
Abstract
Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) has been utilized for cancer therapy, but the resistance of cancer cells to TRAIL remains an obstacle. Ether à go-go 1 (Eag1) channel is overexpressed in a variety of cancers and implicated in tumor progression. However, the therapeutic potential of Eag1 in osteosarcoma remains elusive. In this study, we generated CRAd5.TRAIL/siEag1 adenoviral vector that permitted simultaneous knockdown of Eag1 and overexpression of TRAIL and investigated its antitumor effects on human osteosarcoma MG-63 cells. Our results showed that CRAd5.TRAIL/siEag1 induced growth arrest and apoptosis of MG-63 cells in a more efficient manner than CRAd5.TRAIL or CRAd5.siEag1, and had no effect on human osteoblastic hFOB 1.19 cells. Furthermore, treatment of an osteosarcoma xenograft model with CRAd5.TRAIL/siEag1 resulted in significant tumor regression and cancer cell apoptosis, compared with treatment with CRAd5.TRAIL or CRAd5.siEag1. Taken together, our results demonstrate that CRAd5.TRAIL/siEag1 may represent an effective strategy for osteosarcoma gene therapy due to the synergistic antitumor effects of Eag1 knockdown and TRAIL overexpression.
Introduction
Osteosarcoma is the most common primary bone tumor in childhood and adolescence. 1–2 About 60% of osteosarcoma cases occur in people ranging from 10 to 20 years old, and about 80% of them appear in the long bones of the body, such as the femur, the tibia, or the humerus. 3 Osteosarcoma derives from primitive bone-forming mesenchymal stem cells and has high metastatic potential. 4 The main sites of metastases of osteosarcoma are the lungs, pleura, and the heart. 5 With the use of combination aggressive surgery and neoadjuvant chemotherapy, 5-year survival rates of osteosarcoma patients without metastatic disease have reached 60%–75%. 6 However, 40%–50% of patients will develop metastases, and the prognosis is poor for them, especially for patients with primary pulmonary metastases. 7 Meanwhile, high-dose chemotherapy has adverse reactions such as gastrointestinal reactions, bone marrow suppression, cardiac toxicity, and renal toxicity, which restrict the application of chemotherapy. Thus, further treatment improvements for osteosarcoma are urgently needed. Several genetic alterations have been found to be associated with osteosarcoma, suggesting the potential application of biological therapy in osteosarcoma treatment.
An increasing number of ion channels have been involved in tumor progression, among which Ether à go-go 1 (Eag1, Kv10.1) channel attracts the most attention. Eag1 is a central nervous system-localized voltage-gated potassium channel that is ectopically expressed in more than 75% of extracranial solid tumors, and its overexpression is associated with poor prognosis in several tumor types. 8 –11 Furthermore, inhibition of Eag1 by specific small interference RNA (siRNA) or nonspecific blockers could decrease tumor progression both in vitro and in vivo. 10 –13 Although a clear relationship between Eag1 and tumor progression has been established, so far little attention has been drawn to the relationship between Eag1 and osteosarcoma, and the therapeutic potential of Eag1 for osteosarcoma.
Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) is a recently identified member of the tumor necrosis factor (TNF) gene superfamily 14 and is a new candidate for cancer therapeutics owing to its ability to selectively induce apoptosis in tumor cells, but not in normal cells. 15 Many experimental studies of the application of TRAIL to tumor cells have been reported, including osteosarcoma. 16 –20 However, the instability of TRAIL, the resistance of some cancer cells to TRAIL, and a requirement for large quantities of TRAIL to suppress tumor growth in vivo present the main obstacles for clinical experimentation. 21,22 Therefore, a direct gene transfer of TRAIL to malignant cells is a part of an alternative delivery strategy. 23
In this study, we employed the human osteosarcoma line MG63 as the experimental model and constructed a recombinant adenovirus CRAd5.TRAIL/siEag1, which could induce the downregulation of Eag1 and overexpression of TRAIL. We investigated the therapeutic potential of CRAd5.TRAIL/siEag1 and explored the cooperative antitumor effects for osteosarcoma. Our results showed obvious antitumor activity of CRAd5.TRAIL/siEag1 both in vitro and in vivo.
