
Abstract
Background:
Cervical cancer (CC) is a common gynecological malignancy with a high risk of recurrence and death. Circular RNAs play a crucial role in the occurrence and development of tumors. This study aimed to investigate the function and mechanism of circ_0000745 in CC.
Materials and Methods:
The levels of circ_0000745, miR-409-3p, and activating transcription factor 1 (ATF1) were determined by quantitative real-time polymerase chain reaction or Western blot assay. Cell proliferation was assessed by colony formation assay. Cell migration and invasion were evaluated by transwell assay. Glycolysis was analyzed by measuring extracellular acidification rate, glucose uptake, and lactate production. Also, the protein levels of glucose transporter 1 and lactate dehydrogenase A were detected using Western blot. The relationship among circ_0000745, miR-409-3p, and ATF1 were confirmed by dual-luciferase reporter assay. Moreover, xenograft assay was performed to analyze tumor growth in vivo.
Results:
Circ_0000745 and ATF1 were upregulated, whereas miR-409-3p was downregulated in CC tissues and cells. Knockdown of circ_0000745 repressed proliferation, migration, invasion, and glycolysis of CC cells. Circ_0000745 regulated CC progression by targeting miR-409-3p. Circ_0000745 modulated ATF1 expression through sponging miR-409-3p. MiR-409-3p hindered CC progression by targeting ATF1. Furthermore, depletion of circ_0000745 impeded tumor growth in vivo.
Conclusion:
Circ_0000745 promoted the progression of CC through modulating miR-409-3p/ATF1 axis, indicating a promising biomarker for CC therapy.
Introduction
Cervical cancer (CC) ranks fourth in cancer incidence and mortality. 1 Human papillomavirus (HPV) infection is known to be a significant risk factor for CC. 2 With the implementation of screening and HPV vaccination programs, the incidence and mortality of CC have been significantly lower. 3 However, for patients with metastatic or recurrent CC, the overall prognosis is still poor. 4 Therefore, it is urgent to explore new therapeutic targets for improving survival from CC.
Circular RNAs (circRNAs) are an unusual class of noncoding RNAs (ncRNAs), which are formed by cyclization of exon or intron at the 3′ and 5′ ends. 5 In recent years, increasing evidence has corroborated that circRNAs regulate gene transcription by acting as microRNA sponges, thus emerging as a regulatory role in various biological and pathological processes. 6 Also, the aberrant expression of circRNAs is associated with the occurrence and development of cancer. 7 For example, hsa_circ_0004491 knockdown facilitates cell metastasis in oral squamous cell carcinoma. 8 Silence of circPVT1 impedes the invasion of esophageal carcinoma cells. 9 CircLMNB1 becomes a promising therapeutic target by promoting colorectal cancer progression. 10 In addition, circRNA expression profiles have been studied in CC using high-throughput RNA sequencing technology. 11 Besides, circ_0000745 is derived from sperm antigen with calponin homology and coiled-coil domains 1 (SPECC1). A recent study detected abnormal expression of circRNAs in CC using circRNA microarrays and found that circ_0000745 contributed to CC progression. 12 Nevertheless, the specific mechanism of circ_0000745 has not been elucidated.
MicroRNAs (miRNAs) are a group of small ncRNAs that regulate the function of mRNA by binding to mRNA 3′UTR. 13 A large number of miRNAs exert a crucial role in CC progression. 14 For instance, miR-139-5p targets TCF4 to block CC progression. 15 MiR-9-5p attenuates the radioresistance of CC cells by binding to SOCS5. 16 Previous research indicated that miR-409-3p was lowly expressed in CC. 17 However, the effect of miR-409-3p in CC is unclear.
Activating transcription factor 1 (ATF1) belongs to the ATF subfamily and contains a basic zipper (bZIP). 18 ATF1 participates in various cellular processes by regulating the transcription of target genes. 19 For example, ATF1 induces the metastasis of lung cancer by increasing EGFR and MMP-2 expression. 20 Nevertheless, the function of ATF1 in CC is unclear.
In the present research, the expression of circ_0000745, miR-409-3p, and ATF1 in CC tissues and cells was investigated. Also, the authors explored the function and regulatory mechanism of circ_0000745 in CC cells. These findings unveiled that circ_0000745 functioned as a competing endogenous RNA (ceRNA) to modulate ATF1 through sponging miR-409-3p in CC.
Materials and Methods
Specimen collection
Fifty-five pairs of CC tissues and adjacent nontumor tissues were obtained from patients with CC who underwent surgical resection at Changzhou No. 2 People's Hospital. All participants signed written informed consent. This study obtained the approval of the Ethics Committee of Changzhou No. 2 People's Hospital. Clinical staging of CC patients was performed according to the International Federation of Gynecology and Obstetrics (FIGO) staging system.
Cell culture
Four CC cell lines (HeLa, Caski, C33A and Siha) and human normal ectocervical epithelial cell line ECT1/E6E7 were obtained from American Type Culture Collection (ATCC, Manassas, VA, USA). Caski cells were cultured in RPMI-1640 medium (Gibco, Carlsbad, CA, USA) with 10% fetal bovine serum (FBS; Gibco), and other cell lines were cultured in Dulbecco's modified Eagle's medium (Gibco) supplemented with 10% FBS.
Quantitative real-time polymerase chain reaction
RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA). Then, RNA was reverse transcribed using the PrimeScript™ RT Reagent Kit (TaKaRa, Dalian, China) or miScript II RT Kit (Qiagen, Frankfurt, Germany). The expression level was tested using TB Green Fast qPCR Mix (TaKaRa). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) or U6 small RNA was used as the internal control. Relative expression levels were quantified using 2−ΔΔCt method. The primer sequences were as follows: circ_0000745-F: 5′-ATGTTGAAAGTAGCCCGAGCAG-3′, circ_0000745-R: 5′-TGGGAGTGTTGGAAGAAGTTGG-3′; SPECC1-F: 5′-ACCCCAGGAAATCAGTGTCCA-3′, SPECC1-R: 5′-GTTCCCGAACTTGGGACTCAA-3′; miR-409-3p-F: 5′-GCGAATGTTGCTCGGTGA-3′, miR-409-3p-R: 5′-GTGCAGGGTCCGAGGT-3’; ATF1-F: 5′-AGGACTCATCCGACAGCATAG-3′, ATF1-R: 5′-TTCTGCCCCGTGTATCTTCAG-3′; GAPDH-F: 5′-GGGAAACTGTGGCGTGAT-3′, GAPDH-R: 5′-GAGTGGGTGTCGCTGTTGA-3′; U6-F: 5′-CTCGCTTCGGCAGCACA-3′, U6-R: 5′-AACGCTTCACGAATTTGCGT-3′.
In situ hybridization
Tissue sections were dewaxed with xylene and rehydrated with gradient ethanol. After reacting with 3% H2O2 for 10 min at room temperature, pepsin containing 3% fresh citric acid was added and digested at 37°C for 10–25 min. Then, sections were incubated overnight in the hybridization solution with digoxin-labeled oligonucleotide probes. Next, according to the ISH Detection Kit (Leica Biosystems, Buffalo Grove, IL, USA), the blocking solution, biotinylated mouse anti-digoxin and SABC were added successively.
Isolation of RNA from nuclear and cytoplasmic fractions
The subcellular localization of circ_0000745 was determined using the mirVana™ PARIS Kit (Invitrogen). GAPDH and U6 were used as positive controls for cytoplasmic and nuclear fractions, respectively.
