
Abstract
Cisplatin-based chemotherapy is considered a golden standard for treatment of advanced non-small cell lung cancer (NSCLC). However, drug resistance is one of the major problems in NSCLC chemotherapy. The mechanisms and related biological pathways that contribute to chemoresistance are relatively poorly understood. Here, we demonstrated that the phosphorylation of eukaryotic translation initiation factor 2α (eIF2α) suppresses cisplatin-induced A549 cell apoptosis. Cisplatin induced eIF2α phosphorylation through protein kinase RNA. Importantly, phospho-eIF2α inhibited cisplatin-induced A549 cells apoptosis, at least in part, by suppressing the p38 pathway. Moreover, analysis of tissue microarrays information demonstrated that phospho-eIF2α predicted a poor prognosis in patients with NSCLC. Taken together, these results provide a potential mechanism that is used for explaining how eIF2α promotes cisplatin resistance in A549 cells. Therefore, the regulation of eIF2α may improve treatment outcomes of cisplatin-based chemotherapy for patients with NSCLC.
Introduction
Lung cancer is one of the most prevalent cancers in the world and is the leading cause of cancer morbidity worldwide. 1,2 Lung cancers are classified into two major cell types by histology: small-cell lung cancer (SCLC) and non-small-cell lung cancer (NSCLC). NSCLC represents ∼80% of lung cancer diagnoses. 3,4 More than 75% of patients with NSCLC have already developed locally invasive or distant metastatic disease at initial diagnosis. 5,6 It has been reported that chemotherapy increases the survival time for patients with NSCLC. 7 Chemotherapy has become a recommended treatment for early-stage NSCLC.
Cisplatin (CDDP) is a platinum-based cytotoxic compound that forms intra- and inter-strand adducts with DNA. Cisplatin is widely used for chemotherapy of a broad range of solid tumors, such as head and neck tumors, hepatocellular carcinoma, ovarian cancer, and advanced NSCLC. 8 –11 Cisplatin-based chemotherapy is considered a golden standard for the treatment of advanced NSCLC, as it can significantly improve survival rates and provide symptom relief in patients with NSCLC. 12 However, the clinical application of cisplatin in NSCLC chemotherapy is limited by acquired or intrinsic resistance of cells to this drug. The molecular mechanisms that underlie cisplatin resistance are poorly understood. Therefore, there is an urgent need to unravel the underlying mechanisms of cisplatin resistance in cancer cells.
Cisplatin has been reported to initiate endoplasmic reticulum (ER) stress, which inhibits cisplatin-induced apoptosis. 13 –15 Previous studies demonstrate that ER stress response can induce eukaryotic translation initiation factor 2α (eIF2α) phosphorylation on S51. 16 The phosphorylation on S51 of eIF2α results in the blocking of protein synthesis initiation through an inhibition of the guanine nucleotide exchange factor. 16 Phosphorylation of eIF2α is mediated by four kinases: protein kinase RNA (PKR)-like ER kinase (PERK), RNA-dependent protein kinase (PKR), general control non-derepressible-2 (GCN2), and heme-regulated kinase (HRI). 16 –20 Among them, PKR is the most widely studied member. The phosphorylation of eIF2α plays an important role in inducing apoptosis and cell cycle arrest in cancer cells. 20,21 Whether eIF2α is involved in cisplatin resistance in cancer cells is unknown.
Here, we report that PKR/eIF2α activation inhibits cisplatin-induced apoptosis in NSCLC cell line A549 via p38 inhibition.
Materials and Methods
Materials
Cisplatin and PKR inhibitor C16 were purchased from Sigma Chemical Company; eIF2α dephosphorylation inhibitor Salubrinal, p38 inhibitor SB203580, and an antibody against β-actin were purchased from Santa Cruz Biotechnology. Antibodies against PARP, phospho-p38, p38, phpspho-eIF2α (Ser51), and eIF2α were purchased from Cell Signaling Technology.
Cell culture and treatment
Human NSCLC cell line A549 was maintained at 37°C in a humidified incubator containing 5% CO2, in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum and 1% penicillin/streptomycin/gentamicin.
