
Abstract
Osteosarcoma (OS) is the most common primary malignant bone tumor. Despite advances in neoadjuvant multi-agent chemotherapy, the outcome of patients has not significantly improved in the last decades, making the search for more effective therapeutic agents imperative. In the present study, we explored the possibility of using activator protein-1 inhibition by 3-[(dodecylthiocarbonyl)methyl]-glutarimide (DTCM-g) as a new therapeutic strategy in two OS cell lines, HOS and MG-63. Our results showed that low concentrations (2.5, 5, 10, and 20 μg/mL) of the drug significantly decreased cell proliferation and clonogenic capacity, albeit it did not significantly induce cell death. DTCM-g also decreased cell invasiveness, and inhibited PDPN, MMP-2, TIMP1, and TIMP2 expressions. Moreover, our results showed that DTCM-g synergized with ionizing radiation in both cell lines while chemosensitized MG-63 cells to doxorubicin treatment. Even though additional laboratorial and preclinical tests are still needed to support our data, we demonstrate that DTCM-g inhibits growth in OS cells, increases the cytotoxicity of other commonly used agents, and may possess antimetastatic activity.
Introduction
Osteosarcomas (OSs) are highly aggressive primary bone tumors of osteoblastic origin that mainly affect children and young adults. 1 Current treatments consist of initial neoadjuvant treatment with high doses of doxorubicin (DXR), cisplatin (CDDP), and methotrexate (MTX); surgical removal of the primary tumor in the affected bone; and adjuvant chemotherapy. 2 Radiotherapy is indicated only in patients who suffer from inoperable (advanced extremity, axial or head-and-neck) OS, patients with painful bone metastases, and those who refuse surgery. 3
OS are notoriously chemo- and radioresistant, 4,5 and despite the advances achieved with multidisciplinary management the outcome of patients has changed very little over the past 30 years; patients still succumb to metastatic disease and long-term survival is achieved in only two-thirds of all patients. 6 Thus, new chemotherapeutic agents are urgently required to improve treatment and outcome.
In recent years, much attention has been focused on the activator protein-1 (AP-1) complex, a pleiotropic transcription factor composed of heterodimers of proteins belonging to the Fos and Jun families, that plays crucial roles in a variety of cellular events involved in normal development or neoplastic transformation. 7 Papachristou et al., 8 have demonstrated that the AP-1 pathway is directly involved in osteoblast transformation and OS development and progression, showing that the study of the expression of AP-1 members could predict clinical outcome in this neoplasia. Moreover, increased AP-1 activity has been associated with the more metastatic OS phenotype in murine models 9 reinforcing the possibility of using AP-1 inhibition as a therapeutic tool in the management of OS.
Recently, 3-[(dodecylthiocarbonyl)methyl]-glutarimide (DTCM-g), a piperidine derived of 9-methylstreptimidone 10 has shown to specifically block the phosphorylation and nuclear translocation of c-Jun and c-Fos and the consequent transcriptional activation of the AP-1 complex. 11 In the present study, we show evidence of antiproliferative effects of this novel inhibitor of AP-1, on HOS and MG-63 OS cell lines offering a rationale for a therapeutic targeting in this highly aggressive and eventually lethal tumor.
Methods
Cell culture
The human OS cell lines HOS and MG-63 were kindly provided by Dr. Jeremy A. Squire (Department of Pathology and Molecular Medicine, Kingston General Hospital, Queen's University). Cells were cultured in HAM F10 (GibcoBRL, Life Technologies) supplemented with 10% fetal bovine serum, penicillin (100 U/mL), and streptomycin (100 μg/mL) at 37°C in a humidified 5% CO2 incubator.
Drug and treatments
DTCM-g was synthesized as previously described. 12 For all experiments, cells were treated with 2.5, 5, 10, and 20 μg/mL. Corresponding control cultures received an equal volume of solvent (not exceeding 0.1%).
For combinatorial treatments CDDP, MTX, and DXR were purchased from Sigma-Aldrich® and diluted in dimethyl sulfoxide (DMSO) or in 0.9% NaCl accordingly. Dilutions were prepared with final test concentrations of (16 and 32 ng/mL) CDDP, (16 and 32 μM) MTX, and (20 and 40 μM) DXR.