RNase R and actinomycin D treatment
For RNase R treatment, 2 μg RNA was incubated with or without RNase R (3 U/μg) (Geneseed, Guangzhou, China) for 30 min at 37°C. For actinomycin D treatment, Caski and Siha cells were exposed to 2 mg/mL Actinomycin D (Seebio, Shanghai, China) to restrict transcription.
Cell transfection
Small interfering RNA (siRNA) against circ_0000745 (si-circ_0000745), siRNA against ATF1 (si-ATF1), the siRNA control (si-NC), miR-409-3p mimics (miR-409-3p), the mimics control (miR-NC), circ_0000745 overexpression vector (circ_0000745), the empty overexpression vector (vector), miR-409-3p inhibitor (anti-miR-409-3p), and the inhibitor control (anti-miR-NC) were obtained from Ribobio (Guangzhou, China). Lipofectamine 3000 (Invitrogen) was used to transfect oligonucleotides and plasmids into CC cells.
Colony formation assay
The transfected cells were plated into six-well plates and cultured for 14 d at 37°C in a medium carrying 10% FBS. After fixing with methanol and staining with 0.5% Crystal Violet, the number of colonies was imaged and counted.
Transwell assay
Cells were injected into the upper chamber. At the same time, 10% FBS was added to the lower chamber. Subsequently, the migrated cells were stained with Crystal Violet for 20 min after incubation for 24 h. Then, the cells were counted using a microscope in three randomly selected fields. For cell invasion assay, the other steps were the same as cell migration assay, except that the upper chamber was coated with Matrigel (BD Biosciences, San Diego, CA, USA).
Detection of extracellular acidification rate
The extracellular acidification rate (ECAR) was detected using the Seahorse XFe 96 Extracellular Flux Analyzer (Seahorse Bioscience, North Billerica, MA, USA). Briefly, cells (2 × 104) were seeded into a Seahorse XF 96-cell culture microplate. After baseline measurements, glucose, oligomycin, and 2-deoxyglucose were sequentially added to each well at the indicated time points. The Seahorse XF-96 Wave software was used to analyze the data. ECAR is expressed in mpH/min. All values were normalized to protein concentration.
Measurement of glucose uptake and lactate production
Cells were seeded in six-well plates. The medium was replaced with fresh complete medium and incubated for another 48 h. Then, glucose uptake and lactate production were measured using the Glucose and Lactate Assay Kits (Sigma-Aldrich, St Louis, MO, USA).
Western blot assay
The cells were lysed using RIPA buffer (Solarbio, Beijing, China) and then subjected to sodium dodecyl sulfate/polyacrylamide gel electrophoresis (SDS-PAGE). Subsequently, the proteins were transferred to polyvinylidene fluoride membranes (Millipore, Billerica, MA, USA). After blocking with 5% skim milk for 2 h, the membranes were incubated with primary antibodies against glucose transporter 1 (GLUT1) (ab15309; Abcam, Cambridge, United Kingdom), lactate dehydrogenase A (LDHA) (ab84716; Abcam), ATF1 (ab189311; Abcam), or GAPDH (ab9485; Abcam) overnight at 4°C. Subsequently, the membranes were interacted with secondary antibody (ab7090; Abcam) for 2 h at room temperature. Finally, protein bands were visualized with the enhanced chemiluminescence system (Qiagen).
Dual-luciferase reporter assay
The sequences of circ_0000745 or ATF1-3′UTR containing wild-type or mutant binding sites of miR-409-3p were inserted into pmirGLO vector (Promega, Madison, WI, USA). The corresponding luciferase reporter and miR-409-3p mimics or miR-NC were cotransfected into CC cells. Then, the luciferase activity was examined by the Dual-Lucy Assay Kit (Solarbio).
Xenograft assay
Siha cells were infected with lentivirus containing short hairpin RNA targeting circ_0000745 (sh-circ_0000745) or the negative control (sh-NC) (HanBio, Shanghai, China). Next, stably transfected Siha cells (5 × 106) were subcutaneously injected into the right abdomen of BALB/c nude mice (4-week-old). Tumor volume was measured every week. Four weeks later, the xenografts were excised and weighed. The xenograft experiment was approved by the Animal Welfare Committee of Changzhou No. 2 People's Hospital.
Statistical analysis
GraphPad Prism 7.0 software (GraphPad, San Diego, CA, USA) was executed to analyze data. Data were shown as mean ± standard deviation. Differences were assessed using Student's t-test or one-way analysis of variance. All experiments were repeated three times independently. p < 0.05 was considered statistically significant.
Results
Circ_0000745 was upregulated in CC tissues and cells
To investigate the role of circ_0000745 in CC, the authors first examined the expression of circ_0000745 in CC tissues and paired adjacent tissues. The results demonstrated that circ_0000745 level in CC tissues was distinctly higher compared with adjacent tissues (Fig. 1A). Besides, circ_0000745 expression in patients with stage III+IV was remarkably higher compared with patients with stage I + II (Fig. 1B). Also, the level of circ_0000745 in the positive invasion group was overtly higher compared with the negative invasion group (Fig. 1C). The in situ hybridization results further confirmed the enhancement of circ_0000745 in CC tissues (Fig. 1D). In addition, circ_0000745 expression in CC cells (HeLa, Caski, C33A and Siha) was drastically raised in comparison with the normal ectocervical epithelial cell line ECT1/E6E7 (Fig. 1E). These data reflected that circ_0000745 might play an oncogene in CC.