Materials and Methods
Cell culture
Human osteosarcoma cell line MG-63, human embryonic kidney cell line 293 (HEK293), and human osteoblastic cell line hFOB 1.19 were purchased from the American Type Culture Collection (ATCC, Rockville, MD). MG-63 and HEK293 cells were cultured in an RPMI-1640 medium (Gibco, Rockville, MD) supplemented with 10% fetal bovine serum (FBS; Gibco), 100 IU/mL penicillin, and 100 μg/mL streptomycin in a humidified atmosphere of 5% CO2 at 37°C. hFOB 1.19 cells were cultured in ham's F12/Dulbecco's modified Eagle's medium (DMEM; Gibco) medium supplemented with 10% FBS, 100 IU/mL penicillin, and 100 μg/mL streptomycin in a humidified atmosphere of 5% CO2 at 33.5°C. All cells were subcultured every 3–4 days.
Construction of gene silencing and adenoviral vectors
The human TRAIL cDNA was amplified by PCR and cloned into pcDNA (Invitrogen) to make pcDNA-CMV-TRAIL. An hU6-adapted Eag1 shRNA construct was designed with the following sequence: 5′-AGCCATCTTGGTC CCTTATAA-3′. The hU6-shEag cassette was inserted into pcDNA-CMV-TRAIL, named pcDNA-CMV-TRAIL-hU6-shEag1, and the CMV-TRAIL-hU6-shEag1 fragment was subcloned into the pAdeno-X backbone to make a recombinant pCRAd5.TRAIL/siEag1 plasmid. According to the previous method, 24 the pAdeno-X backbone was constructed. As the controls, CRAd5.TRAIL, CRAd5.siEag1, and Ad5.eGFP plasmids were constructed as well, and particle titers of adenovirus preparations were determined using an absorbance at 260 nm and plaque assays in HEK 293 cells.
Methyl thiazolyl tetrazoliym assay
The cell proliferation was analyzed by using methyl thiazolyl tetrazoliym (MTT) assay according to the manufacturer's protocol. Briefly, 20 μL of cell suspension (1×105 cells/mL) was distributed into each well of flat-bottomed 96-well culture plates (Sigma), and each plate was incubated for 24 hours with 5% CO2 at 37°C. Then, the cells were treated with various adenovirus constructs. After 0, 12, 24, 36, 48, and 72 hours, the cells were harvested, respectively. Thereafter, 20 μL of MTT dye (5 mg/mL; Sigma) was added into each well. Four hours later, the media were carefully discarded, and 150 μL of dimethyl sulfoxide (Shanghai Sangon) was added into each well. The absorbance (A) values of each well at 490 nm was read using an automatic multiwell spectrophotometer (Bio-Rad). Experiments were performed at least three times with representative data presented.
Apoptosis assay
Apoptosis was determined using the Apo-ONE homogeneous caspase-3/7 assay (Promega) according to the manufacturer's instructions. Briefly, 20 μL of cell suspension (1×105 cells/mL) was seeded into 96-well plates, and each plate was incubated overnight with 5% CO2 at 37°C. After the cells were treated with various adenovirus constructs, an equal volume of Apo-ONE Caspase-3/7-reagent was added to a 96-well plate and incubated at room temperature for 1 hour. The luminescence of each sample was measured in a plate-reading luminometer (Tecan). Experiments were performed at least three times with representative data presented.
Western blot analysis
About 5–6×107 cells were collected and lysed in an ice-cold lysis buffer containing 50 mM Tris–Cl (pH 7.5), 150 mM NaCl, 0.2 mM EDTA, 1 mM PMSF, and 1% Nonidet-P40 for 30 minutes. The lysates were centrifuged at 13,200 rpm for 10 minutes at 4°C, and the supernatants were collected. About 25 μg protein was resolved by a 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and blotted on nitrocellulose membranes (Bio-Rad). Membranes were blocked with 10% nonfat milk or dry milk at room temperature for 1 hour, and then incubated with antibodies against Eag1 (Alomone laboratories), TRAIL (Abcam), poly(ADP-ribose) polymerase (PARP), cleaved caspase-3, B-cell lymphoma 2 (Bcl-2), Bcl2-associated X protein (Bax; Cell Signaling), and GAPDH (Abcam) overnight, followed by incubation with horseradish peroxidase-conjugated goat anti-rabbit secondary antibody (Santa Cruz Biotechnology). Then, the membranes were developed with a chemiluminescent detection kit (Zhongshan Biotechnology) and exposed to X-ray films. Experiments were performed at least three times with representative data presented.