Western blot assay
A549 cells were harvested in the appropriate buffer for the purpose of each experiment. Protein concentrations were measured using the BCA assay (Santa Cruz Biotech). Equal proteins were applied to SDS-PAGE. After electrophoresis, proteins were blotted to PVDF membranes. Then, the blots were blocked with 5% skim milk powder supplemented with 0.1% Tween-20, probed with the relevant primary antibodies for 2 hours, and, subsequently, incubated with the appropriate horseradish-peroxidase-conjugated secondary antibodies for 1.5 hours at room temperature. Immunoreactive material was detected using the ECL kit (Santa Cruz Biotech) according to the manufacturer's instructions.
Immunohistochemistry
Immunohistochemistry on tumor arrays was performed as follows. The slides were incubated with the following primary antibodies: anti-phpspho-eIF2α (1:50) rabbit monoclonal. Anti-rabbit peroxidase-conjugated secondary antibodies (Santa Cruz Biotech) were applied. Diaminobenzidine colorimetric reagent solution from Dako was used. Slides were counterstained with hematoxylin (Sigma). The density of positive staining was measured with the use of a computerized image system (Leica Microsystems Imaging Solutions Ltd.). Under high-power magnification (×200 and×400), photographs of three representative fields were captured by the Leica QWin Plus v3 software; identical settings were used for each photograph. The density was counted by Image-Pro Plus v6.2 software (Media Cybernetics, Inc.). For the reading of each antibody staining, a uniform setting for all the slides was applied. Integrated optical density of all the positive staining in each photograph was measured, and its ratio to the total area of each photograph was calculated as density.
Statistical analysis
An analysis was performed with SAS 9.1.3 software (SAS Institute, Inc.); the t test, paired Wilcoxon signed-rank test, or Spearman rank correlation test was used to compare quantitative variables; and qualitative variables were analyzed by the χ 2 test or Fisher's exact test. The statistical differences between these groups were analyzed by one-way analysis of variance. The survival was determined by Kaplan–Meier analysis. Log-rank test was used to compare patients' survival between subgroups. Data were presented as the mean±SEM. All statistical tests were two sided, and P<0.05 was considered statistically significant.
Results
Effect of cisplatin on A549 cells apoptosis
To evaluate the sensitivity of A549 cells to cisplatin-induced apoptosis, we treated A549 cells with various doses of cisplatin for 24 hours. As shown in Figure 1A, cisplatin induced apoptosis of A549 cells in a concentration-dependent manner. Furthermore, time-course experiments revealed that cisplatin induced A549 cells apoptosis in a time-dependent manner (Fig. 1B). These data suggest that cisplatin can initiate A549 cells apoptosis.

Effect of cisplatin on A549 cells apoptosis.
Cisplatin initiates eIF2α and p38 phosphorylation in A549 cells
To examine the effect of cisplatin on eIF2α and p38 phosphorylation, the protein levels of phospho-eIF2α and phospho-p38 were determined in cisplatin-treated A549 cells. We found that cisplatin elevated the protein levels of phospho-eIF2α in a concentration- and time-dependent manner in A549 cells (Fig. 2A, B). Similarly, the phosphorylation level of p38 was also increased in a time-dependent manner in cisplatin-treated A549 cells (Fig. 2C). These results suggest that cisplatin imitates the phosphorylation of eIF2α and p38 in A549 cells.
