
Abstract
Background:
Osteosarcoma (OS) is a common malignant bone cancer and usually occurs in adolescents and children. Circular RNAs (circRNAs) play essential roles in tumor development and progression. This study aimed to explore the function and molecular basis of circ_0016347 in OS progression.
Materials and Methods:
The levels of circ_0016347, miR-1225-3p, and ether à go-go 1 (KCNH1) were measured by quantitative real-time polymerase chain reaction or Western blot assay. Cell proliferation was assessed by 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide (MTT) assay and colony formation assay. Cell migration and invasion were evaluated by transwell assay. Glucose consumption and lactate production were detected by glucose detection and lactic acid detection kits. The levels of Ki-67, matrix metalloproteinase-9 (MMP-9), and hexokinase-2 (HK2) were examined by Western blot assay. The interaction among circ_0016347, miR-1225-3p, and KCNH1 was validated by dual-luciferase reporter assay. Xenograft assay was conducted to analyze tumor growth in vivo.
Results:
Circ_0016347 and KCNH1 were upregulated, while miR-1225-3p was downregulated in OS tissues or cells. Circ_0016347 and KCNH1 promoted proliferation, migration, invasion, and glycolysis of OS cells. Circ_0016347 regulated OS progression by modulating KCNH1. Circ_0016347 was a sponge of miR-1225-3p, and miR-1225-3p targeted KCNH1. Circ_0016347 regulated KCNH1 expression via sponging miR-1225-3p. Moreover, silencing of circ_0016347 inhibited tumor growth in vivo.
Conclusion:
Circ_0016347 contributed to OS progression through the miR-1225-3p/KCNH1 axis, which might provide a promising biomarker for OS therapy.
Introduction
Osteosarcoma (OS) is a common malignant bone cancer with high invasiveness and rapid systemic metastasis. 1 OS is usually formed in the bones of children and adolescents. 2 With the development of surgery and chemotherapy, the 5-year survival rate of patients with nonmetastatic OS has reached 60–70%. 3 However, 15–20% of OS patients have metastatic OS, and the 5-year survival rate for metastatic patients is less than 30%. 4 Therefore, exploring new biomarkers for early diagnosis of OS is essential to improve the survival rate in OS patients.
Ether à go-go 1 (Eag1, KCNH1) potassium channel belongs to the voltage-gated potassium channel family. 5 KCNH1 is mainly distributed in the central nervous system and neoplasm cells and is a potential target for the diagnosis and treatment of various cancers. 5,6 Previous studies have shown KCNH1 to be a potential early marker for cervical, colon, and breast cancer. 7 Moreover, the Eag1 channel plays a vital role in tumorigenesis and progression. 8 In OS, Eag1 knockdown suppressed tumor development by inhibiting cell proliferation and cell cycle. 9
Circular RNAs (circRNAs) are a class of noncoding RNAs (ncRNAs), which are covalently closed endogenous biomolecules different from linear RNA. 10 More and more investigations have confirmed that circRNAs play a crucial role in various cancers, including OS. For example, Zheng et al. revealed that circRNA LRP6 facilitated OS progression by downregulating KLF2 and APC expression. 11 Yanbin et al. found that circSAMD4A expedited cell proliferation in OS via the microRNA-1244/MDM2 axis. 12 In addition, previous studies confirmed that KCNH1 was overexpressed in OS and induced OS proliferation and metastasis. 9,13 A recent study disclosed that circ_0016347 derived from KCNH1 transcript was a tumor-promoting factor in OS. 14 However, the molecular mechanism of circ_0016347 needs to be further studied.
MicroRNAs (miRNAs) are highly conserved endogenous ncRNAs, consisting of 18–24 nucleotides. 15 Accumulating evidence has validated that miRNA dysregulation plays a vital role in the development of cancer. 16 Besides, many circRNAs exert crucial regulatory functions in a variety of biological processes by acting as miRNA molecular sponges or competitive endogenous RNAs (ceRNAs). 17,18 For instance, circ-SERPINE2 sponged miR-375 and regulated YWHAZ expression to expedite gastric carcinoma progression. 19 CircDOCK1 contributed to the malignancy of bladder carcinoma by sponging miR-132-3p and increasing Sox5 expression. 20 CircFBLIM1 and FBLIM1 acted as ceRNAs to boost hepatocellular cancer development through regulation of miR-346. 21 Nevertheless, the relationship between circ_0016347 and miR-1225-3p has not been investigated.
In the present research, we detected the expression of circ_0016347 and KCNH1 in OS tissues and cells. Further, we explored the function and potential mechanism of circ_0016347 in OS. These findings indicated that circ_0016347 might be a potential biomarker for OS therapy.
Materials and Methods
Specimen collection
Fifty OS samples and paired adjacent normal tissues were collected from The Second Affiliated Hospital, University of South China. None of the patients received preoperative treatment. This study was authorized by the Ethics Committee of The Second Affiliated Hospital, University of South China. All participants signed written informed consent. Some clinical characteristics of OS patients are listed in Table 1.
Correlation Between KCNH1 Expression and Clinicopathological Characteristics in Osteosarcoma
p < 0.05.
Cell culture and treatment
OS cell lines (U2OS, HOS, SaoS2, and MG63) and human fetal osteoblastic cells (hFOB1.19) were obtained from American Type Culture Collection (ATCC, Manassas, VA). Cells were incubated at 37°C in Dulbecco's modified Eagle's medium (DMEM; Solarbio, Shanghai, China) supplemented with 10% fetal bovine serum (FBS; Solarbio). For Actinomycin D assay, hFOB1.19 cells were exposed to 2 μg/mL Actinomycin D (Solarbio) to block transcription for 0, 6, 12, 18, and 24 h.
Cell transfection
KCNH1 overexpression vector pcDNA3.1-KCNH1 (KCNH1), circ_0016347 overexpression vector pCD5-ciR-circ_0016347 (circ_0016347), the empty overexpression vector (vector), small interfering RNA (siRNA) targeting KCNH1 (si-KCNH1; 5′-CTCTATTTCCTATTAAAGT-3′), siRNA against circ_0016347 (si-circ_0016347), the siRNA negative control (si-NC), miR-1225-3p mimic (miR-1225-3p), the mimic control (miR-NC), miR-1225-3p inhibitor (anti-miR-1225-3p), and the inhibitor control (anti-miR-NC) were synthesized by Ribobio (Guangzhou, China). The vectors and oligonucleotides were transfected into OS cells at 70% confluence using Lipofectamine 2000 (Invitrogen, Carlsbad, CA).