Measurement of cell growth
Cell survival was assessed using the XTT assay (XTT II; Roche Molecular Biochemicals). Briefly, 2500 cells were seeded in 96-well flat-bottom plates and allowed to attach for 24 hours. Subsequently, cells were treated with different concentrations of DTCM-g (2.5, 5, 10, and 20 μg/mL), and incubated for 24, 48, and 72 hours. For combinatorial treatments, cells were simultaneously treated with combinations of DTCM-g 10 μg/mL with CDDP (16 or 32 ng/mL); MTX (16 or 32 μM); or DXR (20 or 40 μM) and incubated for 48 hours. After treatment, the culture medium was removed and replaced with medium containing 10 μL of XTT dye (3 mg/mL) in each well. The plates were incubated for 2 hours at 37°C and the formazan product was measured at 455 and 650 nm by using an iMarkmicroplate reader (Bio-Rad Laboratories®). For growth analyses, trypan blue exclusion and cell counting were also implemented. Each experiment was performed in triplicate wells and repeated in three sets of tests.
Colony formation assay
Clonogenic assays were performed according to Franken et al. 13 Single cell suspensions of 300 cells were seeded in six-well plates and treated with DTCM-g 2.5, 5, 10, and 20 μg/mL concentrations for 24 hours. Following that period, the culture medium was removed and replaced with drug-free medium. The cell cultures were incubated for 10 days and then the colonies were rinsed with phosphate-buffered saline, fixed with methanol and stained with Giemsa. Only colonies with >50 cells were counted. Assays were performed in triplicate.
Analysis of caspase activation
For apoptosis, 3×104 cells were seeded on six-well plates containing 3 mL of culture medium. After 24 hours, the medium was replaced and cells treated with the different concentrations of DTCM-g or vehicle only and cultured for additional 48 hours. Caspase activation was determined using the NucView™ 488 Caspase-3 Detection in Living Cells kit (Biotium, Inc.) according to the manufacturer's instructions. Then, samples were mounted and analyzed under an epifluorescence microscope. Five hundred nuclei were analyzed per treatment and cells were scored and categorized according to differential staining.
Detection of necrotic cells by differential staining
Necrotic cells were recognized by differential staining according to Lee and Shacter. 14 Treated cells were trypsinized, centrifuged and incubated for 5 minutes at 37°C with bisbenzimide (Hoechst 33342), propidium iodide and fluorescein diacetate (Sigma Chemical Co.). Then, samples were mounted, coverslipped and analyzed by fluorescence microscopy with a triple filter. Cells were scored and categorized as follows: (1) normal: blue nucleus and green cytoplasm, (2) apoptotic: fragmented blue nucleus and green cytoplasm and (3) necrotic: red nucleus. Five hundred nuclei were analyzed per treatment.
In vitro scratch assay for analysis of cell migration
In vitro scratch assays to quantify tumor migration rates were performed according to Liang et al. 15 with minor modifications. Briefly cells were grown to confluence on 12 well plates and scratch wounds were then created using a pipet tip (200 μL) and the wound site photographed digitally at time zero. Cells were then treated with different concentrations of DTCM-g and subcultured for 24 hours in medium supplemented with only 1% fetal bovine serum. After that period, cells were photographed and the Motic Images Plus v2.0 software (Motic China Group Co., Ltd.) was then used to calculate the cell-free area. Cell migration rate was calculated as the distance (nanometers) travelled by the cells in this area over time.
Invasion assay
5×105 cells were treated with different concentrations of DTCM-g and transferred to the top of Matrigel-coated invasion chambers (24-well insert, 8-μm pore size; Becton Dickinson & Co.) according to the manufacturer's protocol. After 22 hours incubation, noninvading cells were removed from the upper surface of the membrane by scrubbing with moistened swabs. The invasive cells attached to the lower surface of the membrane insert were fixed in 100% methanol for 10 minutes and stained with Giemsa (Sigma-Aldrich). Membranes were then removed from the insert housing with scalpel blade, placed on a microscope slide, mounted with Entellan®, and coverslipped. Invading cells were photographed under the microscope at 100×magnification and counted with the CytolabView software (Applied Spectral Imaging).