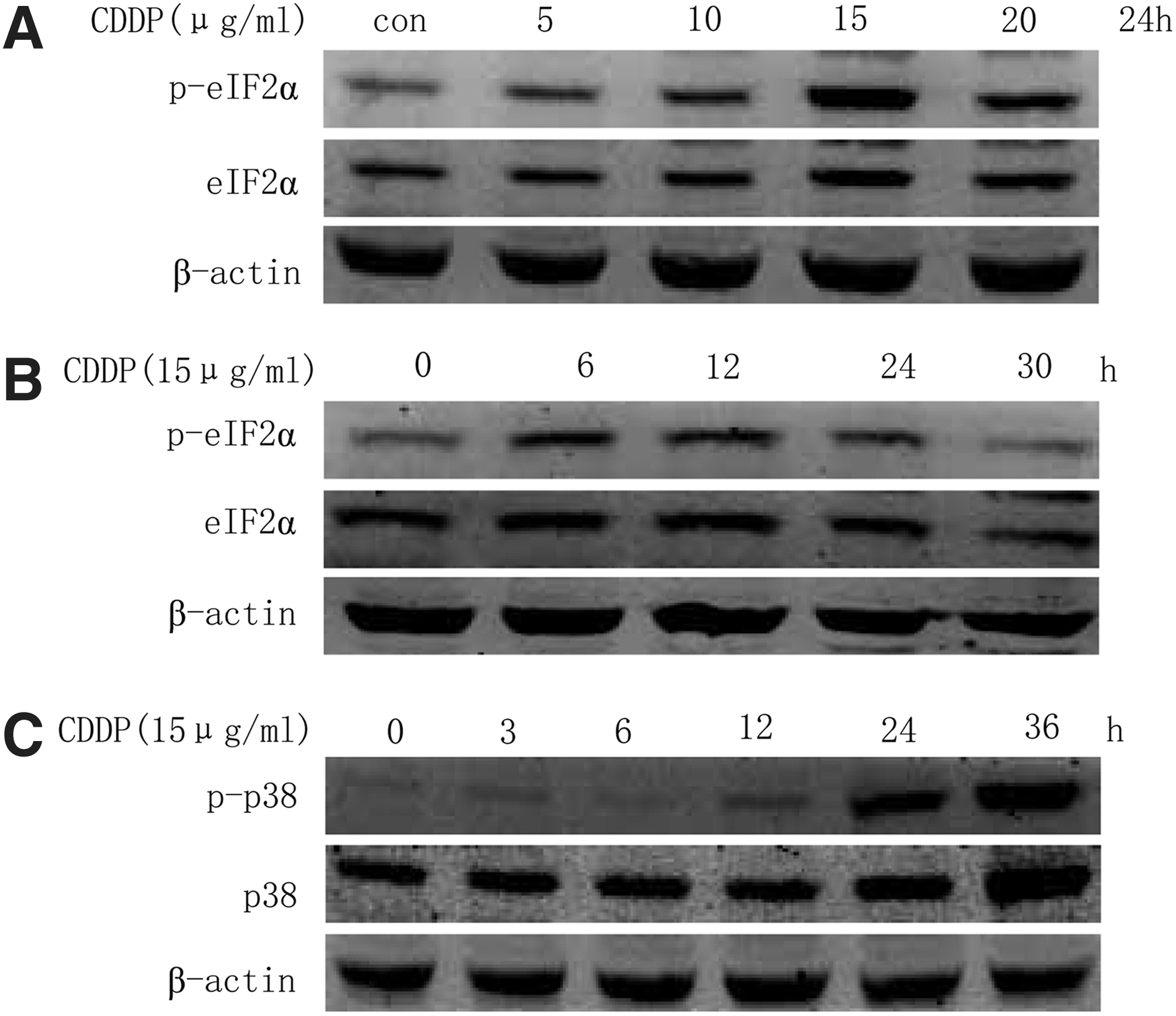
Cisplatin initiates eukaryotic translation initiation factor 2α (eIF2α) and p38 phosphorylation in A549 cells.
PKR/eIF2α signaling cascade is associated with cisplatin resistance
To determine the role of eIF2α in cisplatin-induced apoptosis, we examined the effect of phospho-eIF2α on cisplatin-induced A549 cells apoptosis. We found that salubrinal (which selectively blocks the dephosphorylation of eIF2α) pretreatment inhibited cisplatin-induced A549 cells apoptosis (Fig. 3A, B). To investigate whether PKR was involved in cisplatin-induced eIF2α phosphorylation, we treated A549 cells with PKR inhibitor C16 before cisplatin incubation. The result showed that C16 inhibited cisplatin-mediated eIF2α phosphorylation. Furthermore, C16 preincubation promoted A549 cells apoptosis on cisplatin treatment (Fig. 3C, D). Taken together, these data suggest that cisplatin initiates the phosphorylation of eIF2α through PKR, and the activation of PKR/eIF2α signaling is involved in cisplatin resistance in A549 cells.
