR at the cell surface. The urokinase receptor (uPAR) converts inactive urokinase (pro-uPA) to active uPA. Active uPA proteolytically activates plasminogen, resulting in active plasmin. Plasmin is able to degrade the extracellular matrix to induce invasion and migration. uPA can be inactivated by the uPA-specific plasminogen-activator inhibitor 1 or 2 (PAI-1 or PAI-2). uPAR, urokinase plasminogen activator receptor.
uPAR Expression in Gastric Cancer
Many malignant cultured cell lines and human neoplasms have been characterized by their increased uPAR expression, 19,20 thus suggesting that the inhibition of uPAR activity could be a promising strategy to prevent cancer invasion and metastasis. Choi et al. 21 demonstrated that overexpression of uPAR in human gastric carcinomas correlated closely with their invasiveness and tumorigenicity. A significant association between high expression levels of uPAR in primary tumors and uPAR-positive disseminated tumor cells was also reported. 22 These observations suggested that elevated expression of uPAR in tumors, and particularly in malignant cells, may be indicative of more aggressive gastric cancers.
Increased expression of uPAR antigen or mRNA in tissue extracts and blood from gastric cancer patients has been associated with some clinicopathological aspects of the disease, including poor prognosis. 18,23,24 Kita et al. 25 assessed the clinicopathologic significance of uPAR expression in the bone marrow and peripheral blood of 846 gastric cancer patients, and found that uPAR mRNA expression was significantly higher than in normal controls. They also determined that uPAR expression in peripheral blood can be a preferential tumor marker to predict disease-free survival in gastric cancer outpatients. Zhang et al. 18 used in situ hybridization and immunohistochemical techniques to study the expression of uPA, uPAR, vascular endothelial growth factor (VEGF), and CD34 protein in 105 gastric carcinoma specimens, and found that uPA and uPAR expression correlated with enhanced VEGF-induced tumor angiogenesis, and may play a role in invasion and nodal metastasis of gastric carcinoma, thereby serving as prognostic markers of gastric cancer. Kaneko et al. 23 also found that uPAR expression was correlated with tumor size, depth, lymph node involvement, differentiation, vascular invasion, and survival rate. By univariate analysis, a significant association of MVD with uPAR was identified, suggesting that uPAR may contribute to tumor invasion.
The aforementioned results showed that uPAR is involved in invasion through cell surface-associated plasminogen activation and that its presence on gastric cancer cells is linked to micrometastasis and poor prognosis.
Relationship Between uPAR and
H. pylori
The majority of gastric cancers are end products of an inflammatory cascade. H. pylori infection is now accepted as the major cause of chronic gastritis. This inflammatory cascade is caused by infection with the bacterial pathogen H. pylori, which is a necessary, but not sufficient, cause of gastric cancer. Current data support the notion that gastric cancer related to H. pylori can largely be prevented if the infection is eradicated in the preatrophic stage. However, after the development of atrophic gastritis, H. pylori eradication can reduce the risk of cancer, but does not completely prevent it. 26 –29 Several epidemiological studies have shown that H. pylori infection is linked to severe gastritis-associated diseases, including peptic ulcers and gastric cancer. 30 In addition, studies in gastric cancer cell lines have reported increased uPAR mRNA levels when neoplastic cells are cocultured with H. pylori, 31,32 suggesting that H. pylori may induce the expression of uPAR in gastric mucosa. This may link uPAR with the early stages of gastric carcinogenesis, which contributes to the explanation of the association between H. pylori and gastric cancer, and the correlation observed between the incidence rates of this malignancy and the prevalence of the bacterium. 33
It has been reported that the mechanism by which H. pylori induces gastric cancer involves the induction of chronic inflammation in the gastric mucosa, and during progression through to precancerous lesions (atrophic gastritis, metaplasia, and dysplasia) H. pylori induces mutation of genes 34 related to gastric cancer angiogenesis and metastasis, including interleukin-8, 35 VEGF, 36 and MMPs. 37 Semiquantitative analysis of H. pylori–positive cases for the presence of uPAR-positive surface epithelium revealed a significant association, and double immunofluorescence analysis also identified H. pylori clusters in proximity to uPAR-positive surface epithelium. This led to the speculation that H. pylori can stimulate the expression of uPAR in epithelial cells, which under certain circumstances could potentiate the development of gastric pathological conditions that lead to gastric neoplasia. 38 Another study 39 showed that H. pylori stimulates the expression of uPA, its receptor uPAR, and its inhibitor PAI-1 in gastric epithelial cells. H. pylori stimulated epithelial cell proliferation, which was inhibited by uPA immunoneutralization and uPAR knockdown; this occurs through activation of heparin-binding epidermal growth factor, and is normally inhibited by concomitant induction of PAI-1. These findings raise new and exciting questions regarding the role of H. pylori and uPAR in the development of gastric cancer, including elucidation of the role of the various H. pylori virulence factors in the induction of uPAR and the potential prognostic value of uPAR in gastric cancer.