Quantitative real-time polymerase chain reaction
To study the effects of DTCM-g on the expression of genes regulated by AP-1, cells were treated with 2.5 μg/mL concentration and subcultured for 24 hours in medium supplemented with only 1% fetal bovine serum. RNA was extracted with the TRIzol® reagent (GibcoBRL, Life Technologies). The total RNA was converted to single-stranded cDNA using a High Capacity kit (Applied Biosystems) according to the manufacturer's instructions. Measurement of mRNA levels of PDPN, MMP2, TIMP1, and TIMP2 were based on SYBR® Green PCR Master Mix (Applied Biosystems) using the 7500 Real Time PCR System (Applied Biosystems). The primers for genes MMP2, TIMP1, TIMP2, and PDPN were designed using the Oligo 3.0 software (Whitehead Institute/MIT Center for Genome Research). 16,17 The mRNA levels of internal control genes, human GUSB and HPTR, were also measured and geometric average used to normalize the mRNA levels of the studied genes. The real-time amplification reaction was performed in the presence of 200 nM of each forward and reverse primer, and 5 μL SYBR Green PCR Master Mix (Applied Biosystems) in a total volume of 10 μL. In the reaction, 5 μL of diluted (1:10) cDNA was used as a template in duplicate for all samples. To compare the expression levels among different patient samples, the relative expression of the studied genes and GUSB levels was calculated using the 2−ΔΔCT method. 18 Each run was completed with a melting curve analysis to confirm the specificity of amplification and the lack of primer dimmers. Serial dilutions of reference sample cDNA were used to generate standard curves. At least three points were used for the construction of the standard curves, which reached 90%–110% efficiency.
Cell irradiation
To test the effect of DTCM-g on radioresistance clonogenic assays were performed. After tripsinization, single cell suspensions of 500 cells were seeded in six-well plates and treated with DTCM-g (20 μg/mL) for 24 hours. Then, the culture medium was replaced with drug-free medium and cell cultures were irradiated using γ-rays with energy of 1.25 MV, dose rate of 0.48 Gy/min, and final doses with 2, 4, and 6 Gy. The irradiation equipment used is dedicated only for experimental research (Gammatron S-80; Siemens Medical Systems, Inc.) at the University Clinical Hospital (FMRP-USP). After irradiation cell cultures were incubated for 10 days and then the colonies analyzed as described above. The radiation dose enhancement ratio (DER) by DTCM-g was calculated using the following formula: DER=(surviving fraction at an indicated dose of radiation alone)/(surviving fraction at an indicated dose of radiation+DTCM-g). DER=1 suggests an additive radiation effect and DER >1, a supra-additive effect as against a sub-additive effect in the case of DER <1. 19
Statistical analysis
All experiments were performed in triplicate and repeated in three sets of tests. Statistical analyses were performed by using the SigmaStat software (Jandel Scientific Company) and confirmed by using the SPSS software, version 15.0 for Windows. One way repeated measures analysis of variance followed by the Holm–Sidak Pairwise Multiple Comparison was used to establish whether significant differences existed between groups. All tests were carried out for α=0.05. Effective concentrations (IC50) were analyzed using the CalcuSyn software v2.0 (Biosoft). This program provides a measure of the combined drug interaction by the generation of a combination index (CI) value. The CI value is based on the multiple drug-effect equation of Chou and Talalay 20 and defines the drug interactions as synergistic CI value <1, additive CI value=1 or antagonistic CI value >1. The Calcusyn® software was also used to calculate the dose reduction index of drug combinations, which estimates the extent to which the dose of one or more agents in the combination can be reduced to achieve effect levels that are comparable with those achieved with single agents.