Quantitative real-time polymerase chain reaction
RNA extraction was carried out using TRIzol reagent (Invitrogen). For RNase R digestion, 2 μg of RNA was incubated for 30 min at 37°C with or without RNase R (3 U/μg) (Sigma, St Louis, MO) and then purified using RNeasy MinElute Cleanup Kit (Qiagen, Frankfurt, Germany). Then, 2 μg of RNA was reverse-transcribed using M-MLV RT Kit (AiYou Biosciences, Guangzhou, China) or miScript II RT Kit (Qiagen). The expression was monitored by SYBR Green PCR Master Mix (LMAI Bio, Shanghai, China) and calculated using the 2−ΔΔCt method. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) or U6 small RNA was used as the internal control. The primer sequences were as follows: circ_0016347-F: 5′-CCCGAGGCCAGTTTCCTG-3′, circ_0016347-R: 5′-TTAGTATCATTGGACCGCCGA-3′; miR-1225-3p-F: 5′-TGAGCCCCTGTGCCGC-3′, miR-1225-3p-R: 5′-TGGTGTCGTGGAGTCG-3′; KCNH1-F: 5′-AGTGGCCCCTCAAAACACG-3′, KCNH1-R: 5′-CTATCTGAGCATTCCCCAACAC-3′; GAPDH-F: 5′-ACAACTTTGGTATCGTGGAAGG-3′, GAPDH-R: 5′-GCCATCACGCCACAGTTTC-3′; U6-F: 5′-CTCGCTTCGGCAGCACA-3′, U6-R: 5′-AACGCTTCACGAATTTGCGT-3′.
Western blot assay
Cells were lysed using RIPA buffer (Solarbio). Proteins were quantified by the BCA™ Protein Assay Kit (Pierce, Appleton, WI). Then, the same amount of proteins was subjected to sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) to separate proteins. Subsequently, the proteins were transferred onto polyvinylidene fluoride membranes (Millipore, Billerica, MA). The membranes were blocked with 5% skim milk for 2 h, and then incubated with primary antibodies against KCNH1 (ab101174; Abcam, Cambridge, United Kingdom), Ki-67 (ab15580; Abcam), matrix metalloproteinase-9 (MMP-9)(ab38898; Abcam), hexokinase-2 (HK2) (ab227198; Abcam), and GAPDH (ab9485; Abcam) overnight at 4°C. Next, the membranes interacted with the corresponding secondary antibody (ab7090; Abcam) for 2 h at room temperature. Subsequently, the signal intensity was detected by the enhanced chemiluminescence system (Qiagen).
MTT assay
Cells (2.0 × 103) were seeded into 96-well plates. Next, 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide (MTT) solution (Solarbio) was added to each well after incubation for 0, 24, 48, and 72 h. Then, cells were treated with dimethyl sulfoxide (DMSO; Solarbio) for 15 min. Finally, the value of optical density was evaluated by monitoring the absorbance at 490 nm using a Microplate Reader (Bio-Rad, Hercules, CA).
Colony formation assay
The transfected cells were plated into six-well plates covered with DMEM medium carrying 10% FBS to culture 14 d at 37°C. Colonies were fixed with methanol and stained with 0.5% crystal violet. Then, the number of colonies was imaged and counted in at least three independent fields of view.
Transwell assay
For cell migration assay, cells were placed in the upper chamber with a serum-free medium. Besides, 10% FBS was added to the lower chamber. The migrated cells were treated with methanol and stained with crystal violet for 20 min after incubation for 24 h. Then, the cells were counted using a microscope in three randomly selected fields. For cell invasion assay, transwell chambers were coated with Matrigel (BD Biosciences, San Diego, CA), and the other procedures were the same as the migration assay.
Detection of glucose consumption and lactate production
Cells were seeded into six-well plates. Then, the medium was replaced with fresh complete medium and incubated for another 48 h. The glucose consumption and lactate production were assessed using glucose detection and lactic acid detection kits (Zhongsheng Biotech, Beijing, China).
Dual-luciferase reporter assay
U2OS and MG63 cells were seeded in 24-well plates. The sequences of circ_0016347 or KCNH1–3′UTR containing wild-type or mutant binding sites of miR-1225-3p were inserted into pmirGLO vector (Promega, Madison, WI). The corresponding luciferase reporter and miR-1225-3p mimic or miR-NC were co-transfected into OS cells. Finally, the luciferase activity was examined by Dual-Lucy Assay Kit (Solarbio).
Lentivirus infection
Lentivirus vectors containing short hairpin RNA (shRNA) against circ_0016347 (sh-circ_0016347; 5′-GATGAATTCCTTTGAAATT-3′) or the negative control (sh-NC) were constructed in Genechem (Shanghai, China). One × 106 TU/mL lentivirus with Polybrene was infected into MG63 cells at 70% confluence. Subsequently, transfected cells were selected with puromycin.
Xenograft tumor experiment
BALB/c nude mice (6 weeks old) were randomly divided into two groups (n = 6 per group). MG63 cells were infected with lentivirus harboring sh-NC or sh-circ_0016347, respectively. Then, cells (5 × 106) were subcutaneously injected into the right-back of nude mice. Tumor volume was measured every 7 d. The mice were sacrificed after 28 d, and the excised tumors were photographed and weighed. The xenograft experiment was authorized by the Animal Welfare Committee of The Second Affiliated Hospital, University of South China.
Statistical analysis
Graphpad Prism 7.0 software (GraphPad, San Diego, CA) was executed to analyze data. Data were expressed as mean ± standard deviation of three independent experiments. Differences were assessed using Student's t-test or one-way analysis of variance. p < 0.05 was considered statistically significant.