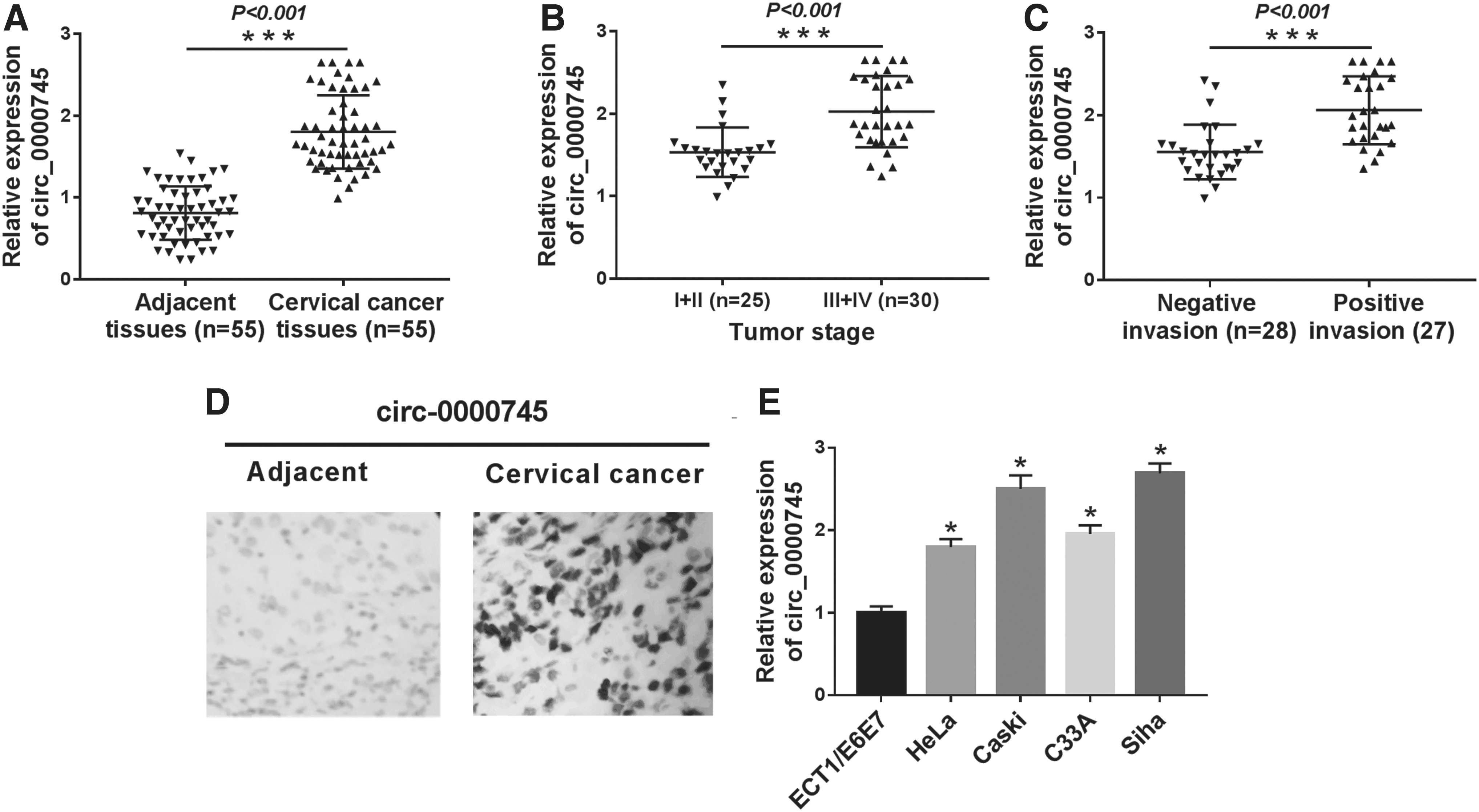
Circ _0000745 was upregulated in CC tissues and cells.
Identification of circ_0000745 in CC cells
First, they determined the subcellular localization of circ_0000745 in Caski and Siha cells by using nuclear and cytoplasmic fraction assay. The results suggested that circ_0000745 was mainly distributed in the cytoplasm (Fig. 2A). Also, the quantitative real-time polymerase chain reaction (qRT-PCR) detection disclosed that circ_0000745 had no poly-A tail (Fig. 2B). To detect the stability of circ_0000745 in CC cells, Caski and Siha cells were treated with or without RNase R. The results revealed that circ_0000745 was resistant to RNase R relative to its linear isoform SPECC1 (Fig. 2C). Similarly, after treatment with Actinomycin D, the half-life of circ_0000745 was prominently higher compared with SPECC1 (Fig. 2D). These data indicated that circ_0000745 is a highly stable circRNA in CC.
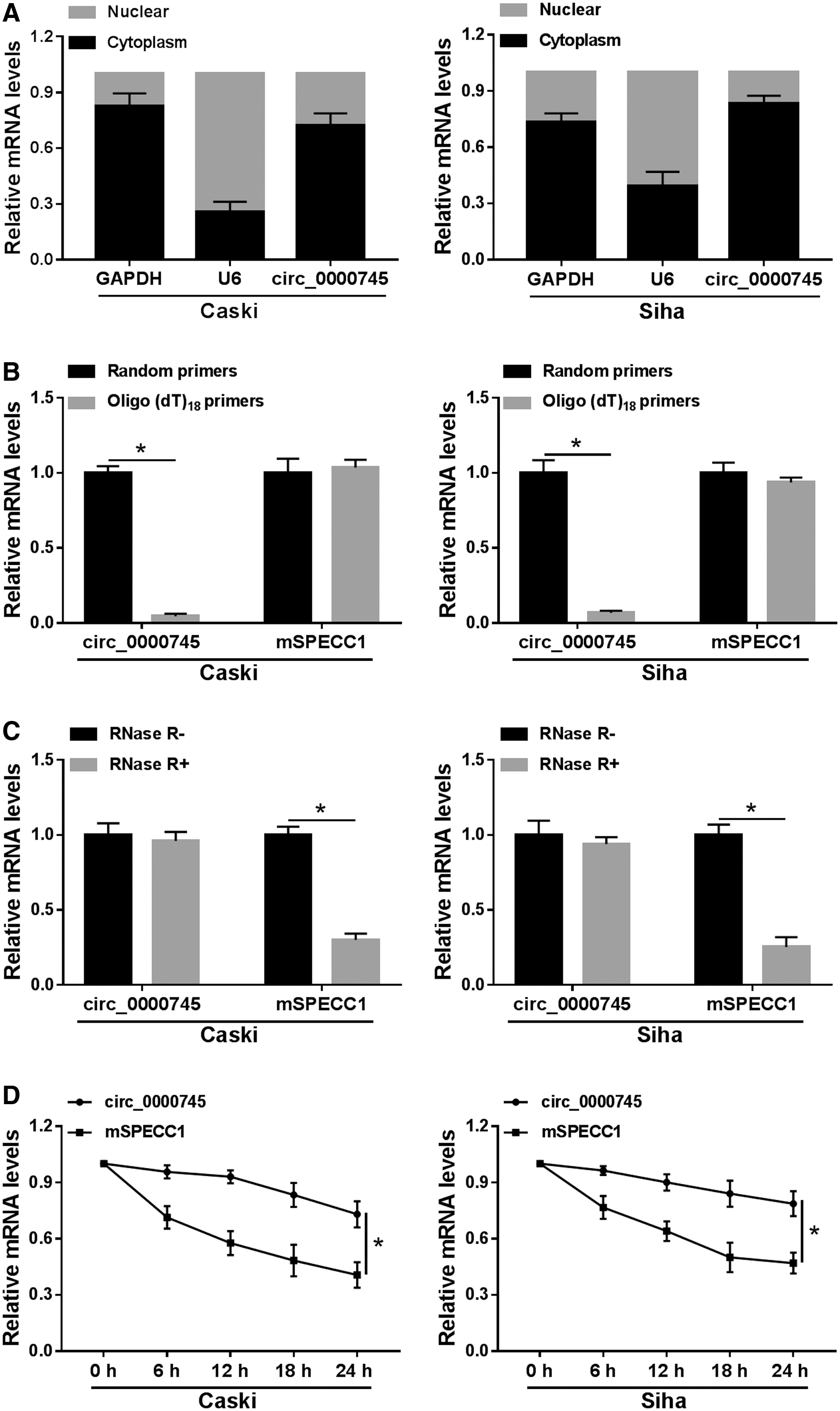
Identification of circ_0000745 in CC cells.