Tumor model
Thymus-null BALB/c nude mice (female, age 6–8 weeks) were obtained from the Animal Center of Chinese Academy of Medical Sciences. All animal procedures were performed according to the approved protocols and in accordance with the recommendations for the proper Use and Care Of Laboratory Animals. Osteosarcoma xenografts were established in nude mice according to a previous report. 25 A total of 1.5×106 MG-63 cells in 150 μL phosphate-buffered Saline (PBS) were subcutaneously injected in the hind right leg. One week later, the tumors grew to a visible size. The osteosarcoma-bearing mice were randomly divided into 4 groups (6 in each group) and received intratumor injections of Ad5.eGFP, CRAd5.siEag1, CRAd5.TRAIL, or CRAd5.TRAIL/siEag1 (1×107 pfu/50 μL), respectively. All treatments were given every other day for a total of seven doses. The tumor volume (cm3) was determined based on the following formula: ab2/2, where a was the length, and b was the width of the tumor. 25
TUNEL assay
Cell apoptosis of tumor sections was assessed using a TUNEL kit (Invitrogen, Rockville, MD, USA). Briefly, 14 days after the first intratumor injection, the nude mice were anesthetized with ether and killed, and then tumors of hind right leg were excised and fixed in 3.7% formaldehyde. Then, tumor sections were fixed with a freshly prepared paraformaldehyde solution (4% in PBS, pH 7.4) at room temperature for 30 minutes. The sections were washed with PBS and resuspended in a permeabilization solution (0.1% Triton X-100 in 0.1% sodium citrate) for 2 minutes on ice. After additional washing with PBS, cells were labeled using a TUNEL kit according to the manufacturer's instructions. The apoptotic index was calculated using the formula: apoptotic index=(total number of apoptotic cells/total number of cells)×100%.
Statistical analysis
All the data were presented as mean±standard deviation (SD). Statistical significance was determined using t-test or analysis of variance (ANOVA) using SPSS18.0 software. p<0.05 was considered as a statistically significant difference.
Results
Characterization of CRAd5.TRAIL/siEag1 vector
To achieve maximum efficiency of Eag1 silencing and TRAIL expression in human osteosarcoma cells, adenoviral vectors (Ad5.eGFP, CRAd5.siEag1, CRAd5.TRAIL, and CRAd5.TRAIL/siEag1) were generated. To compare the efficiency of siRNA-mediated silencing of Eag1 and overexpression of TRAIL, we infected MG63 cells with four adenoviral constructs, and the expression of Eag1 and TRAIL was measured. As shown in Figure 1, infection with CRAd5.siEag1 or CRAd5.TRAIL/siEag1 resulted in significant reduction in Eag1 expression. In contrast, the expression of Eag1 remained unchanged in cells infected with Ad5.eGFP or CRAd5.TRAIL. On the other hand, the TRAIL expression level was obviously increased in the cells infected with CRAd5.TRAIL or CRAd5.TRAIL/siEag1, but not in those infected with Ad5.eGFP or CRAd5.siEag1. These results indicated that CRAd5.TRAIL/siEag1 could induce both efficient silencing of Eag1 and overexpression of TRAIL.