In vitro studies have shown increased levels of both uPAR mRNA and uPAR antigen when gastric cancer cell lines are challenged with H. pylori, 32,40 and when cocultured with H. pylori cytotoxin-associated gene A (cagA)–positive versus –negative strains. 31,41 Taken together, these observations suggest that H. pylori may be directly involved in the induction of uPAR in the gastric mucosa. Kim's study 32 also supports the idea that H. pylori can induce the uPAR expression and stimulate invasiveness of human gastric cancer AGS cells, suggesting that overexpression of uPAR promoted by H. pylori is involved in increased cell invasiveness. In summary, these reports have indicated the possibility that H. pylori infection may play a role in the process of destruction of the ECM.
uPAR is also associated with other molecules in the mucosal microenvironment, including mediators of inflammation released in response to H. pylori infection (Fig. 2). Iwamoto et al. 31 found that H. pylori infection may be involved in degradation of the ECM by inducing uPA and uPAR complex in cancer cells. They also demonstrated that MG132, an NF-κB inhibitor, downregulated H. pylori–induced uPA and uPAR expression in cancer cells, indicating that NF-κB is also a key factor in H. pylori–induced uPA and uPAR expression. Furthermore, Kim et al. 32 also reported the role of H. pylori in the regulation of uPAR in gastric cancer cells, and strongly suggested that ERK-1/2 and JNK triggered by H. pylori induce AP-1 activity, which subsequently upregulates uPAR expression, resulting in increased cell invasiveness in gastric cancer cells. Iwamoto et al. 41 demonstrated that uPA and uPAR expression levels in gastric cancer cells relatively increased with H. pylori stimulation, and that COX-2 mRNA expression level and the amount of PGE2 antigen increased with H. pylori stimulation: H. pylori–induced uPA and uPAR expression levels were suppressed with COX2 inhibitor and PGE2 receptor antagonist treatment, indicating that the COX2-PGE2 pathway is involved in H. pylori–associated uPA system induction. Understanding the mechanism by which uPAR expression is regulated by H. pylori may lead to the development of new strategies for treatment of gastric carcinomas; therefore, the possible connection between H. pylori and uPAR and its potential implications in gastric cancer development and progression need to be further explored.