Knockdown of circ_0000745 impeded proliferation, migration, invasion, and glycolysis in CC cells
To investigate the effect of circ_0000745 on CC progression, they transfected Caski and Siha cells with si-circ_0000745. First, the knockdown efficiency was detected by qRT-PCR, and the results showed that the expression of circ_0000745 in the si-circ_0000745 group was drastically lower compared with the si-NC group (Fig. 3A). Next, colony formation assay suggested that inhibition of circ_0000745 strikingly reduced the colony number (Fig. 3B). Transwell assay exhibited that circ_0000745 silencing observably suppressed migration and invasion of Caski and Siha cells compared with the si-NC group (Fig. 3C, D). Moreover, to explore the role of circ_0000745 in glycolysis of CC cells, ECAR was first tested. The results showed that introduction of si-circ_0000745 predominantly decreased the ECAR relative to the control group (Fig. 3E). Also, knockdown of circ_0000745 reduced glucose uptake and lactate production (Fig. 3F, G). They also examined the expression of glycolysis-related proteins using Western blot, and the results revealed that the levels of GLUT1 and LDHA in Caski and Siha cells transfected with si-circ_0000745 were markedly downregulated compared with the si-NC group (Fig. 3H). These data evidenced that circ_0000745 depletion inhibited proliferation, migration, invasion, and glycolysis of CC cells.
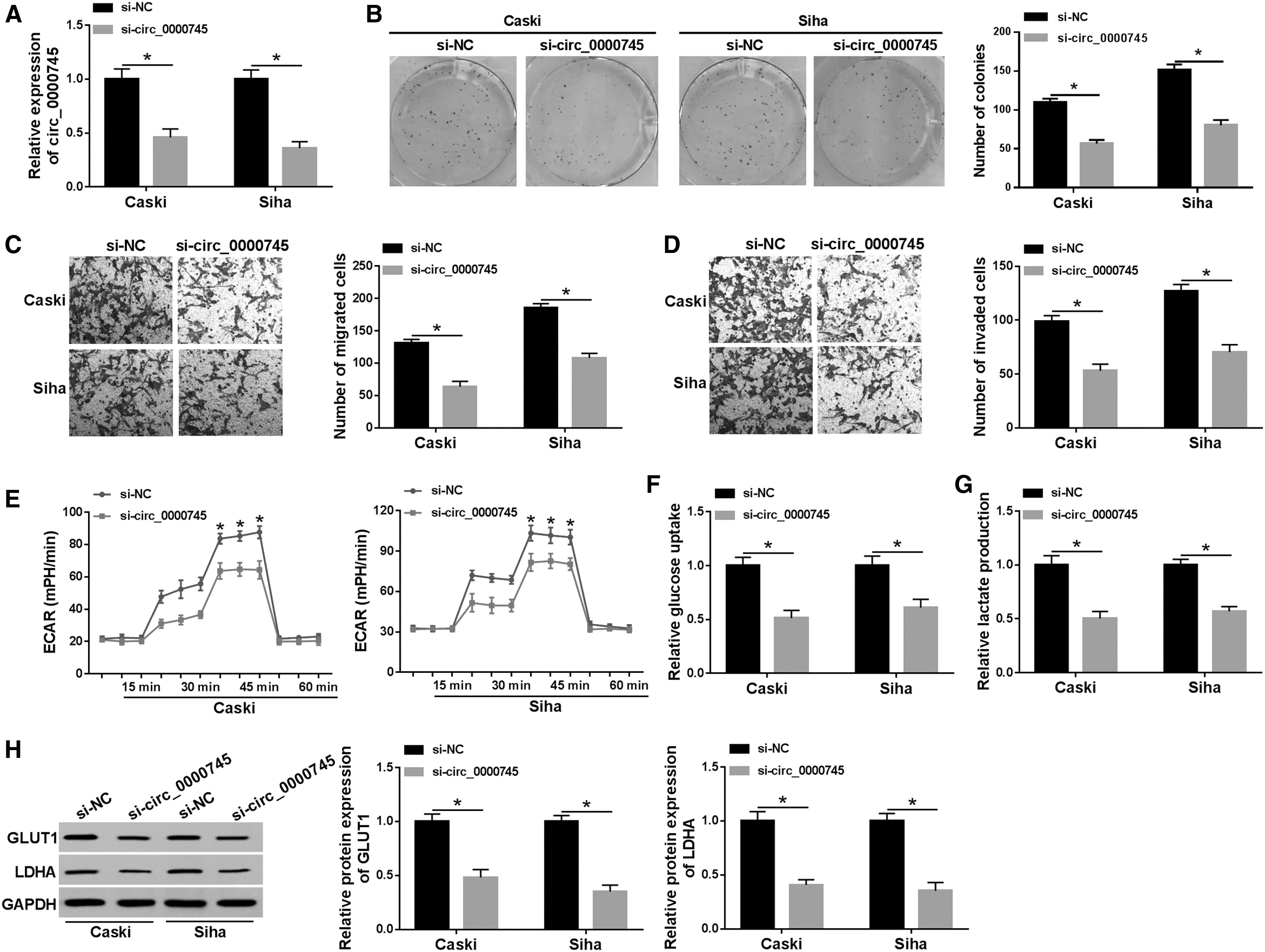
Knockdown of circ_0000745 impeded proliferation, migration, invasion, and glycolysis in CC cells. Caski and Siha cells were introduced with si-NC or si-circ_0000745.
Circ_0000745 was a sponge of miR-409-3p
To elucidate the mechanism of circ_0000745 in CC, they used circRNA Interactome online database to identify putative miRNAs that might bind to circ_0000745. They screened five miRNAs (miR-1246, miR-136, miR-409-3p, miR-145, and miR-296-5p) downregulated in CC. Then, qRT-PCR results showed that the expression difference of miR-409-3p was the most significant among the five differentially expressed miRNAs, so miR-409-3p was selected for subsequent research (Supplementary Fig. S1A). As shown in Figure 4A, miR-409-3p has binding sites with circ_0000745. To confirm whether circ_0000745 targeted miR-409-3p, a dual-luciferase reporter assay was performed. The results exhibited that miR-409-3p mimics strikingly reduced the luciferase activity of WT-circ_0000745 reporter (Fig. 4B). Besides, miR-409-3p expression was dramatically decreased in CC tissues and cells compared with adjacent tissues and ECT1/E6E7 cells (Fig. 4C, D). Also, the expression of circ_0000745 was overtly elevated in Caski and Siha cells transfected with circ_0000745, indicating a productive transfection efficiency (Fig. 4E). Next, circ_0000745 overexpression evidently reduced miR-409-3p level, and circ_0000745 knockdown significantly raised the miR-409-3p level (Fig. 4F). Collectively, these data unveiled that circ_0000745 directly targeted miR-409-3p in CC.