Results
DTCM-g inhibits cell proliferation in vitro
DTCM-g effectively reduced the growth of HOS cells after 24 and 48 hours at 20 μg/mL (p<0.05) when compared with control (DMSO 0.1%). Similarly, MG-63 cells showed significant growth reduction after 48 hours for DTCM-g 5, 10, and 20 μg/mL. After 72 hours both cell lines presented reduced growth at all concentrations tested (p<0.05) (Fig. 1A). The maximum effect was observed at the highest concentration after 72 hours reducing the amount of living cells in the cultures by 89% and 77% for HOS and MG-63, respectively (Fig. 1A). IC50 values were determined as 4.20 and 3.76 μg/mL for HOS and MG-63, respectively, after 72 hours of treatment. In addition, cell growth monitored at selected intervals by trypan blue exclusion showed a marked reduction in the number of cells along with increasing DTCM-g concentrations (Fig. 1B).
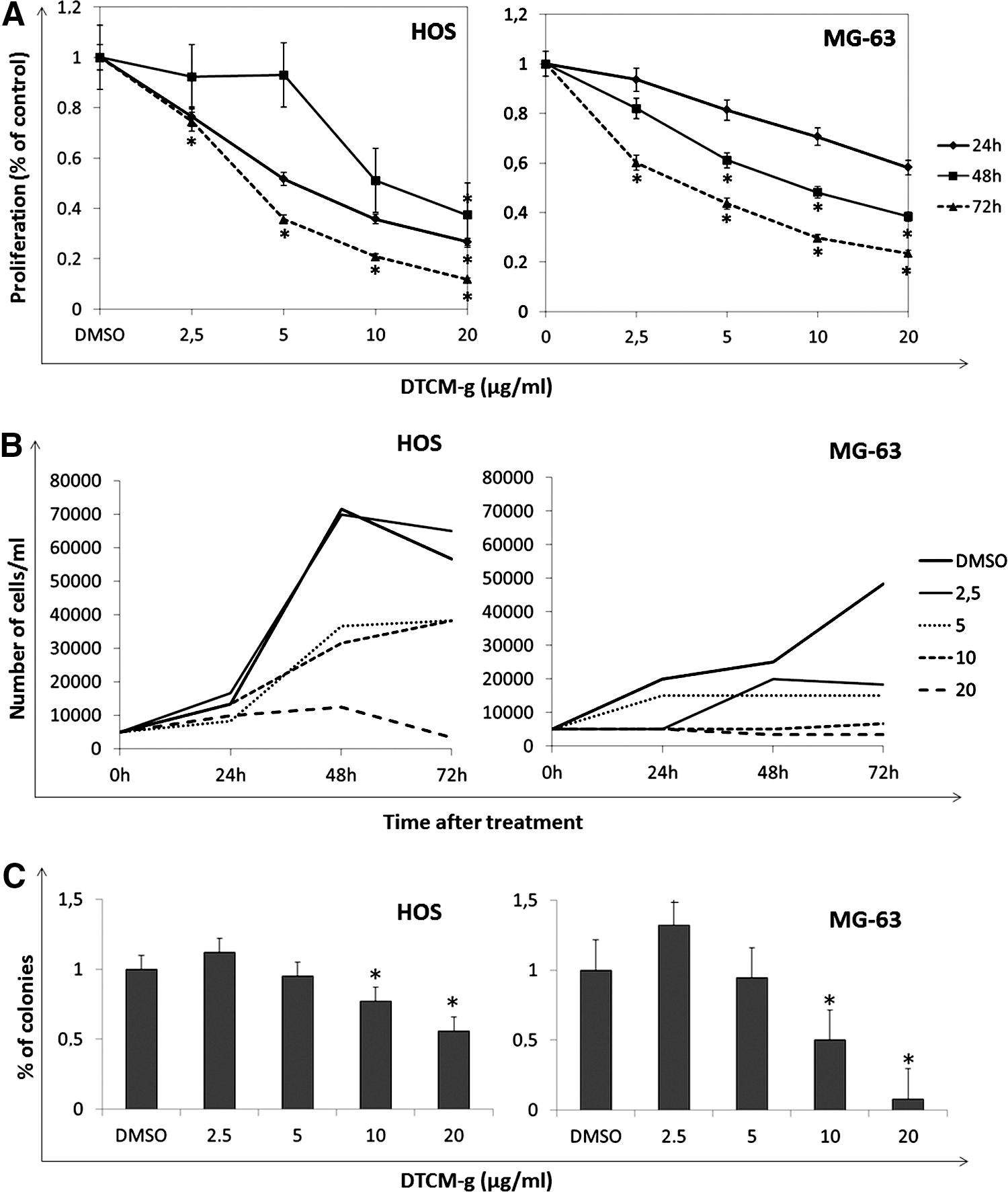
Characterization of the effects of AP-1 inhibition on cell growth and clonogenic survival in HOS and MG-63 cells treated with DTCM-g.