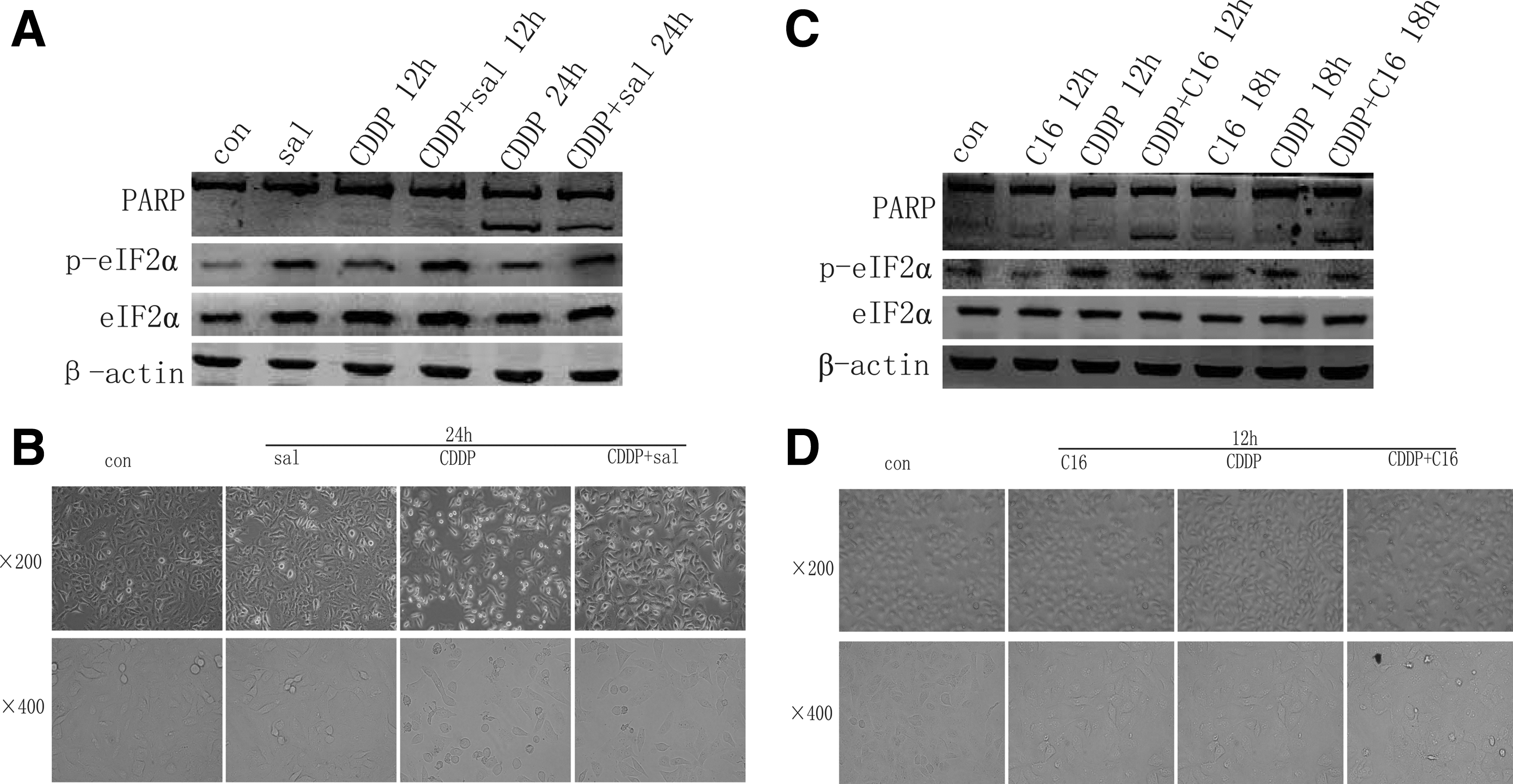
Protein kinase RNA (PKR)/eIF2α signaling cascade is associated with cisplatin resistance.
eIF2α phosphorylation inhibits p38 activation in A549 cells
Since p38 plays an important role in apoptosis regulation, it is interesting to investigate the role of p38 activation in cisplatin-induced A549 cells apoptosis. SB203580, a selective inhibitor of p38, was used to block cisplatin-induced p38 activation in A549 cells. We found that SB203580 apparently inhibited cisplatin-induced A549 apoptosis (Fig. 4A). More importantly, salubrinal preincubation decreased the phosphorylation levels of p38 in cisplatin-treated A549 cells (Fig. 4B). These results, when taken together, indicate that the activation of PKR/eIF2α signaling promotes cisplatin resistance, at least in part, through the inhibition of p38 in A549 cells.