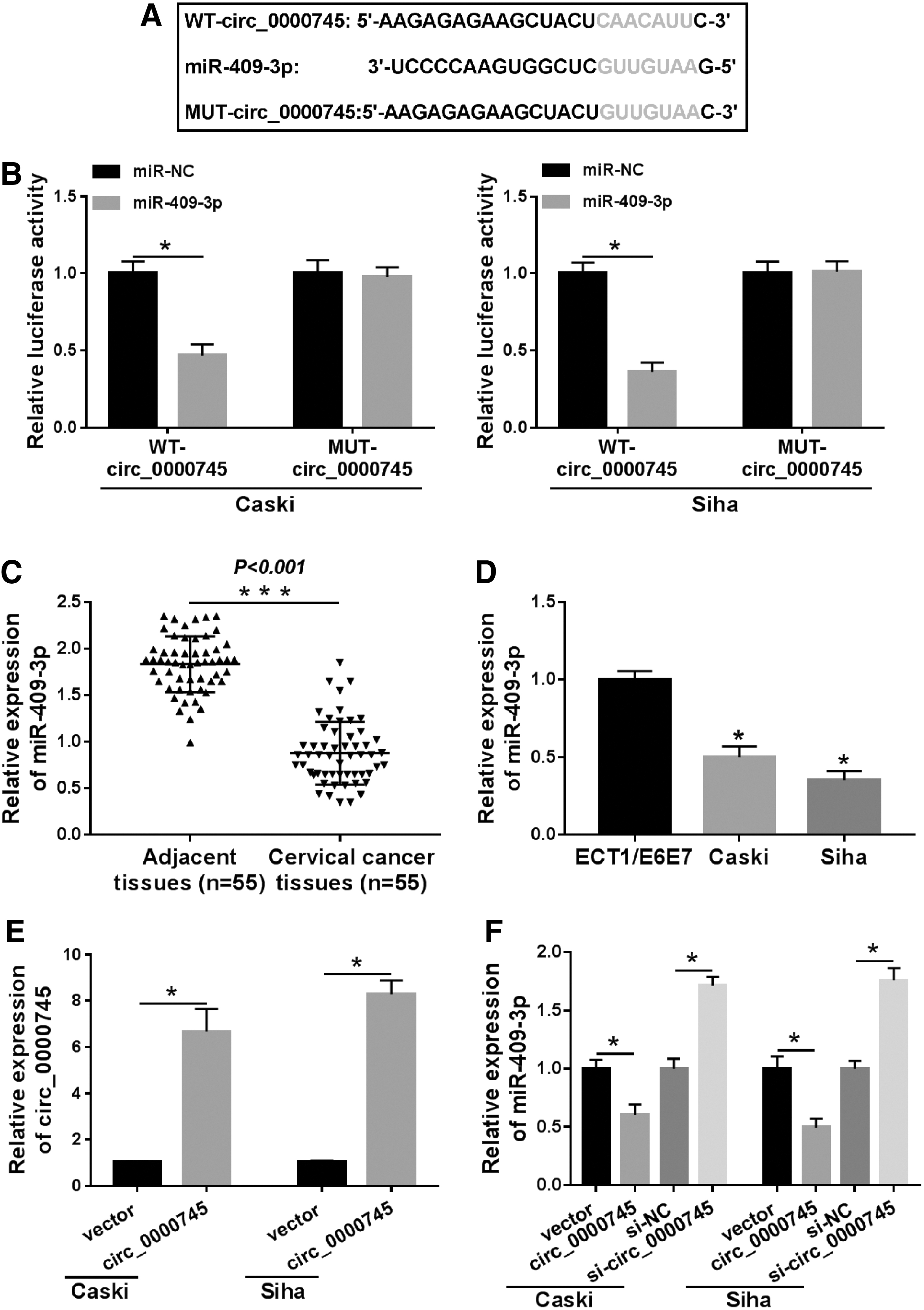
Circ _0000745 was a sponge of miR-409-3p.
Circ_0000745 facilitated CC progression by targeting miR-409-3p
To explore whether circ_0000745 could act as a sponge molecule of miR-409-3p to regulate CC progression, Caski and Siha cells were transfected with vector+miR-NC, vector+miR-409-3p, miR-NC+circ_0000745, or circ_0000745+miR-409-3p for rescue experiments. First, transfection of miR-409-3p substantially increased the expression of miR-409-3p, and overexpression of miR-409-3p reversed the inhibition of circ_0000745 upregulation on miR-409-3p level (Fig. 5A). In addition, upregulation of miR-409-3p remarkably reduced the number of colonies, migrated cells, and invaded cells, while cotransfection of miR-409-3p and circ_0000745 alleviated the promotion of circ_0000745 overexpression on cell proliferation, migration, and invasion (Fig. 5B–D). Besides, miR-409-3p mimics drastically diminished ECAR, glucose uptake, and lactate production compared with the control group, whereas the glycolytic process induced by circ_0000745 upregulation was reversed after transfection with miR-409-3p (Fig. 5E–G). Simultaneously, miR-409-3p overexpression resulted in a marked reduction of GLUT1 and LDHA levels, while increased GLUT1 and LDHA proteins induced by circ_0000745 overexpression were reverted by upregulating miR-409-3p (Fig. 5H). These data certified that circ_0000745 expedited CC progression by regulating miR-409-3p in CC cells.
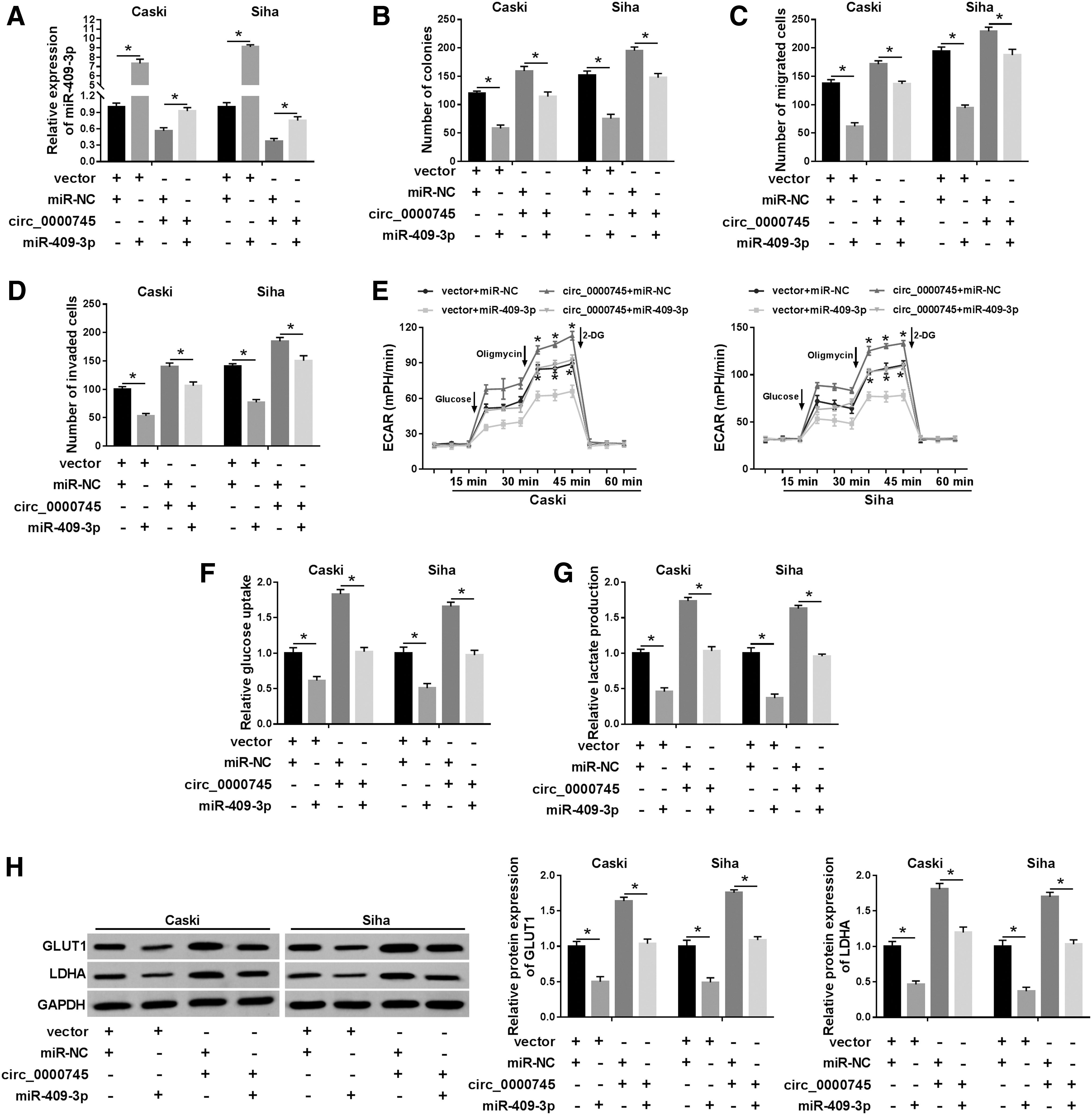
Circ _0000745 facilitated CC progression by targeting miR-409-3p. Caski and Siha cells were introduced with vector+miR-NC, vector+miR-409-3p, miR-NC+circ_0000745, or circ_0000745+miR-409-3p.