Results
Circ_0016347 and KCNH1 were upregulated in OS tissues and cells
To investigate whether circ_0016347 was dysregulated in OS, we first examined the expression of circ_0016347 in OS tissues and matched adjacent tissues. The results exhibited that the levels of circ_0016347 and KCNH1 were distinctly increased in OS tissues compared with adjacent tissues (Fig. 1A, B). Circ_0016347 expression was positively correlated with KCNH1 expression in OS tissues (Fig. 1C). Additionally, the protein level of KCNH1 in OS tissues was remarkably elevated in comparison with adjacent tissues (Fig. 1D). The quantitative real-time polymerase chain reaction (qRT-PCR) detection confirmed that circ_0016347 had no ploy-A tail (Fig. 1E). Compared to KCNH1, circ_0016347 was resistant to RNase R (Fig. 1F). To test the stability of circ_0016347, hFOB1.19 cells were treated with the transcription inhibitor Actinomycin D and then analyzed by qRT-PCR. The results revealed that circ_0016347 had a half-life of more than 24 h, while its linear isoform KCNH1 mRNA had a half-life of less than 18 h (Fig. 1G). Moreover, the expression of circ_0016347 and KCNH1 were notably higher in OS cells (U2OS, HOS, SaoS2, and MG63) than that in hFOB1.19 cells (Fig. 1H–J). We also investigated some clinical features of patients with OS, and the data showed that KCNH1 expression was not related to gender, histological grade, lymphatic metastasis, and distant metastasis, but was associated with age, TNM stages, tumor size, and invasion depth (Table 1). Taken together, these results indicated that circ_0016347 was a highly stable and dysregulated circRNA in OS.
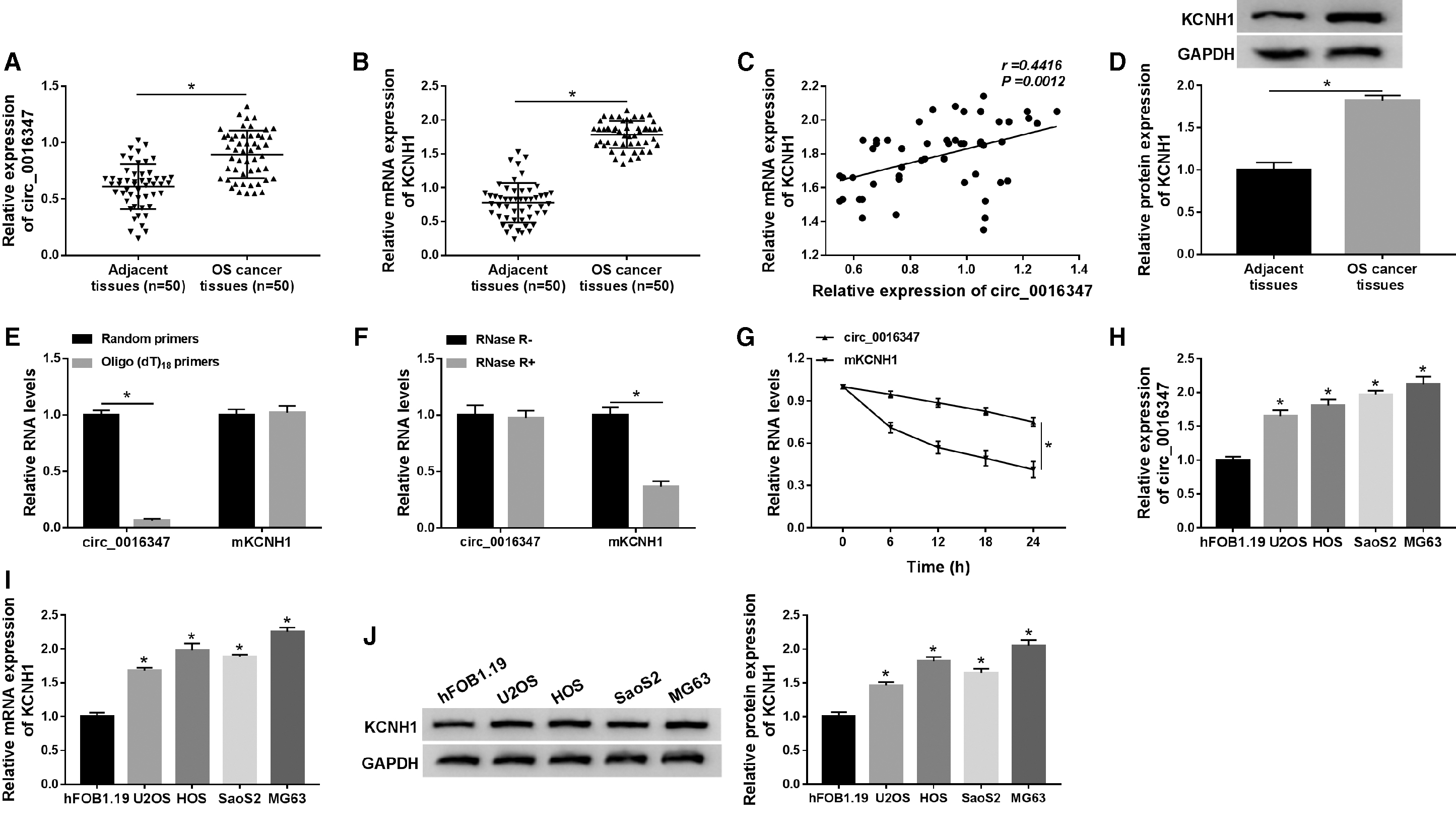
Circ_0016347 and KCNH1 were upregulated in OS tissues and cells.
KCNH1 facilitated cell proliferation, migration, invasion, and glycolysis in OS cells
To explore the effect of KCNH1 on OS progression, KCNH1 or si-KCNH1 was transfected into U2OS or MG63 cells, respectively. The results revealed that U2OS cells with high expression of KCNH1 were established by transfection with KCNH1, and MG63 cells with low expression of KCNH1 were constructed by introduction with si-KCNH1 (Fig. 2A, B). MTT assay exhibited that KCNH1 overexpression promoted the proliferation of U2OS cells, and the proliferation of MG63 cells transfected with si-KCNH1 was repressed (Fig. 2C, D). Colony formation assay suggested that the colony number of U2OS cells was strikingly increased, while introduction of si-KCNH1 observably reduced colony number (Fig. 2E). Transwell assay showed that cell migration and invasion were remarkably promoted after transfection with KCNH1 and hindered after introduction with si-KCNH1 (Fig. 2F, G). In addition, upregulation of KCNH1 overtly increased the levels of glucose consumption and lactate production in U2OS cells, whereas knockdown of KCNH1 conspicuously reduced the levels in MG63 cells (Fig. 2H, I). Moreover, Western blot analysis indicated that transfection with KCNH1 led to a significant increase in Ki-67, MMP-9, and HK2 levels, while KCNH1 depletion obviously reduced Ki-67, MMP-9, and HK2 levels (Fig. 2J, K). All these data demonstrated that KCNH1 contributed to proliferation, migration, invasion, and glycolysis of OS cells.