DTCM-g reduces the clonogenic capacity OS cell lines
DTCM-g treatment for 24 hours significantly reduced the colony formation capacity for both cell lines when compared with control (p<0.05) (Fig. 1B). At 10 and 20 μg/mL treatments the clonogenic capacity of HOS cell line was reduced in 23% and 44%, respectively. MG-63 cells resulted more sensitive with reductions of 50% and 91% for the same concentrations.
DTCM-g does not induce cell death in HOS and MG-63 cells
The analysis of cells treated with DTCM-g through caspase-3 activation showed no effect on apoptosis rates. Comparatively, the analysis of treated cells by differential fluorescent staining with propidium iodide was also unable to demonstrate higher frequency of necrotic-like cells for both cell lines, suggesting that reduced cell numbers in vitro result from impaired proliferation rather than cell death (data not shown).
DTCM-g inhibits cell migration and invasion
DTCM-g significantly reduced both OS cell lines migration as measured by in vitro wound healing assays (p<0.05), however dose-dependency was only observed for MG-63 cells (Fig. 2A). Invasion assay using transwell chambers coated with Matrigel® showed significant reductions of invasion by 13% and 47% compared with controls in HOS cells after treatment with 10 and 20 μg/mL, respectively. In the case of MG-63 cells, the inhibition was more effective with a reduction of almost 80% for both treatments (Fig. 2B). The effects of DTCM-g on invasiveness were also evinced by the decreased expression of AP-1 regulated genes, including PDPN, MMP2, TIMP1, and TIMP2 (Fig. 2C).
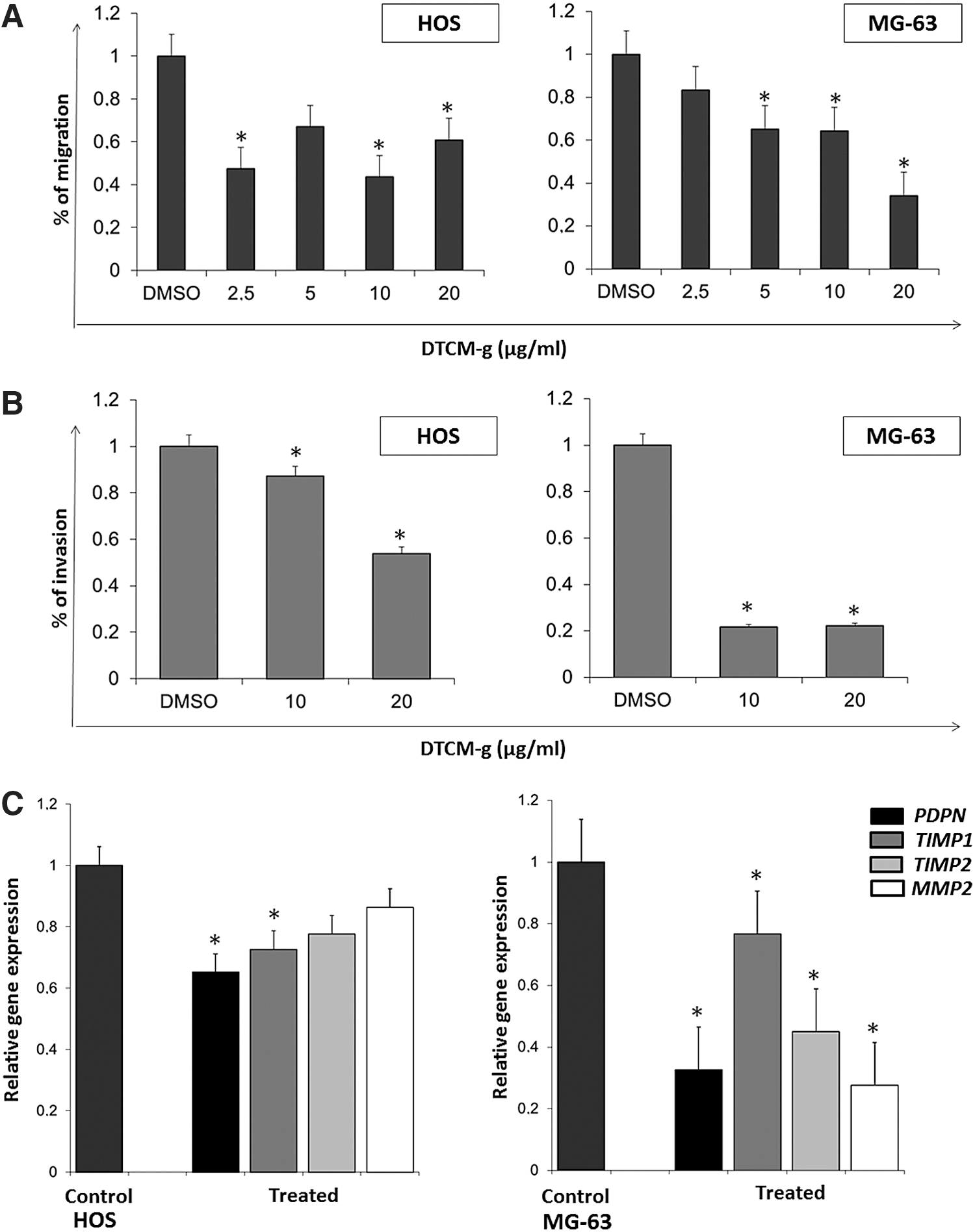
Pretreatment with DTCM-g sensitizes cells to ionizing radiation
To study the cytotoxic effects of DTCM-g in association with γ-radiation, HOS and MG-63 cells were incubated with 20 μg/mL concentration of the drug for 24 hours. After the treatment, the cell culture medium was replaced and cells irradiated with a final doses of 2, 4, and 6 Gy. The results showed that DTCM-g pretreatment led to radiosensitization for both cell lines showing supra-additive effects at all doses tested (Table 1).