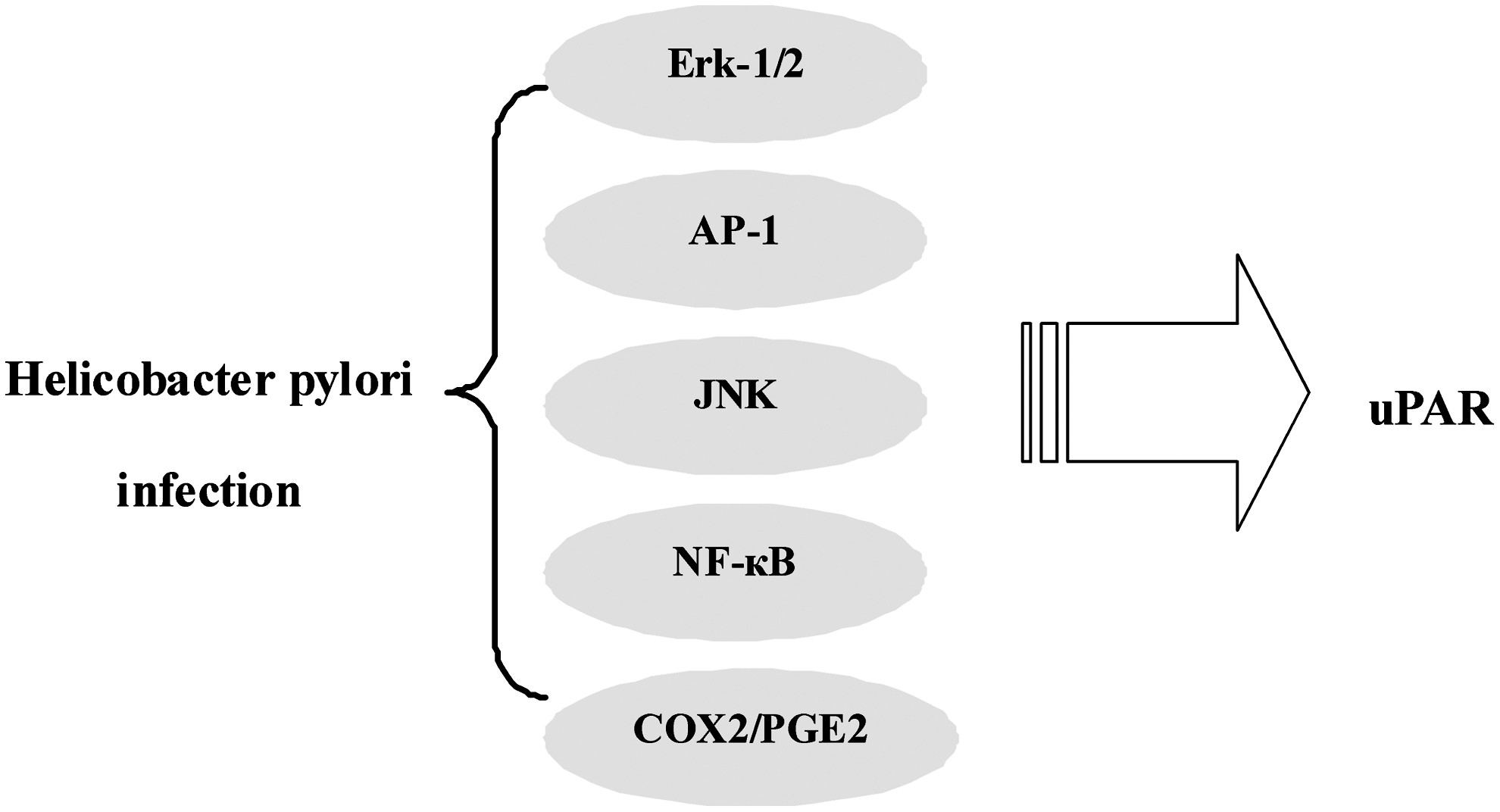
Schematic demonstrating the relationship between uPAR and Helicobacter pylori infection. Signals triggered by H. pylori induce uPAR expression, which subsequently results in increased invasiveness of gastric cancer cells.