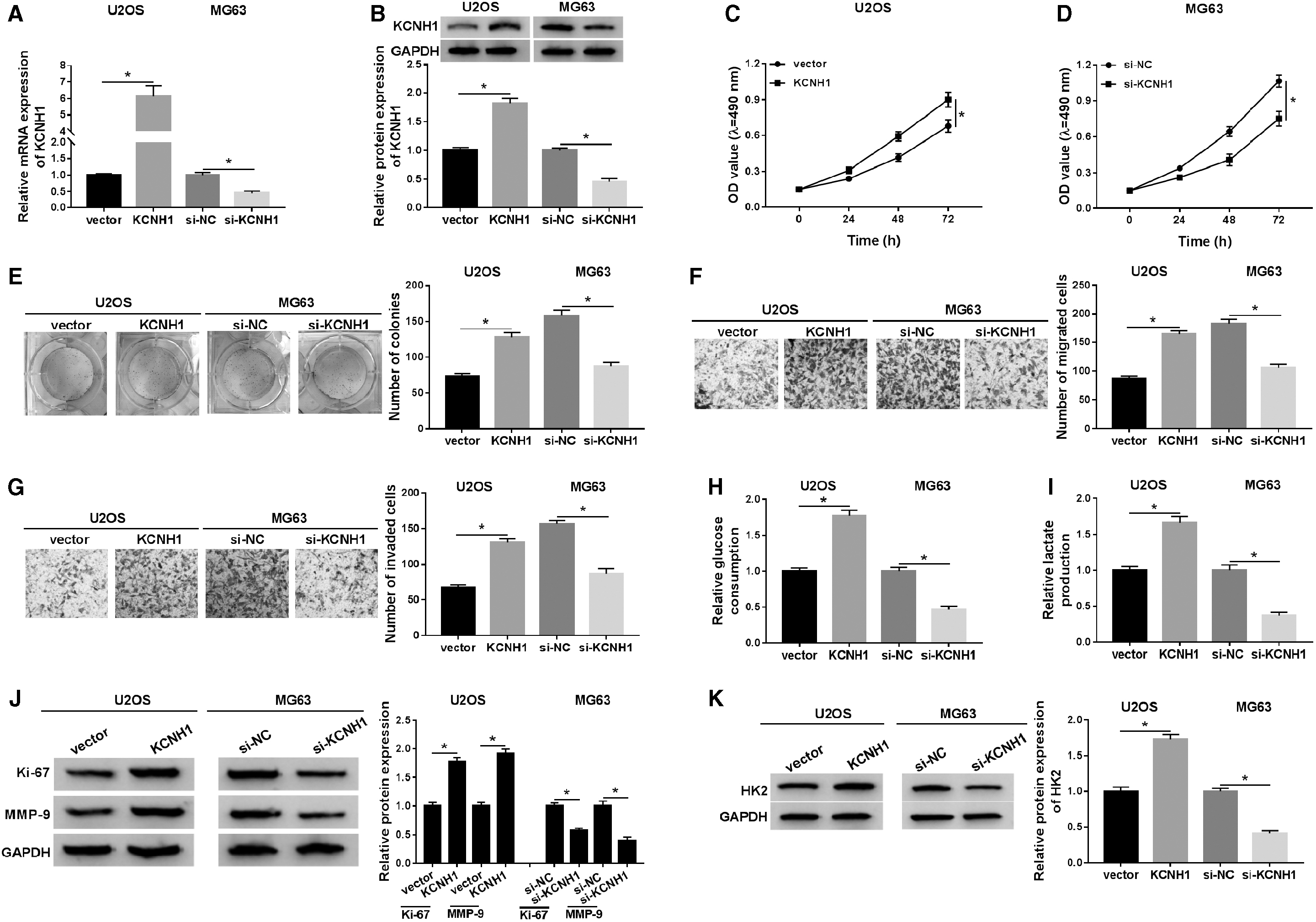
KCNH1 facilitated cell proliferation, migration, invasion, and glycolysis in OS cells. U2OS cells were introduced with vector or KCNH1, and MG63 cells were transfected with si-NC or si-KCNH1.
Aberrant expression of circ_0016347 modulated the progression of OS
Sequence alignment results showed that circ_0016347 was derived from exon 6 to exon 7 of linear KCNH1, and its full length was 904 bp (Fig. 3A). To investigate the effect of circ_0016347 on OS progression, U2OS or MG63 cells were introduced with circ_0016347 or si-circ_0016347, and transfection efficiency was determined by qRT-PCR (Fig. 3B). In addition, upregulation of circ_0016347 prominently promoted cell proliferation (Fig. 3C, D), colony formation (Fig. 3E), and cell migration and invasion (Fig. 3F, G) in U2OS cells, whereas downregulation of circ_0016347 had the opposite effect on MG63 cells. Furthermore, transfection with circ_0016347 promoted glycolysis and resulted in a distinct increase in the protein levels of Ki-67, MMP-9, and HK2 compared with the vector group, while transfection of si-circ_0016347 had the opposite effect on OS cells (Fig. 3H–K). These data indicated that circ_0016347 had a positive effect on OS progression.