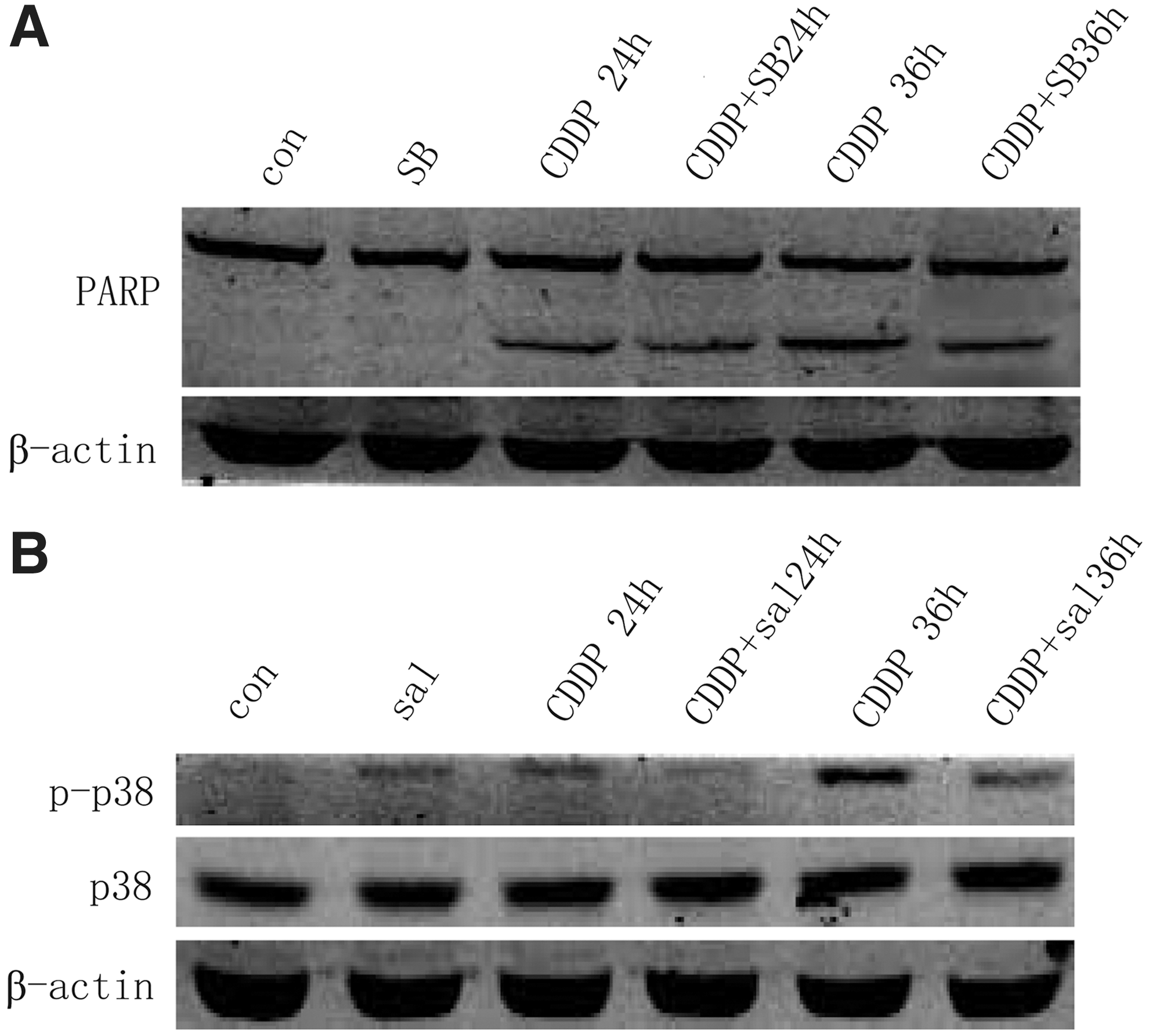
eIF2α phosphorylation inhibits p38 activation in A549 cells.
eIF2α phosphorylation predicts a poor prognosis in patients with NSCLC
Since eIF2α plays an important role in cisplatin resistance in A549 cells, we investigated whether phospho-eIF2α expression in NSCLC is associated with malignant degree and poor survival. Tissue microarrays from 61 patients with NSCLC who had undergone lung resection were examined by immunostaining with phospho-eIF2α monoclonal antibody. All 61 patients with NSCLC were divided into two groups: the high-expression (33 patients) and low-expression (28 patients) group (Fig. 5A). The phospho-eIF2α protein expression levels were found to be significantly higher in NSCLCs with increased tumor volume (p=0.047), gender (p=0.009), lymph node metastasis rate (p=0.000), and histologic typing (p=0.001) (Table 1). Consistently, the 5-year OS after surgery were much worse for phospho-eIF2α-high than for phospho-eIF2α-low expression group. (5-year OS, median OS time was 47 and 66 months, respectively, difference=19 months, p=0.046) (Fig. 5B and Table 2). Thus, phospho-eIF2α expression is a valuable predicting factor for malignant degree and survival in patients with NSCLC.