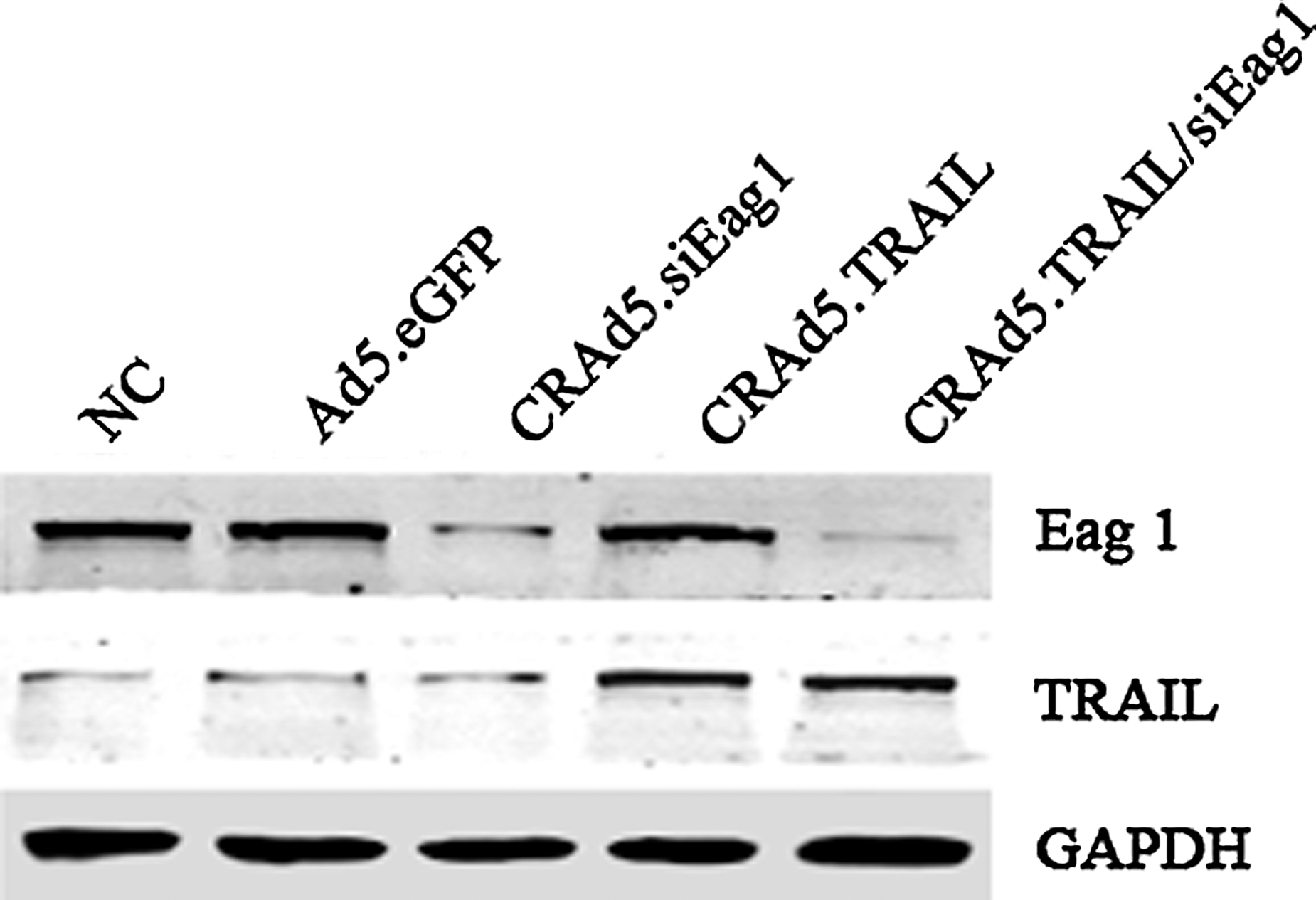
Characterization of CRAd5.TRAIL/siEag1 adenoviral vector. MG-63 cells were infected with different adenoviruses Ad5.eGFP, CRAd5.siEag1, CRAd5.TRAIL, or CRAd5.TRAIL/siEag1 at multiplicity of infection (MOI, calculated as pfu/cell numbers) of 10, respectively. After 48 hours, the relative expression levels of Ether à go-go 1 (Eag1) and tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) were examined by Western blot analysis. NC, negative control (cells without infection).
CRAd5.TRAIL/siEag1 induces the growth arrest and apoptosis of osteosarcoma cells in vitro
To investigate whether silencing of Eag1 and overexpression of TRAIL have synergistic antitumor effects on osteosarcoma cells, we performed cell proliferation assay for MG-63 cells infected with different adenoviral vectors. As shown in Figure 2A, infection with CRAd5.TRAIL/siEag1 resulted in a significantly higher reduction in cell proliferation than infection with Ad5.eGFP, CRAd5.TRAIL or CRAd5.siEag1. These results suggested that Eag1 silencing may render MG-63 cells more susceptible to TRAIL. Meanwhile, neither CRAd5.siEag1, CRAd5.TRAIL nor CRAd5.TRAIL/siEag1 exhibited cytotoxicity in human osteoblastic cells hFOB 1.19 (Fig. 2A). These data indicated that CRAd5.TRAIL/siEag1 had enhanced antitumor activity in human osteosarcoma cells and maintained low toxicity to normal osteoblastic cells.