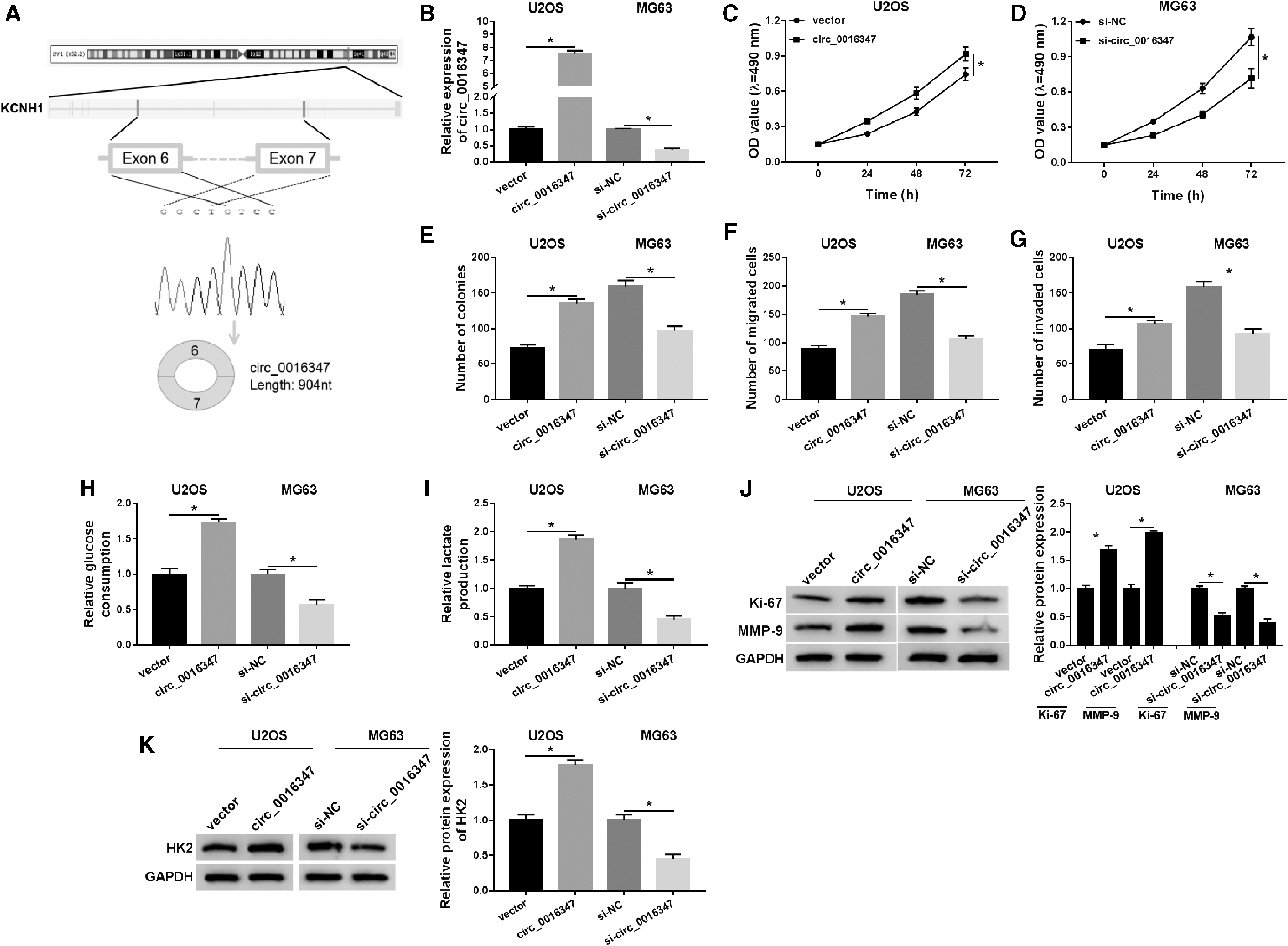
Aberrant expression of circ_0016347 modulated the progression of OS.
Circ_0016347 facilitated OS progression by regulating KCNH1
First, transfection of si-KCNH1 in U2OS cells resulted in an apparent reduction in the mRNA and protein expression of KCNH1, and introduction with KCNH1 in MG63 cells led to a significant increase in KCNH1 expression (Fig. 4A, B). MTT assay suggested that circ_0016347 overexpression increased cell proliferation ability, while co-transfection with circ_0016347 and si-KCNH1 relieved the effect (Fig. 4C). Besides, circ_0016347 knockdown remarkably restrained cell proliferation, whereas the effect was attenuated by upregulating KCNH1 (Fig. 4D). Moreover, introduction of circ_0016347 distinctly induced colony number, cell migration and invasion, and glycolysis, but the effect was rescued after transfection with si-KCNH1 (Fig. 4E–I). Also, KCNH1 overexpression reversed the inhibitory effect of circ_0016347 silencing on colony formation, cell migration and invasion, and glycolysis (Fig. 4E–I). Furthermore, knockdown of KCNH1 abrogated the stimulatory effect on Ki-67, MMP-9, and HK2 levels induced by circ_0016347 overexpression (Fig. 4J, L). Upregulation of KCNH1 undermined the inhibitory effect on Ki-67, MMP-9, and HK2 levels caused by circ_0016347 depletion (Fig. 4K, L). These data certified that circ_0016347 facilitated OS progression through KCNH1 in OS cells.
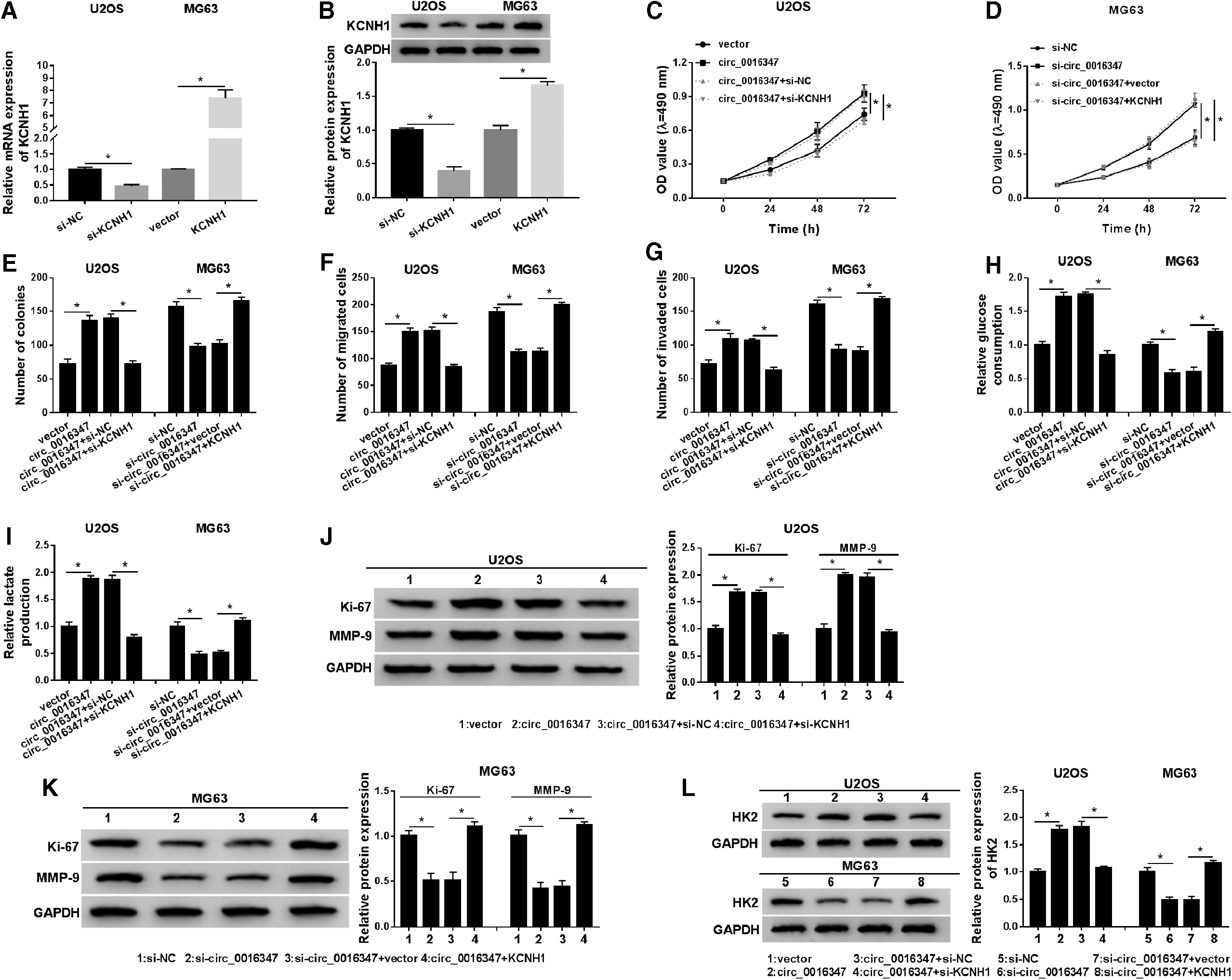
Circ_0016347 facilitated OS progression by regulating KCNH1.