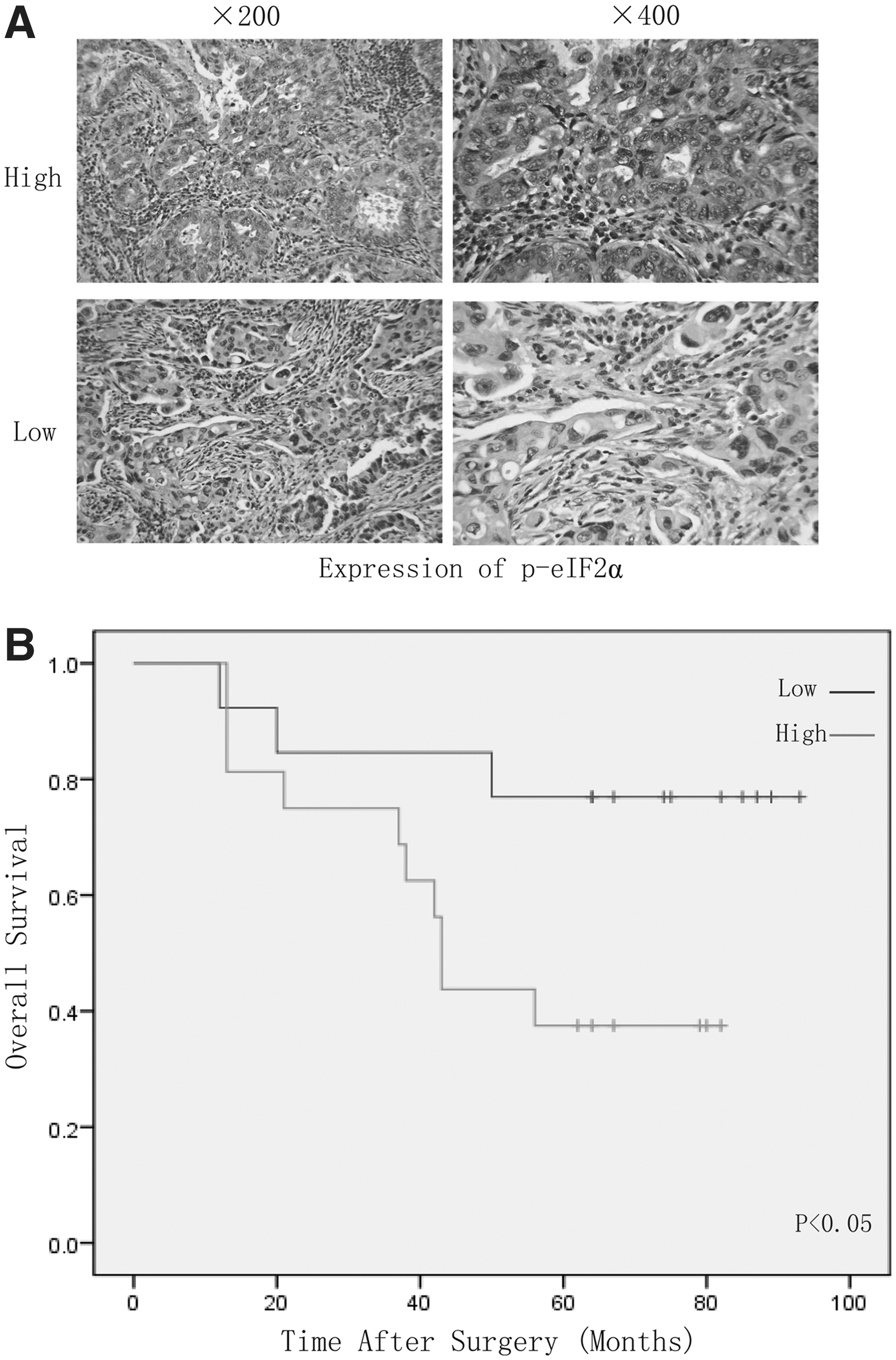
eIF2α phosphorylation predicts a poor prognosis in patients with non-small-cell lung cancer (NSCLC).