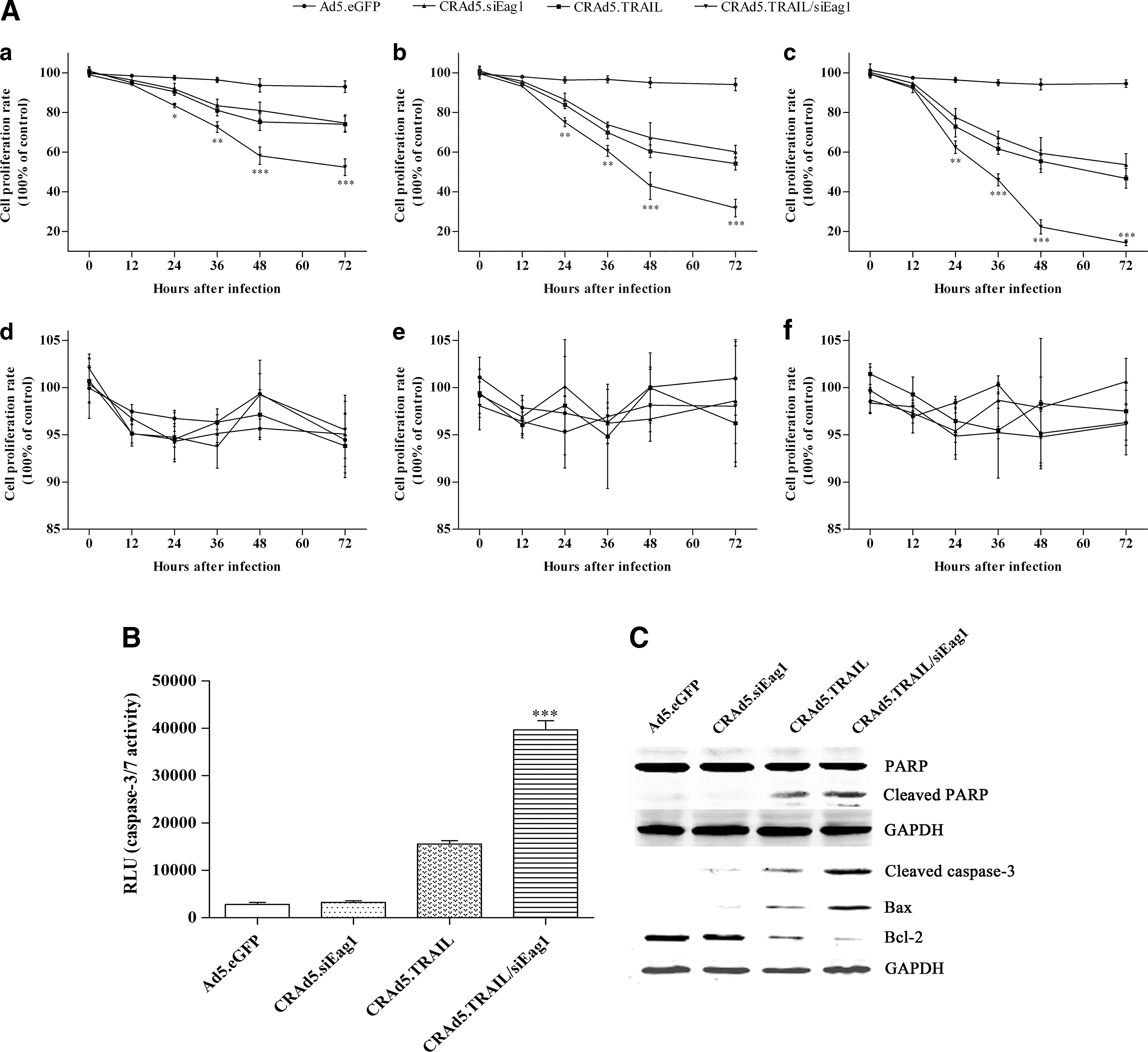
Strong induction of cell growth arrest and apoptosis of MG-63 by CRAd5.TRAIL/siEag1 in vitro.