uPAR-Mediated Signaling in Gastric Cancer
Interest in uPAR has intensified since its upregulated expression in human diseases, including many cancers, was first observed. 42 –44 Therapeutic targeting of uPAR to interrupt proteolytic cascades and block intracellular signaling has therefore become an attractive concept. 45
Recent reports revealed that signaling pathways triggered by other molecules induce uPAR expression, which subsequently result in increased invasiveness of gastric cancer cells (Fig. 3). Many types of carcinoma cells, including gastric cancers, express EGF and EGFR at high levels; these proteins have been implicated in tumor development and progression. 46 EGF has been shown to regulate tumor cell invasion through induction of many proteolytic enzymes, including MMPs and uPA systems. Baek et al. 47 also investigated the effect of EGF on uPAR expression and the underlying signaling pathways in human gastric cancer AGS cells. These researchers found that EGF induced uPAR mRNA expression in a time- and concentration-dependent manner, and promoted activation of ERK-1/2 and P38 mitogen-activated protein kinase (MAPK) in human gastric cancer cells, suggesting that EGF induces uPAR expression via ERK-1/2-, AP-1-, and NF-κB-signaling pathways and, in turn, stimulates cell invasiveness in human gastric cancer. Analysis of the genomic structure of uPAR also reveals many potential targets for regulation at both the transcriptional and post-transcriptional levels. Within its 50-flanking region, the uPAR gene contains putative binding sites for AP-1, AP-2, and Sp1. 48 Multiple lines of evidence demonstrate that macrophage-stimulating protein (MSP) and its receptor, recepteur d'Origine nantais (RON), are closely associated with cancer development and progression. MSP signaling induces epithelial-to-mesenchymal transition in immortalized cells, 49 and influences cancer cell migration and invasion. 50,51 RON and its splice variant are also reported to play important roles in cell invasion and metastasis of gastric cancer. 52 Park et al. 53 reported the role of MSP and RON in the regulation of uPAR and invasiveness in gastric cancer cells, and found that MSP induced uPAR mRNA expression and promoter activity. The introduction of siRNA-RON significantly affected MSP-induced uPAR transcription. Deletion and site-directed mutagenesis studies and expression vectors studies also confirmed that NF-κB and AP-1 were essential for MSP-induced uPAR expression. These above results strongly support the suggestion that MAPK, NF-κB, and AP-1, triggered by MSP, induce uPAR expression, which subsequently increases gastric cancer cell invasion.
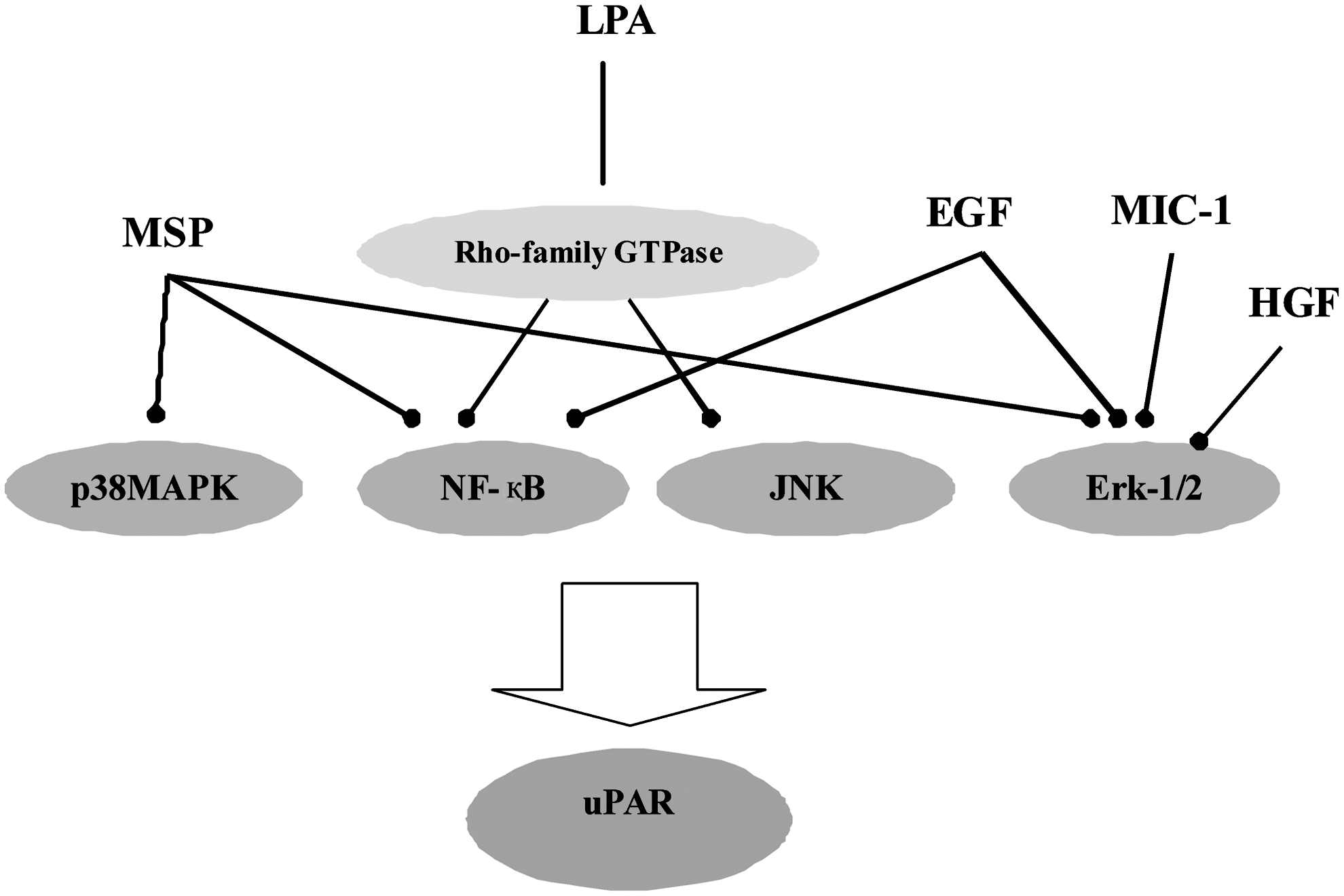
uPAR-mediated signaling in gastric cancer.