Patients with NSCLC were divided into p-eIF2α “high” group (whose final density was higher than the median) and “low” group (whose final density was lower than the median). The patient and disease profiles in each group were compared. Lymph node metastasis rate=positive lymph node number/total biopsied lymph node number.
NS, not significant between any groups; NSCLC, non-small-cell lung cancer; eIF2α, eukaryotic translation initiation factor 2α.
OS, overall survival.
Discussion
Cisplatin is the important established chemotherapeutic agent that is currently available for antitumor treatment of advanced NSCLC. It can significantly improve survival rates and provide symptom relief in patients with NSCLC. However, resistance to cisplatin is a significant impediment to successful chemotherapy. Current research demonstrates that chemoresistance has been associated with decreased susceptibility of cancer cells to apoptosis, thus raising the possibility that increases in cancer cell death determinants may influence treatment outcomes. Here, we report that eIF2α phosphorylation is involved in cisplatin resistance in NSCLC cell line A549.
Cisplatin initiated the phosphorylation of eIF2α in A549 cells. The current studies demonstrate that phosphorylation of eIF2α is mediated by four kinases, PKR, PERK, GCN2, and HRI, under distinct forms of stress condition. Among them, PKR is the most widely studied member. 22 In this study, we found that cisplatin-induced eIF2α phosphorylation was mediated, at least in part, by PKR. Further studies are needed to investigate whether other kinases are involved in cisplatin-induced eIF2α phosphorylation in A549 cells. It has been reported that ER stress plays a pivotal role in protecting cancer cells from cisplatin-induced apoptosis. It should be noted that PKR/eIF2α activation inhibited cisplatin-induced A549 cells apoptosis.
p38 signaling is involved in the regulation of cell death that is induced by many chemotherapeutic agents. Cisplatin clearly induced the activation of the p38 pathway, and SB203580 apparently inhibited cisplatin-induced apoptosis. Based on these data, it is reasonable to suggest that p38 promotes cisplatin-induced A549 cells apoptosis. More importantly, our results demonstrated that eIF2α phosphorylation decreased the activation of p38 on cisplatin treatment, indicating that eIF2α phosphorylation promotes cisplatin resistance through, at least in part, the inhibition of the activation of p38 in A549 cells. Further studies are needed to investigate the detailed link between the PKR/eIF2α and p38 pathways on cisplatin treatment. There is mounting evidence for the involvement of autophagy in mediating resistance of cancer cells to anticancer therapy. 23 Therefore, we investigated the effect of eIF2α on autophagy regulation in cisplatin-treated A549 cells. Our data suggest that autophagy is not involved in eIF2α-mediated cisplatin resistance in A549 cells (data not shown).
An analysis of tissue microarrays information demonstrated that the phospho-eIF2α protein expression levels were found to be significantly higher in male patients, squamous-celled carcinoma. As we all know, smoke is a high risk of squamous-celled carcinoma; so, it is interesting to know whether smoke is relevant to the activation of eIF2α. Several recent studies have begun to amass data showing that the induction of ER stress in combination with chronic activation of multiple effectors of the UPR pathway elements by chronic smoke may play a pivotal role in the etiology and progression of lung cancers and other pulmonary diseases. Perturbations in phospho-eIF2α may have diagnostic and therapeutic potential in the clinical management of patients with various lung malignancies. 24 –26 Consistently, we found that the tumor volume, lymph node metastasis rate, and 5-year OS after surgery were much worse for phospho-eIF2α-high than for phospho-eIF2α-low expression groups.
Conclusion
Our present work identified eIF2α as an important regulator that controls the sensitivity of A549 cells to cisplatin. In particular, phospho-eIF2α inhibits the activation of p38, consequently leading to cisplatin resistance (Fig. 6). Moreover, the data has led us to propose that phospho-eIF2α protein is a novel marker in the prognosis of NSCLC, which predicts malignant degree and poor survival in patients. We believe that there will be more emerging studies about eIF2α protein that contribute to the understanding of molecular mechanisms of cisplatin resistance and the development of new therapeutic strategies against lung cancer.