Next, we performed apoptosis assay. The results showed that the amount of activated caspase-3/7 was significantly higher in CRAd5.TRAIL/siEag1-infected cells than in other adenoviral vector-infected cells (Fig. 2B). It is widely known that PARP is a zinc-finger DNA-binding protein that detects DNA strand breaks and functions in base excision repair. 26 During apoptosis, PARP is cleaved into 89- and 24-kDa fragments by caspase, and PARP cleavage is considered to be one of the classical characteristics of caspase-dependent apoptosis. 27,28 As shown in Figure 2C, the amount of cleaved PARP was significantly higher in CRAd5.TRAIL/siEag1-infected cells than in other adenoviral vector-infected cells. In addition, Western blot analysis showed that the expression of cleaved caspase-3 and Bax was significantly higher, but the level of Bcl-2 was lower in CRAd5.TRAIL/siEag1-infected cells than in other adenoviral vector-infected cells, indicating that CRAd5.TRAIL/siEag1 has stronger effect to induce apoptosis of osteosarcoma cells in vitro. Collectively, these results demonstrated that siEag1 silencing and TRAIL overexpression induced growth arrest and apoptosis of MG-63 cells in a synergistic manner.
CRAd5.TRAIL/siEag1 inhibits tumor growth and promotes apoptosis of osteosarcoma cells in vivo
To characterize the antitumor effects of different adenoviruses in vivo, we made a xenograft model of osteosarcoma using nude mice, and treated the xenografts by intratumor injection of Ad5.eGFP, CRAd5.siEag1, CRAd5.TRAIL, and CRAd5.TRAIL/siEag1, respectively. The results showed that the tumor volume was significantly smaller in CRAd5.TRAIL/siEag1
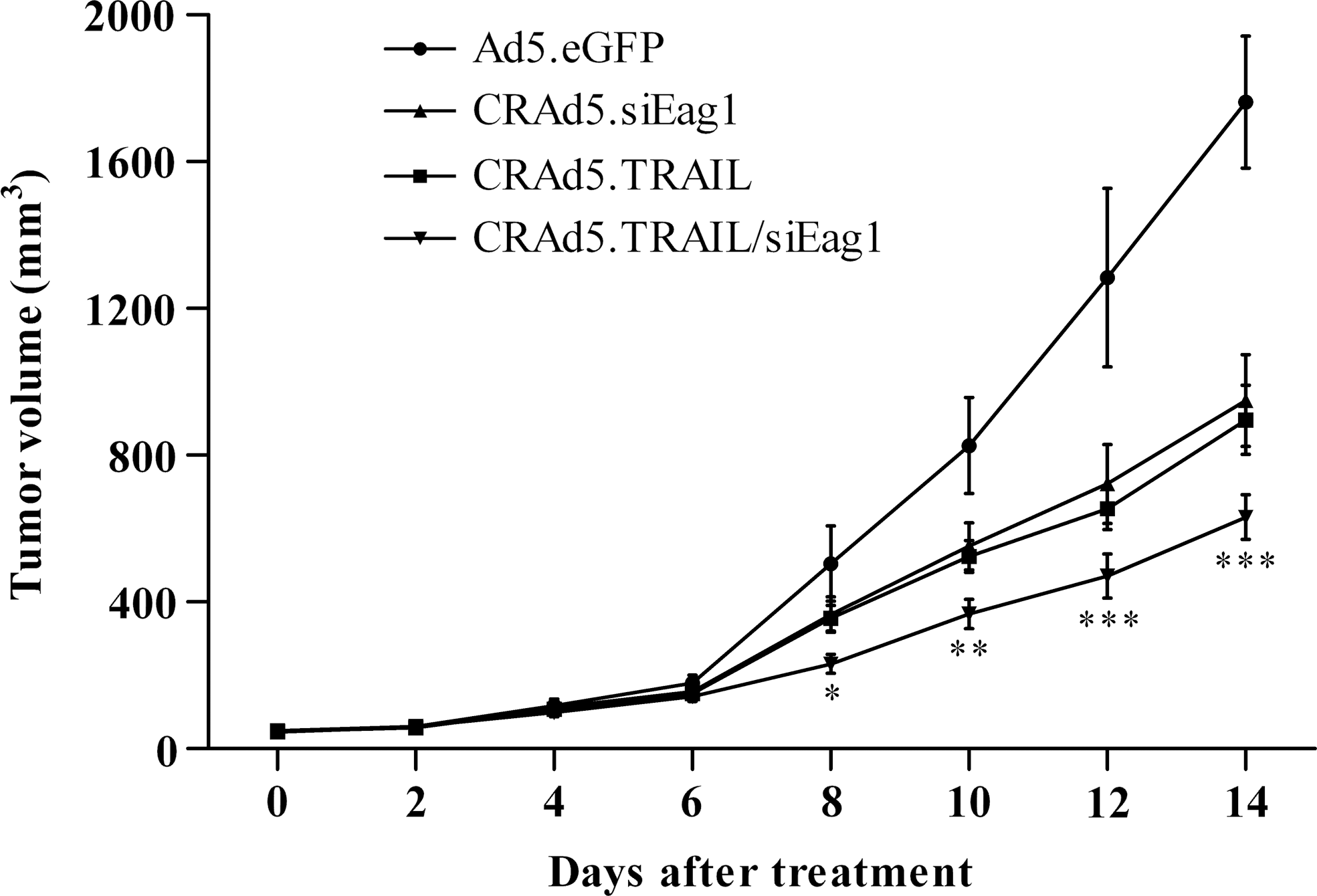
CRAd5.TRAIL/siEag1 significantly inhibits the growth of MG-63 cells in nude mice. Intratumor injection of CRAd5.TRAIL/siEag1 significantly reduced the size of MG-63-derived tumor implanted subcutaneously in nude mice during the 14-day follow-up period. *p<0.05, **p<0.01, ***p<0.001, compared with Ad5.eGFP, CRAd5.siEag1, or CRAd5.TRAIL, respectively (n=6).