Circ_0016347 was a sponge of miR-1225-3p, and miR-1225-3p targeted KCNH1 in OS cells
We used bioinformatics analysis and dual-luciferase reporter assay to elucidate the underlying mechanism of circ_0016347. First, the online database circRNA Interactome predicted that circ_0016347 had putative binding sites with miR-1225-3p, and TargetScan predicted that KCNH1 might be a target of miR-1225-3p (Fig. 5A). Dual-luciferase reporter assay revealed that miR-1225-3p mimic overtly reduced the luciferase activity of WT-circ_0016347 reporter, but did not affect the luciferase activity of MUT-circ_0016347 reporter (Fig. 5B, C). Also, the luciferase activity of WT-KCNH1–3′UTR reporter was strikingly decreased after transfection with miR-1225-3p mimic (Fig. 5D, E). Moreover, miR-1225-3p expression was markedly reduced in U2OS and MG63 cells compared with hFOB1.19 cells (Fig. 5F). After transfection with si-circ_0016347, circ_0016347 expression in U2OS cells was conspicuously decreased, while circ_0016347 expression in MG63 cells was prominently increased after transfection with circ_0016347 (Fig. 5G). Moreover, circ_0016347 overexpression significantly decreased miR-1225-3p expression, while circ_0016347 knockdown elevated miR-1225-3p expression in U2OS and MG63 cells (Fig. 5H, I). MiR-1225-3p mimic evidently increased miR-1225-3p expression, and miR-1225-3p inhibitor strikingly reduced miR-1225-3p expression (Fig. 5J, K). The mRNA and protein levels of KCNH1 were dramatically decreased in OS cells transfected with miR-1225-3p compared to the miR-NC group and increased in the anti-miR-1225-3p group compared with the anti-miR-NC group (Fig. 5L–O). These data indicated that circ_0016347 was a sponge of miR-1225-3p, and KCNH1 was a target of miR-1225-3p in OS cells.
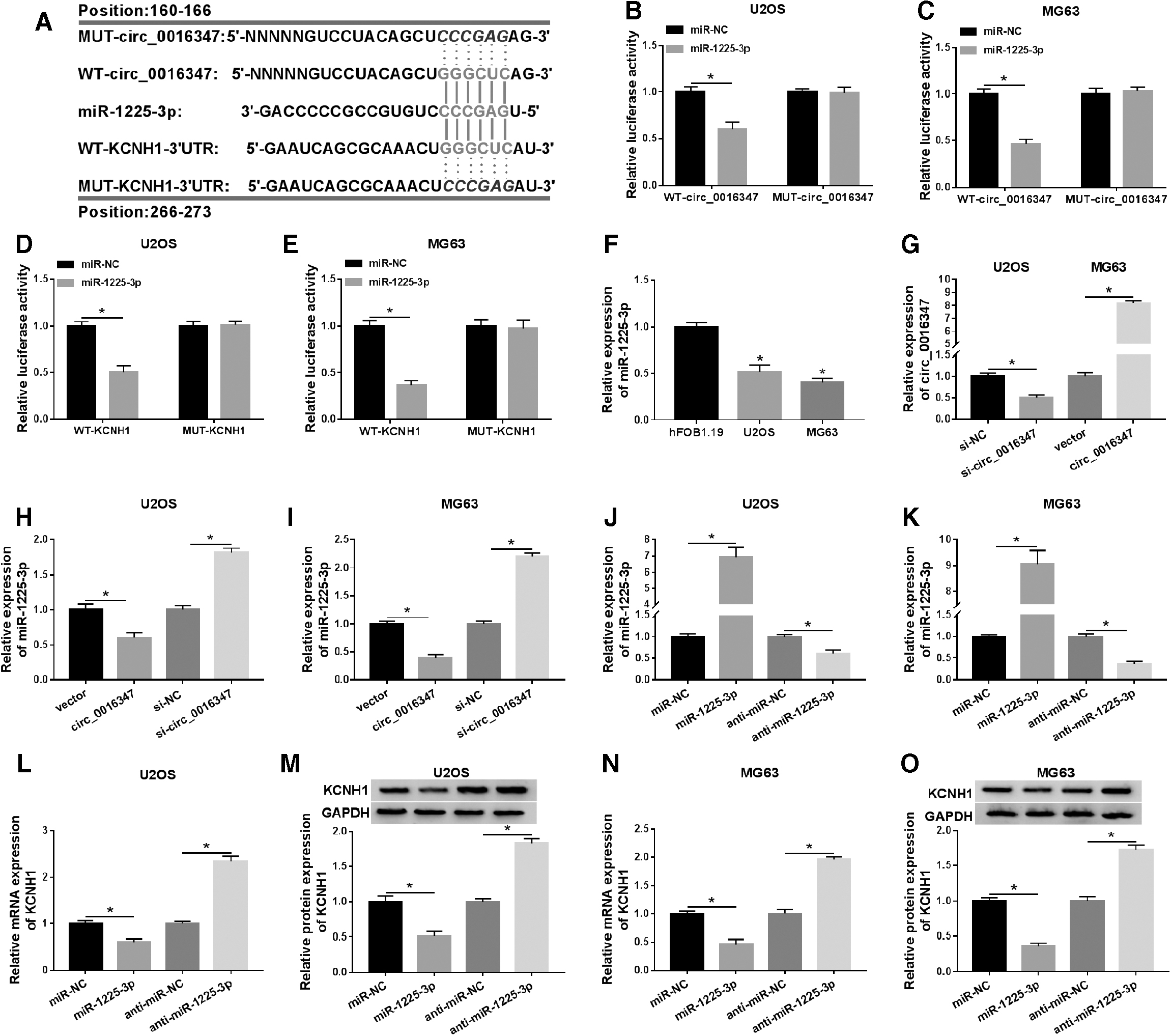
Circ_0016347 was a sponge of miR-1225-3p, and miR-1225-3p targeted KCNH1 in OS cells.