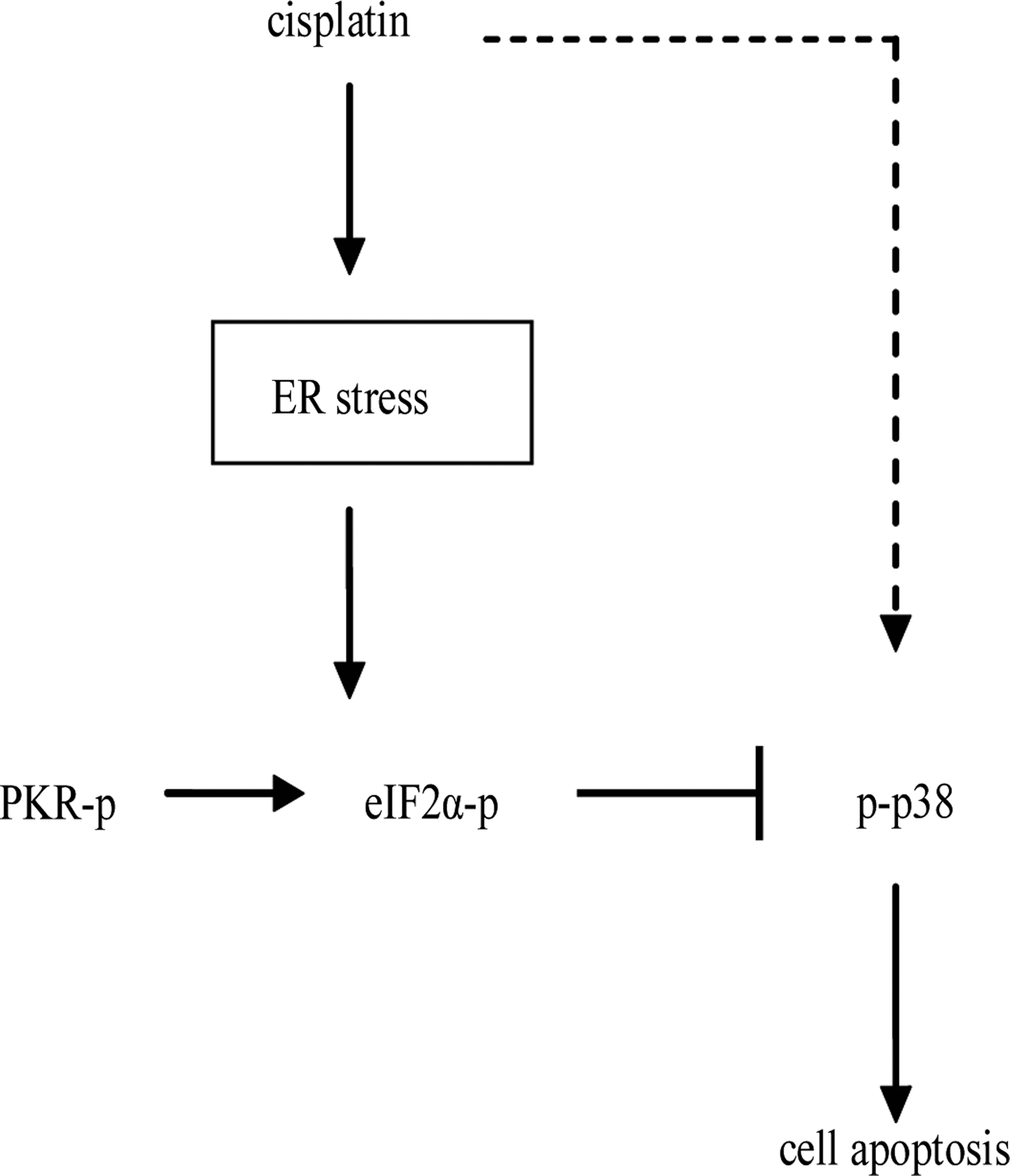
Schematic summarizing the mechanisms by which PKR/eIF2α signaling induces cisplatin resistance in A549 cells.
Footnotes
Acknowledgment
This work was supported by the project from the Health Bureau of Shanghai (2010131).
Disclosure Statement
There were no financial conflicts as required by law.