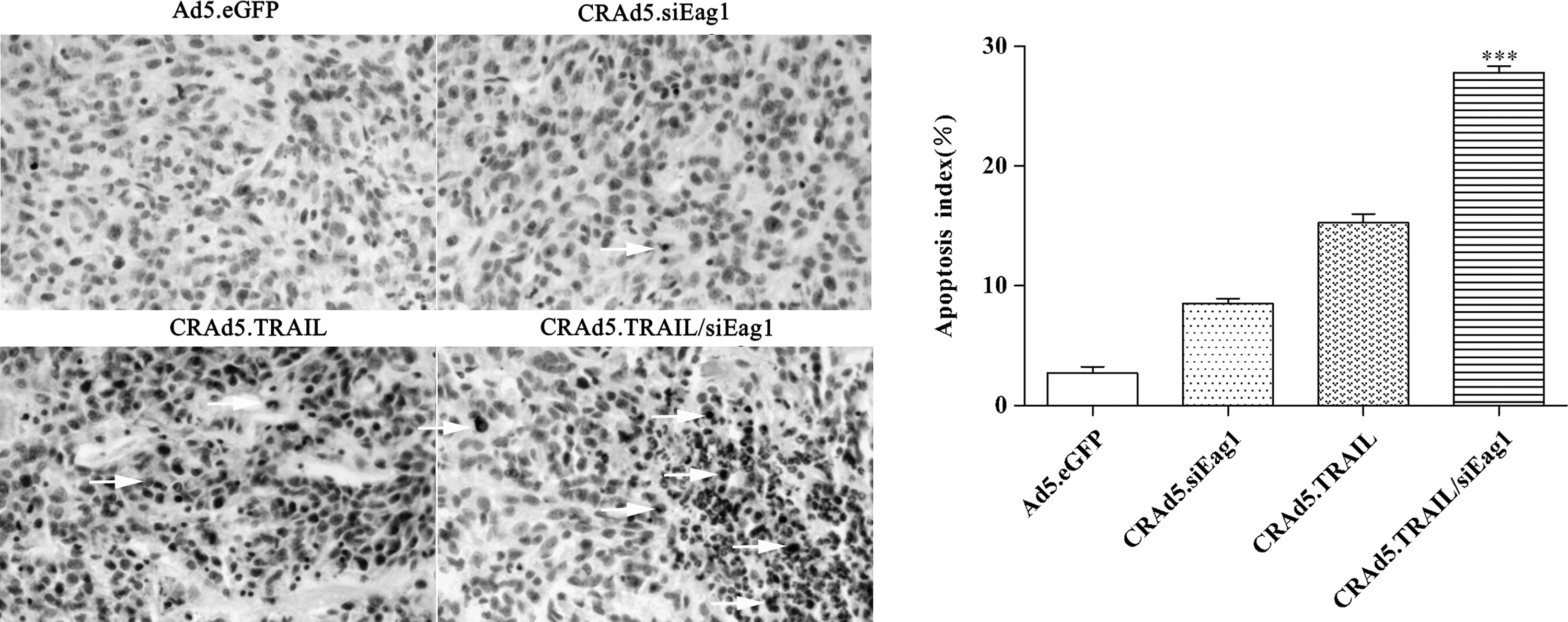
CRAd5.TRAIL/siEag1 significantly induces the apoptosis of MG-63 cells in nude mice. Apoptosis index of MG-63 xenograft tumors in nude mice treated with Ad5.eGFP, CRAd5.siEag1, CRAd5.TRAIL, or CRAd5.TRAIL/siEag1 was calculated based on the TUNEL assay and represented by the average percentage of TUNEL-positive cells from 6 randomly selected areas (mean±SD, n=6). The apoptotic cells were indicated by arrows (
Discussion
The Eag family consists of three closely related subfamilies of genes defined by sequence homology, Eag, Erg (ether-a-go-go related gene), and Elk (ether à go-go like gene). Each of the three subfamilies is defined by the high degree of homology shared among members. 29 The Eag subfamily includes two members Eag1 (KCNH1 and Kv10.1) and Eag2 (KCNH5 and Kv10.2). 30 Normally, Eag1 is exclusively expressed in the brain, slightly in the placenta, and transiently in myoblasts. 31 However, Eag1 is abnormally overexpressed in a variety of cancers, and the use of Eag1-siRNA is a potential new approach to the treatment of cancers, including osteosarcoma.
TRAIL has the unique ability to induce apoptosis in many transformed cell lines, but not in normal tissue. Therefore, TRAIL is one of the most promising candidates for cancer therapeutics. However, how TRAIL induces tumor cell apoptosis is still largely unknown. TRAIL can bind to two death receptors: death receptor 4 (DR4 and TRAIL-R1) and death receptor 5 (DR5 and TRAIL-R2). 32,33 Upon binding to TRAIL, DR4 and DR5 receptors recruit Fas-associated death domain, leading to apoptosis due to caspase-8 activation and caspase-3 cleavage. 34 However, the resistance of some cancer cells to TRAIL is a major obstacle to clinical application of TRAIL. Recently, the combined treatment with TRAIL and other agents has been suggested as new modalities for cancer therapy. A single-chain antibody against Eag1 was fused to TRAIL. Subsequently, selective apoptosis in Eag1-positive cancer cells and the bystander effect in Eag1-negative cancer cells were observed, indicating the synergistic antitumor effect of targeting Eag1 and TRAIL. 35