ATF1 was a target of miR-409-3p
The starBase database predicted that miR-409-3p and ATF1-3′UTR had putative binding sites (Fig. 6A). Besides, they selected four other target genes (HSPA8, RRM2, FBXL5, and ANXA2) that might bind to miR-409-3p. Next, qRT-PCR results revealed that ATF1 was most significantly upregulated among the five differentially expressed target genes, so ATF1 was selected for further research (Supplementary Fig. S1B). Dual-luciferase reporter assay was utilized to verify the relationship between miR-409-3p and ATF1, and the results displayed that mature miR-409-3p evidently decreased the luciferase activity of WT-ATF1-3′UTR reporter (Fig. 6B). Additionally, the mRNA and protein levels of ATF1 in CC tissues were notably higher than those in adjacent tissues (Fig. 6C, D). Compared with ECT1/E6E7 cells, the mRNA and protein levels of ATF1 in Caski and Siha cells were dramatically elevated (Fig. 6E, F). Also, miR-409-3p had an outstanding knockdown efficiency in Caski and Siha cells transfected with anti-miR-409-3p (Fig. 6G). Moreover, overexpression of miR-409-3p repressed the protein level of ATF1, while the effect was rescued after introduction with circ_0000745 (Fig. 6H). These data indicated that ATF1 was a target of miR-409-3p in CC cells.
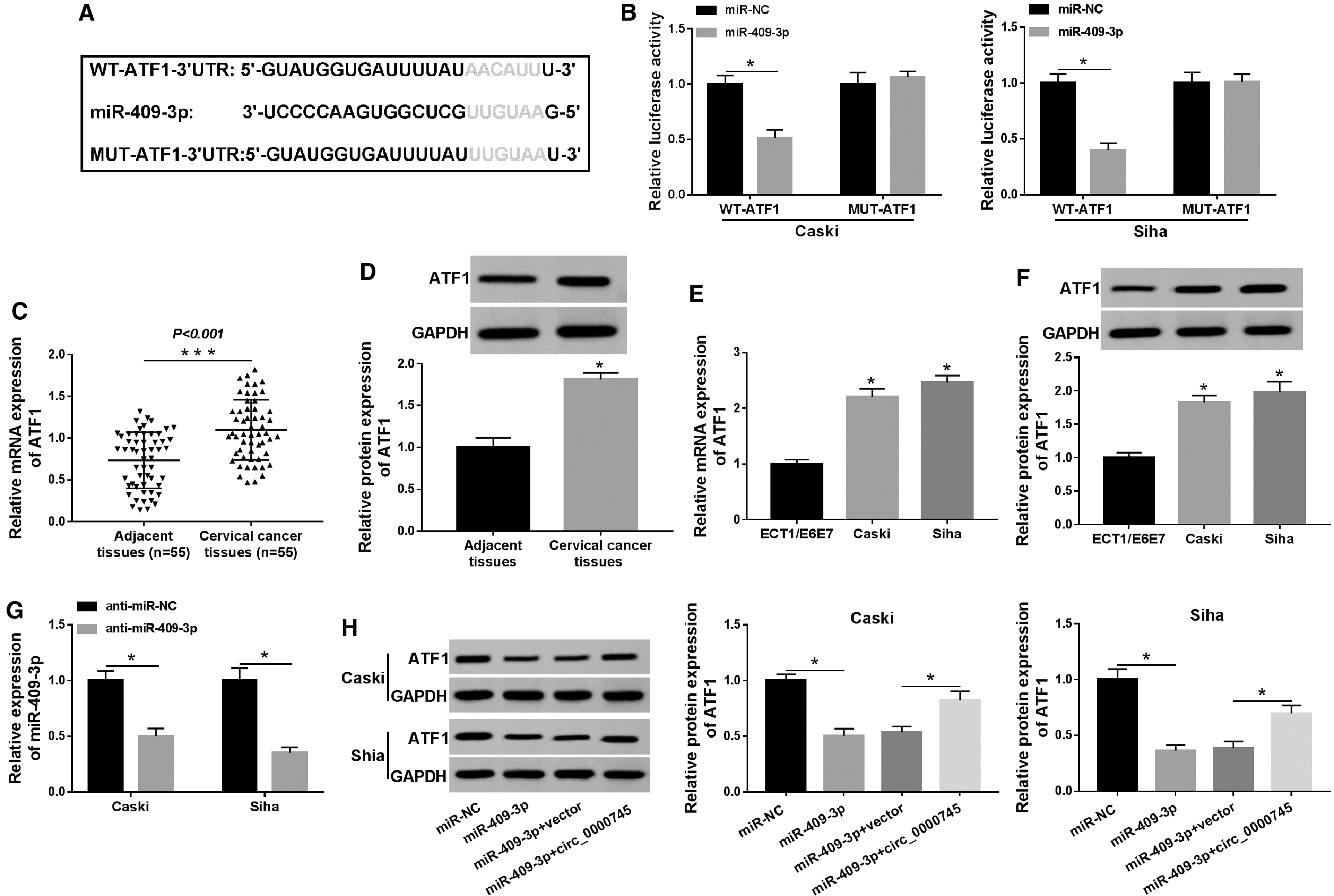
ATF1 was a target of miR-409-3p.