DER=1 suggests an additive radiation effect and DER >1, a supra-additive effect as against a sub-additive effect in the case of DER <1.
DER, dose enhancement rates; DTCM-g, 3-[(dodecylthiocarbonyl)methyl]-glutarimide.
DTCM-g sensitizes MG-63 cells to DXR
To study the cytotoxic effects of DTCM-g in association with commonly used drugs, HOS and MG-63 cells were concomitantly treated with DTCM-g 20 μg/mL and different concentrations of CDDP, DXR, and MTX. Our results showed that co-treatment with DTCM-g did not appreciably sensitize OS cells to CDDP and MTX induced cytotoxicity (CI>1). Conversely, sensitization of MG-63 cells to DXR was achieved at 40 μM concentration (Table 2).
CI values <1 correspond to a synergistic interaction. DRI reflects the fold reduction in the required concentration of tested agents when used in combination to achieve the comparable AF.
AF, affected fraction; CI, combination index; DRI, dose reduction index; CDDP, cisplatin; DXR, doxorubicin; MTX, methotrexate.
Discussion
Contemporary treatment of OS involves aggressive chemotherapy, extensive surgery, and occasional radiation. However, these tumors frequently exhibit chemo- and radioresistance, contributing to a poor clinical response and patient outcome. As a result, identifying novel strategies to overcome this resistance may aid in the development of improved therapeutics.
Altered expression of AP-1 members has repeatedly been described to be associated with many types of human cancer. 21 –24 Moreover, constitutive activation of AP-1 was significantly associated with poor prognosis in lymphomas, hepatocarcinomas, melanomas, and gastric tumors. 25 –28 Overexpression of AP-1 members has also shown to be directly involved in osteoblast transformation and progression of OS 8,29 –31 , representing a potential target for therapy intervention.
In recent years, several chemical drugs with potential AP-1 inhibiting activities have been synthesized, such as geldanamycin, 32 flavopiridol, 33 or T5224. 34 Natural compounds such as curcumin, 35 Silibinin 36 , and cinnamon extract 37 have also shown to inhibit AP-1, though the exerted inhibition may be indirect or not selective.