Circ_0016347 regulated KCNH1 expression by sponging miR-1225-3p
To explore whether circ_0016347 regulated KCNH1 expression by sponging miR-1225-3p, a rescue experiment was performed in U2OS and MG63 cells introduced with miR-NC, miR-1225-3p, miR-1225-3p+vector, or miR-1225-3p+circ_0016347. The results of qRT-PCR and Western blot showed that transfection with miR-1225-3p led to a noticeable decrease in the mRNA and protein expression of KCNH1, which was abolished after co-transfection with miR-1225-3p and circ_0016347 (Fig. 6A–D). These data indicated that circ_0016347 sponged miR-1225-3p to upregulate KCNH1 expression in OS cells.
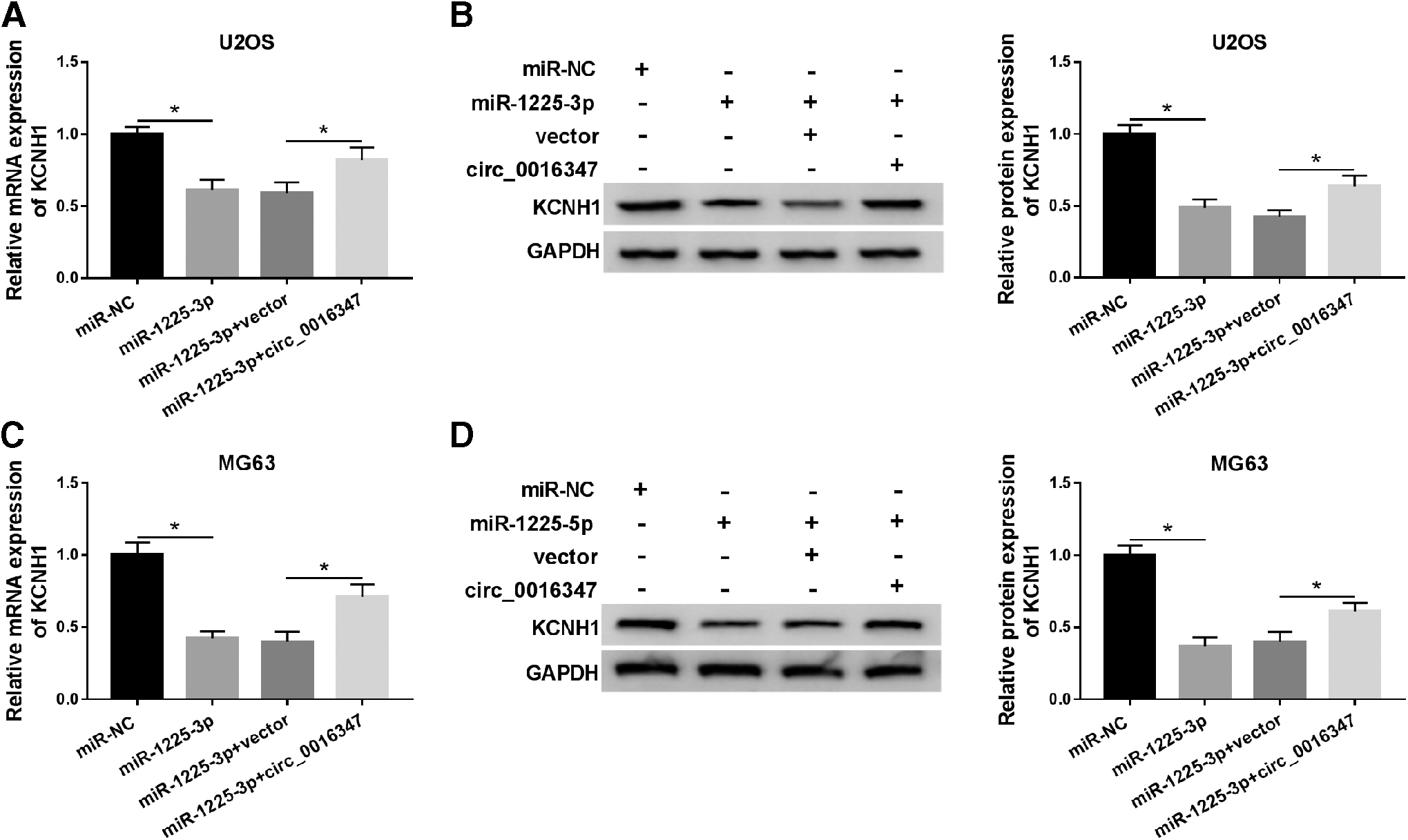
Circ_0016347 regulated KCNH1 expression by sponging miR-1225-3p. U2OS and MG63 cells were transfected with miR-NC, miR-1225-3p, miR-1225-3p+vector, or miR-1225-3p+circ_0016347.
Knockdown of circ_0016347 blocked tumor growth in vivo
To investigate the role of circ_0016347 in OS tumorigenesis in vivo, we established a xenograft mice model. The results exhibited that the tumor volume and weight in the sh-circ_0016347 group were remarkably lower than those in the sh-NC group (Fig. 7A, B). In addition, circ_0016347 expression was evidently reduced, and miR-1225-3p expression was distinctly increased in the sh-circ_0016347 group compared with the sh-NC group (Fig. 7C, D). Immunohistochemistry, qRT-PCR, and Western blot assays exhibited that circ_0016347 silencing observably inhibited KCNH1 expression (Fig. 7E–G). These data certified that circ_0016347 knockdown hindered tumor growth in vivo.