Inhibition of miR-409-3p expedited CC progression by regulating ATF1
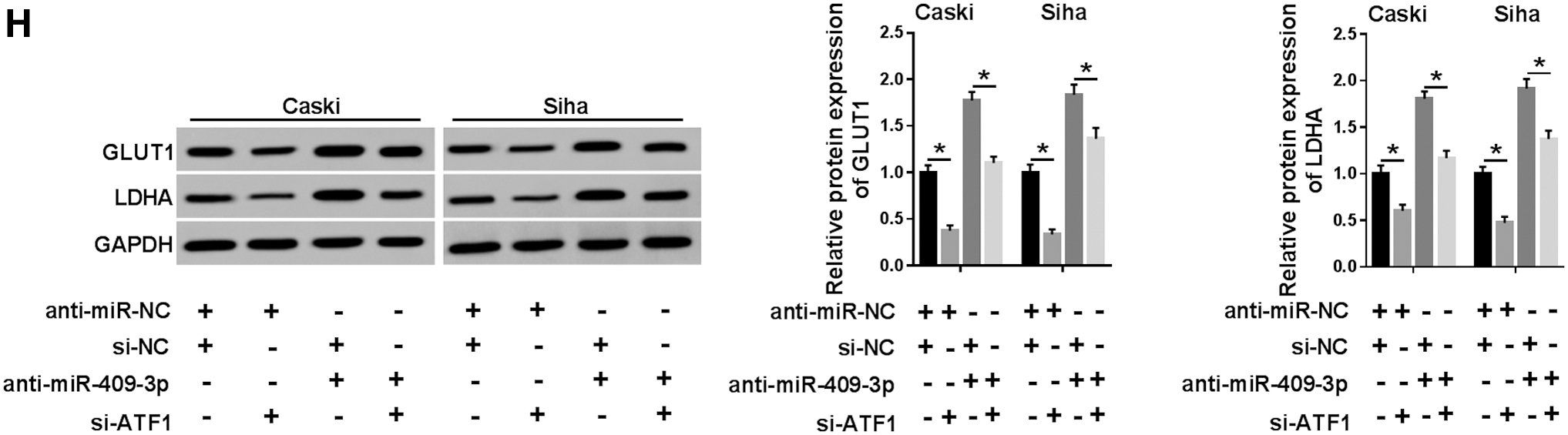
We further performed rescue experiments to clarify the role of miR-409-3p targeting ATF1 in CC development. First of all, knockdown of ATF1 remarkably decreased the protein level of ATF1, and cotransfection of si-ATF1 and anti-miR-409-3p reversed the promotion of miR-409-3p inhibition on ATF1 expression (Fig. 7A). Besides, ATF1 silencing suppressed proliferation, migration, and invasion of Caski and Siha cells, whereas the promoting effect caused by miR-409-3p knockdown was abolished by reintroduction of si-ATF1 (Fig. 7B–D). In addition, depletion of ATF1 evidently declined ECAR (Fig. 7E), glucose uptake (Fig. 7F), lactate production (Fig. 7G), and the expression of GLUT1 and LDHA (Fig. 7H) relative to the control group, whereas cotransfection of si-ATF1 and anti-miR-409-3p undermined the promotion of glycolysis induced by downregulating miR-409-3p (Fig. 7E–H). These data concluded that suppression of miR-409-3p accelerated CC progression by modulating ATF1.
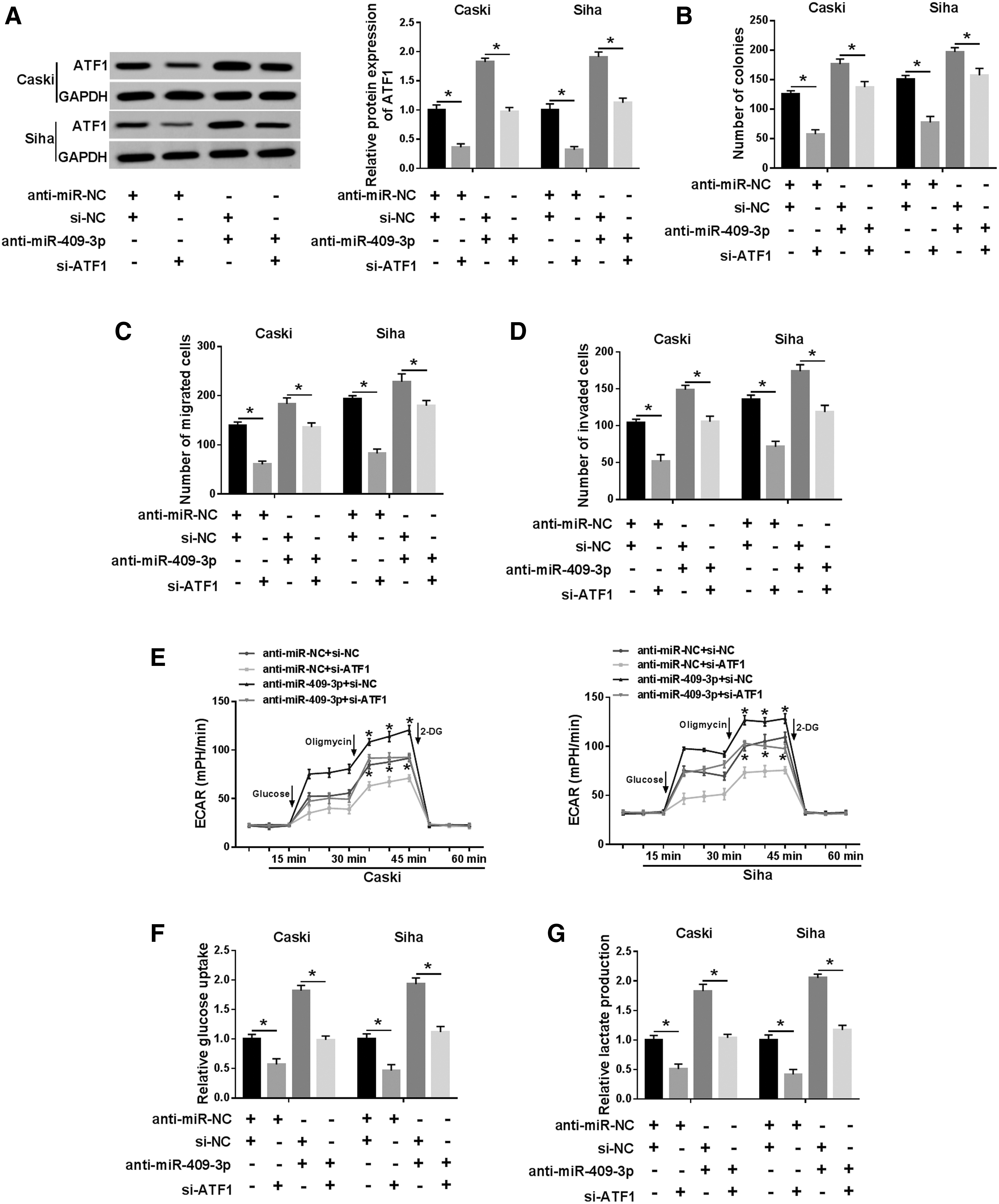
Inhibition of miR-409-3p expedited CC progression by regulating ATF1. Caski and Siha cells were transduced with anti-miR-NC+si-NC, anti-miR-NC+si-ATF1, si-NC+anti-miR-409-3p, or anti-miR-409-3p+si-ATF1.
Knockdown of circ_0000745 blocked tumor growth in vivo
Next, a xenograft mice model was established to analyze the role of circ_0000745 in tumorigenesis in vivo. The results suggested that tumor volume and weight in the sh-circ_0000745 group were observably reduced in comparison with the sh-NC group (Fig. 8A, B). Besides, the expression of circ_0000745, miR-409-3p, and ATF1 was measured using qRT-PCR or Western blot. The results showed that knockdown of circ_0000745 resulted in a striking decrease in circ_0000745 expression and an evident increase in miR-409-3p expression (Fig. 8C, D). Western blot analysis revealed that the protein level of ATF1 in the sh-circ_0000745 group was markedly lower compared with the sh-NC group (Fig. 8E). These data indicated that circ_0000745 depletion impeded tumor growth in vivo.