In the present study, we explored the possibility to use DTCM-g, a novel inhibitor of AP-1 that exerts its activity by blocking the phosphorylation and nuclear translocation of c-fos and c-jun, as a new therapeutic strategy in OS. Our results showed an impaired survival and clonogenic capacity after exposure to treatment with low concentrations of the drug in both OS cell lines. The ability of cancer cells to form colonies is an indispensable condition for the spread of a malignant tumor to distant organs. 38 OSs frequently metastasize to the lung 39 and our results also evinced a significant inhibition of the migratory potential of both cell lines after exposure to DTCM-g. Extensive data demonstrate that AP-1 plays a major role in the transcriptional regulation of direct effectors of invasion such as metalloproteinases (MMP1, MMP2, MMP3, MMP7, MMP10, MMP11, and MMP13), PDPN, CD44, Ezrin, and Krp1. 40 –42 AP-1 overexpression was previously associated with increased motility and invasion in OS 9 being essential during the early stages of metastasis. In support of this possibility, DTCM-g significantly decreased the expression of AP-1 regulated genes, including PDPN, MMP2, TIMP1, and TIMP2, all of which may contribute to invasion process. Similar results have also been demonstrated after inhibition of AP-1 action by Silibinin 36 and through the catalysis of c-Jun mRNA by the DNAzymeDz13, which also has activity against tumor spreading to the lungs in xenografted models. 43,44
On the other hand, overexpression or constitutive activation of AP-1 has repeatedly shown to contribute to the enhanced chemoresistance in different tumors such as gliomas, 45 –48 breast carcinomas 49,50 , and leukemia. 51 In the present study, inhibition of AP-1 by co-treatment with DTCM-g enhanced the response to DXR of MG-63 cells, though it was not able to sensitize neither of the cell lines studied to CDDP or MTX in our experimental setting. Akin to our findings c-Jun abrogation by Dz13 also showed sensitizing effects to DXR. 43 This drug is a cornerstone chemotherapeutic agent for OS treatment, though the favorable therapeutic response to DXR is often associated with severe toxicity. Therefore, the inadequacy of current chemotherapeutic protocols requires new tactics.
In this respect, radiotherapy presents itself as a valid tool for chemoresistant OS. Gamma-radiation is a locally effective and common treatment strategy used in cancer, 52 though its use in OS treatment was long considered controversial because of primary radioresistance. Nonetheless, current data suggest that the administration of radiotherapy may have a positive influence on prognosis as local treatment of unresectable tumors, following intralesional resection, or as palliation of symptomatic metastases. 53 However, in disseminated tumors, even doses as high as 60 Gy do not guarantee local control and adverse side effects may also limit the dose that can be offered. 54 Thus, finding agents that sensitize malignant cells to radiation would further intensify tumor response and mitigate toxicity to neighboring organs, as effective therapeutic doses could be lowered. In the present study, inhibition of AP-1 by DTCM-g potentiated the cytotoxicity of radiation at very low doses (2, 4, and 6 Gy), depicting a radiosensitizer profile of DTCM-g in OS cell lines. This attribute may be a very important aspect of this new AP-1 inhibitor since the high irradiation doses needed for tumor control may be prohibitive for patients with lesions in the proximity of delicate structures. Although the mechanisms by which AP-1 activity leads to radiation induced cell death are currently undefined, there is evidence suggesting the involvement of Fra-1, Fra-2, and JunD. 55
Taken together, our results provide evidence that strategies focused on inhibiting AP-1 may hamper the metastatic potential and sensitize OS cells to commonly used cytotoxic agents and ionizing radiation. These findings support prospective preclinical and clinical evaluations on the therapeutic intervention with the AP-1 inhibition by DTCM-glutarimide in advanced or disseminated OSs.
Footnotes
Acknowledgments
We are grateful to Leonardo L. Amaral for his cooperation on the irradiation of cell cultures and to Gabriela M. Roberto for her assistance with migration and invasion assays.
Disclosure Statement
No competing financial interests exist.