Macrophage inhibitory cytokine-1 (MIC-1) is the first member of a divergent group within the TGF-β superfamily. 54,55 Recent findings suggest the role of MIC-1 in tumor development or progression. 56 –58 Lee et al. 59 found specific expression of MIC-1 in gastric tumor tissues and gastric cancer cells, and this expression level was correlated with invasive potential in various human gastric cancer cell lines. Stable transfection of MIC-1 significantly increased invasiveness and the activity of uPA, and uPA and uPAR expression. Similarly, dose-dependent stimulation of gastric cancer cell lines with purified recombinant MIC-1 increased cell invasiveness and uPAR expression. Their study also indicated that MIC-1 may contribute to invasion of gastric cancer cells through the upregulation of uPAR via an ERK-1/2-dependent pathway. In vitro experiments were conducted in another study 60 to determine the function of HGF/c-Met signaling and its direct effects on the growth and invasion of gastric cancer cell lines. Treatment with HGF increased c-Met phosphorylation in a dose-dependent manner, as well as increasing cell proliferation. An antibody against human uPAR inhibited HGF-mediated tumor cell invasion, which may provide a therapeutic target by inhibiting uPA- and uPAR-mediated proteolysis.
Lysophosphatidic acid (LPA), which is produced through the action of lysophospholipids from inflammatory cells and platelets, 61 is a bioactive phospholipid that is present in serum and induced by various genes related to cancer angiogenesis and metastasis. 62 Kim et al. 63 found that LPA enhances the expression of uPAR and cell invasiveness in human gastric cancer AGS cells through the LPA receptors, Rho-family GTPase, JNK-, AP-1-, and NF-κB-signaling pathways, which in turn stimulate cell invasiveness. Therefore, interruption of proteolytic cascades and block intracellular signaling by targeting uPAR may lead to the development of new strategies for the treatment of gastric carcinomas.
Conclusions and Perspectives
The main objective of cancer therapy is to arrest tumor growth and manage it as a controlled, localized disease. Cellular functions regulated by uPAR, a multifunctional protein, have been shown to be crucial in a variety of physical and pathological processes in vivo, including inflammation, wound healing and fibrogenesis, neoangiogenesis, tumor growth and metastasis, and tissue destruction and regeneration. 64 –67
Accumulated lines of evidence indicate that uPAR plays an essential role in tumor invasion and metastasis. A large body of published evidence suggests that targeting of uPAR could have broad-spectrum antitumor effects. Although anti-uPAR therapeutic agents are yet to enter clinical trials, uPAR presents multiple opportunities for targeted therapies that might be beneficial in gastric cancer. Many advances have been made since the discovery that uPAR regulates cell signaling independently of uPA-mediated proteolysis. Thus, we presume that uPAR plays a major role in tumor progression and that targeting uPAR will lead to clinical benefit in cancer patients. However, the mechanisms of uPAR signaling are still only partially understood, and the exact role of uPAR in these processes is not fully known and requires further study.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (No. 81071991) and Project of Science and Technology of Zhejiang Province (2010C33018).
Disclosure Statement
No competing financial interests exist.