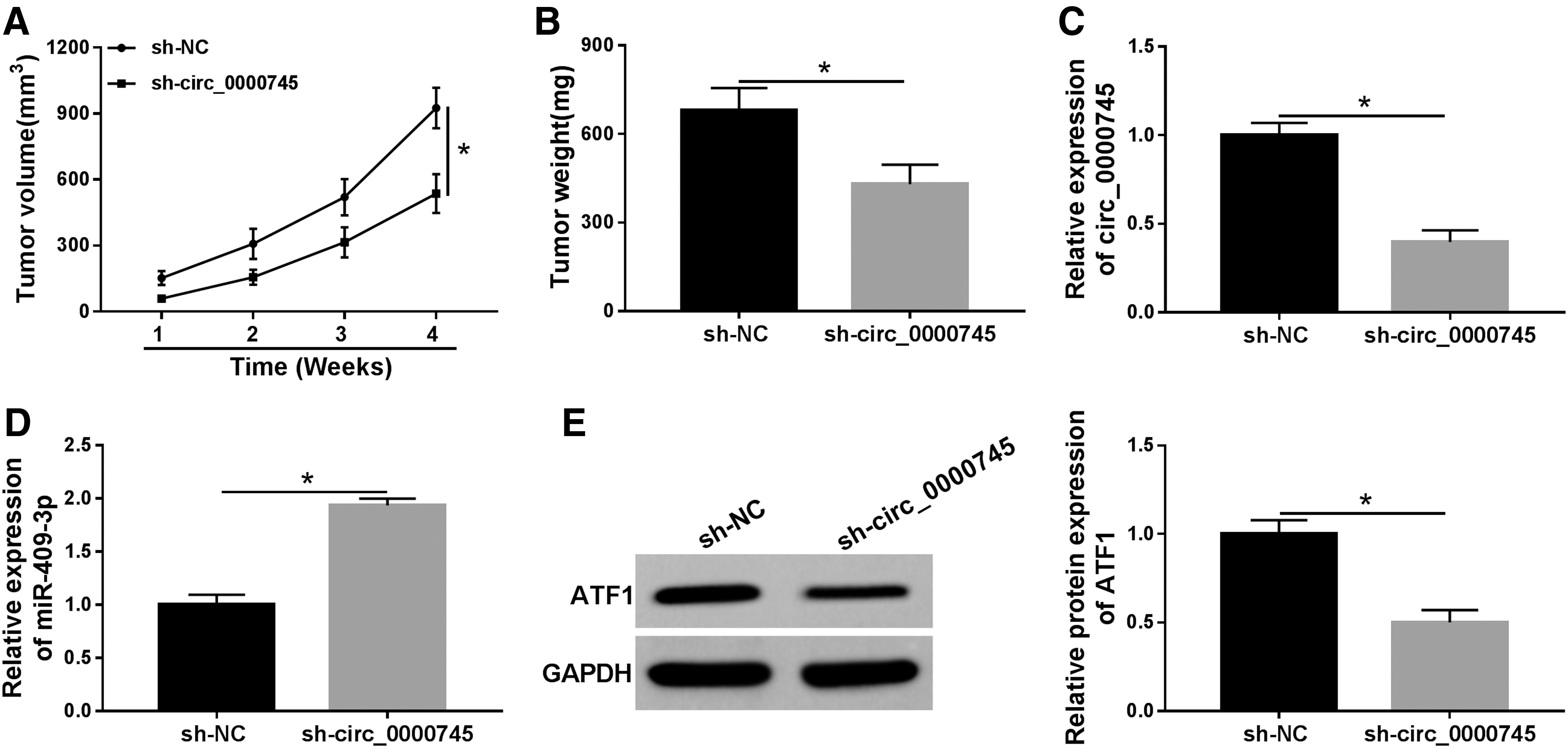
Knockdown of circ_0000745 blocked tumor growth in vivo. Siha cells introduced with sh-NC or sh-circ_0000745 were subcutaneously injected into nude mice.
Discussion
Warburg effect is marked by increased glycolysis of cancer cells, providing a driving force for cancer proliferation and metastasis. 21 GLUT1 is an indicator of anaerobic glycolysis and a speed-limiting transporter for glucose uptake. 22 LDHA accelerates glycolysis by converting pyruvate to lactate. 23 In addition, ncRNAs play a crucial role in glucose metabolism reprogramming. 24
Emerging evidence has demonstrated that circRNAs regulate a range of biological processes by acting as a miRNA molecular sponge. 25 For example, circDENND4C sponged miR-200b/c to modulate glycolysis and metastasis of breast cancer cells under hypoxia. 26 CircRNA_100290 functioned as a ceRNA to facilitate glycolysis and growth in oral squamous cell carcinoma by regulating miR-378a/GLUT1 axis. 27 Besides, some circRNAs have been evidenced to be strongly associated with CC progression. 28 For instance, Rong et al. suggested that circ_0007534 silencing suppressed the development of CC by upregulating miR-498 and repressing BMI-1. 29 Mao et al. discovered that circEIF4G2 aggravated the malignancy of CC by sponging miR-218 and activating HOXA1 expression. 30 Song et al. revealed that circRNA_101996 regulated miR-8075 and TPX2 through the ceRNA mechanism, thus promoting the progression of CC. 31 In the present study, circ_0000745 was conspicuously upregulated in CC tissues and cells, and circ_0000745 knockdown confined CC progression, which is consistent with previous research. 12
The prediction software predicted the possible targets for circ_0000745. The authors chose miR-409-3p for the following study because their relationship has not been investigated, and miR-409-3p plays a pivotal regulatory role in tumors. For example, miRNA-409-3p targeted ZEB1 to curb the development of osteosarcoma. 32 miRNA-409-3p impeded glioma progression through inactivating HMGN5. 33 miRNA-409-3p repressed cell proliferation in breast cancer through regulation of Akt1. 34 In addition, small RNA sequencing results indicated that miR-409-3p expression was overtly reduced in CC. 35 Nevertheless, the specific role of miR-409-3p in CC has not been clarified. Hence, they validated that miRNA-409-3p was a direct target of circ_0000745. What is more, miRNA-409-3p reversed the promotion of CC progression caused by circ_0000745 overexpression.
Accumulating evidence has manifested that miRNAs suppressed protein synthesis by base pairing to mRNA 3′UTR. 13 They also verified that ATF1 was a target of miR-409-3p. ATF1 is classified into the cAMP response element-binding protein family that influences biological processes in tumors by modulating tumor-associated cellular signaling pathways. 36 In nasopharyngeal carcinoma, Pin1 strengthened the transcription activity and tumor-promoting effect of ATF1. 37 In hepatocellular carcinoma, ATF1 abolished the inhibitory effect of lncRNA GHET1 silencing on tumor progression. 38 Moreover, ATF1 and RAS (small GTPase) genes were dramatically upregulated in tumors and blood exosomes in both primary and recurrent CC mouse models. 39 According to previous reports, ATF1 regulated HKII transcription by binding to the promoter region of HKII. 40 In this study, they uncovered that miR-409-3p bound to ATF1 and miR-409-3p inhibition contributed to CC progression by targeting ATF1.
In conclusion, these findings concluded that circ_0000745 acted as a ceRNA of miR-409-3p to activate ATF1. More importantly, circ_0000745 facilitated proliferation, metastasis, and glycolysis of CC cells by regulating the miR-409-3p/ATF1 axis. Also, circ_0000745 depletion blocked tumor growth in vivo. Therefore, this study provided a basis for circ_0000745 as a potential therapeutic target for CC.
Footnotes
Authors' Contributions
Conception and design: X.C.; Development of methodology: J.C. and J.C.; Acquisition, analysis and interpretation of data: Y.Z.; Writing, review, and revision of article: X.C. and H.S.; All coauthors have reviewed and approved of the article before submission.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.