In the present study, we analyzed the antitumor effects of the silencing of Eag1 expression in combination with adenovirus-mediated TRAIL overexpression. In vitro studies showed that adenovirus-mediated siRNA could efficiently silence Eag1 expression and increase the sensitivity of MG-63 cells to CRAd5.TRAIL-mediated apoptosis, without apparent impacts on hFOB 1.19 cells. To further prove the synergistic efficacy, we performed in vivo experiments, and the data showed that CRAd5.TRAIL/siEag1 could inhibit tumor growth and induce apoptosis in osteosarcoma xenografts in a more efficient manner than CRAd5.siEag1 or CRAd5.TRAIL alone. Taken together, these data demonstrate the synergistic antitumor effect of Eag1 knockdown and TRAIL overexpression for osteosarcoma.
Several mechanisms have been proposed to explain the development of TRAIL resistance in tumor, including the overexpression of genes such as cFLIP. 36 Interestingly, overexpression of Eag1 has been documented in osteosarcoma by our group, 37 and Eag1 overexpression may provide protection against CRAd.TRAIL-mediated apoptosis in MG-63 cells, indicating that the caspase machinery activated by TRAIL in MG-63 cells might be preferentially inhibited by Eag1. Therefore, siRNA-mediated silencing of Eag1 could sensitize osteosarcoma to TRAIL treatment, leading to synergistic antitumor effects.
In conclusion, we developed an adenovirus vector that could simultaneously achieve knockdown of Eag1 and overexpression of TRAIL upon infection into osteosarcoma cells. More importantly, this vector exhibited strong antitumor activity for osteosarcoma cells both in vitro and in vivo, due to the synergistic effects of knockdown of Eag1 and overexpression of TRAIL. Our findings have important implication for the development of new strategies for osteosarcoma therapy.
Footnotes
Acknowledgment
This study was supported by the Fujian Natural Science Foundation, China (No. C0710048).
Disclosure Statement
The authors declare that they have no competing interests.