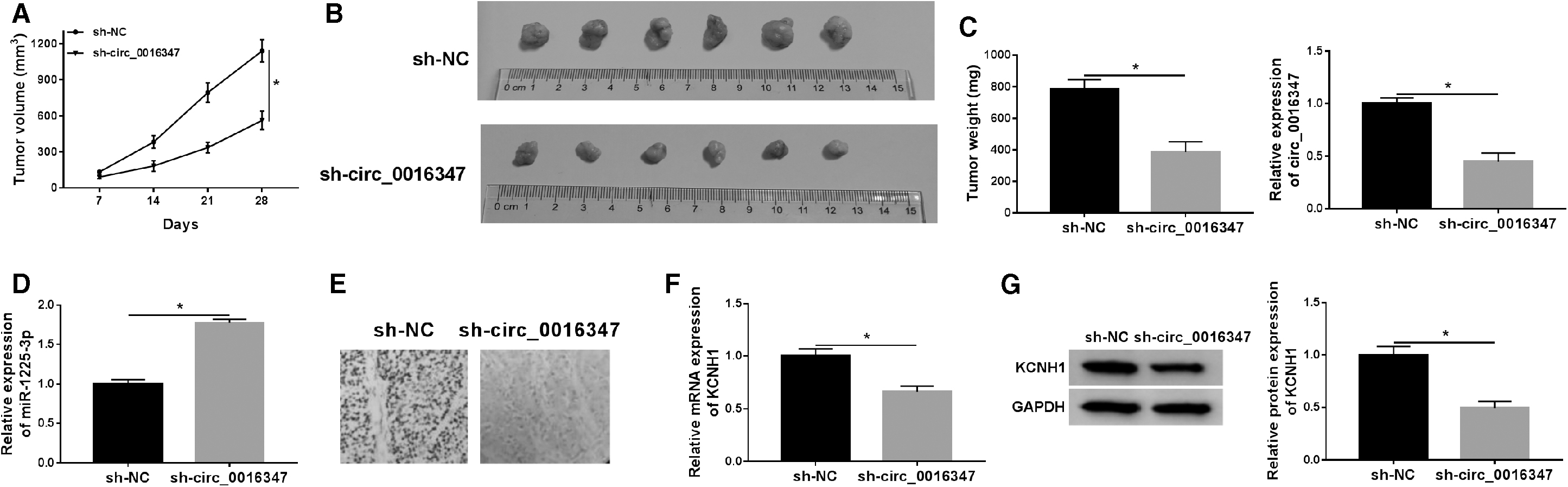
Knockdown of circ_0016347 blocked tumor growth in vivo. MG63 cells stably transfected with sh-NC or sh-circ_0016347 were subcutaneously inoculated into nude mice.
Discussion
Although surgery and radiochemotherapy have improved over the past few decades, the survival rate of OS patients has not increased as expected. Hence, elucidating the precise mechanism of OS occurrence and development is essential for ameliorating the prognosis of OS. 22 Besides, hexokinase (HKs) catalyzes the first step of glucose metabolism, and HK2 is an important indicator of glycolysis in tumors. 23 Numerous studies have shown that many lncRNAs and miRNAs can regulate OS progression. 24 Emerging evidence has demonstrated that aberrantly expressed circRNAs play a vital role in tumorigenesis and development. 17 For example, Jin et al. presented that abnormally expressed circRNA_100876 curbed cell proliferation in OS by acting as a sponge of microRNA-136. 25 Wu et al. unveiled that circTADA2A functioned as a sponge for miR-203a-3p to facilitate OS progression by upregulating CREB3. 26 Qi et al. found that circ_0000502 was firmly related to the unfavorable prognosis of OS and accelerated OS progression by binding to miR-1238. 27 Additionally, circ_0016347 was identified as an oncogene in OS. 14 The results of this study were consistent with previous studies. Circ_0016347 expression was overtly increased in OS tissues and cells. Also, circ_0016347 facilitated OS progression.
In the current study, we corroborated that miR-1225-3p was a target of circ_0016347. Interestingly, miR-1225 has been reported as a modulator of multiple malignancies. In pancreatic cancer, miR-1225 was upregulated and inhibited cell apoptosis through degradation of JAK1. 28 In thyroid cancer, miR-1225-5p targeted sirtuin 3 (SIRT3) to suppress tumor progression. 29 In glioblastoma, miR-1225-5p acted as a tumor-suppressing factor by downregulating the expression of insulin receptor substrate 1 (IRS1). 30 Moreover, Sun et al. discovered that miR-1225-5p triggered cell cycle arrest in laryngeal cancer by regulating CDC14B. 31 In biliary tract cancer and pheochromocytomas, miR-1225-3p was considered to be a predictor of tumor recurrence. 32,33 However, the function and mechanism of miR-1225-3p in OS have not been studied. In this study, we first evidenced that miR-1225-3p was conspicuously reduced in OS cells. Moreover, circ_0016347 was a sponge of miR-1225-3p in OS cells.
KCNH1 is a member of the voltage-activated potassium channel family and has high expression in more than 70% of human tumors. 8 KCNH channels implicated in brain developmental deficits and cancers by serving as an emerging oncological target. 34 In ovarian cancer, depletion of KCNH potentiated the cisplatin sensitivity of SKOV3 and TYK cells. 35 In addition, KCNH knockdown impaired OS progression through deactivation of STAT3-VEGF pathway. 36 MicroRNA-34a impeded the proliferation of OS cells via repressing KCNH expression. 37 In this study, KCNH expression in OS tissues and cells was drastically elevated, which was consistent with previous results. More importantly, restoration experiments demonstrated that circ_0016347 facilitated OS progression by affecting KCNH1.
Conclusion
In conclusion, our study concluded that circ_0016347 functioned as a sponge of miR-1225-3p to modulate KCNH1 expression. Moreover, functional experiments evidenced that circ_0016347 facilitated OS progression by regulating miR-1225-3p/KCNH1 pathway, which might provide a novel therapeutic strategy for OS patients. Nevertheless, there are some limitations in this study. The effect of miR-1225-3p overexpression on circ_0016347 knockdown or the effect of KCNH1 silencing on miR-1225-3p overexpression should be supplemented in rescue experiments.
Footnotes
Disclosure Statement
The authors declare that they have no financial conflicts of interest.
Funding Information
This study was supported by the Clinical Medical Technology Innovation Guidance Project of Hunan Province, Research on Treatment Model of Severe Trauma (Grant No. 2018SK51507).
Supplementary Material
KCNH1 Western blots