
Abstract
MicroRNA-24 (miR-24) has been identified to be related to the development of glioma. However, the exact molecular mechanism of miR-24 in glioma progression remains vague. The aim of the present study was to investigate the role of miR-24 in sepsis and to reveal the associated mechanisms. Quantitative real-time polymerase chain reaction was used to compare the levels of miR-24 in glioma and normal tissue. The miR-24 inhibitor or miR-24 mimic was transfected into glioma cells, and then the effects of miR-24 on cell proliferation and apoptosis were detected using CCK-8 (Cell Counting Kit-8) assay and flow cytometry, respectively. Western blot was used to examine the levels of CDX1 (caudal-type homeobox 1), PI3K, p-PI3K, Akt, p-Akt, Cyclin D1, p27, proliferating cell nuclear antigen, Bcl-2, Bax, and Cleaved-casp3. Luciferase assay was used to identify the target gene of miR-24. An animal model was established in mice to detect the role of miR-24 in vivo. These results suggested that miR-24 was elevated in glioma, and miR-24 could promote glioma progression by facilitating cell proliferation and inducing cell apoptosis through CDX1/PI3K/Akt signaling pathway, indicating a novel pathway underlying progression in glioma cells and providing a potential target for glioma treatment.
Introduction
Glioma is the most common primary brain tumor in adults and accounts for over 70% of malignant brain tumors, with an annual incidence rate of 3.2 per 100,000 population. 1 Despite diversity molecular studies that aimed to develop new treatment strategies for glioma have been investigated, the 5-year overall survival rates of glioma are still pessimistic. 2,3 Thus, it is critical to better understand the underlying molecular mechanisms of glioma carcinogenesis and progression, and find promising diagnostic markers and therapeutic targets that are involved in glioma.
MicroRNA (miRNAs) are a class of small noncoding RNAs molecule, with ∼22 nucleotides in length, which are implicated in gene-regulation at the posttranscriptional level through partially or completely complementary sequence pairing to the 3′-untranslated regions (3′-UTR) of mRNA target, thus inducing translational repression and mRNA degradation. 4,5 MiRNAs have been identified to participate in numerous cellular biological and pathological processes, including growth, proliferation, apoptosis, angiogenesis, and carcinogenesis. 6,7 Until now, a considerable body of evidence has indicated that miRNAs are closely associated with the prognosis and development of glioma. 8 –10 MicroRNA-24 (miR-24) is a member of the miR-23b cluster, and has been reported to act as an oncogene to involve in the development of various cancers, such as breast cancer, gastric cancer, and nonsmall cell lung cancer. 11 –13 In glioma, miR-24 was demonstrated to be upregulated 14 and Chen et al. had reported that miR-24 regulated cell proliferation and invasion in glioma by ST7L via β-catenin/Tcf-4 signaling. 15 However, details regarding the regulation mechanisms of miR-24 in glioma remain obscure. The caudal-type homeobox 1 (CDX1) transcription factor, an intestine-specific transcription factor, belongs to the caudal-related homeobox transcription factor gene family. 16,17 In recent, increasing evidence has identified that abnormally expressed CDX1 is associated with diverse cancers development, and maybe a possible therapeutic target for anticancer therapy. 18 –20 Yang et al. found that CDX1 was a target of miR-155 and involved in miR-155-induced proliferation of glioma cells. Thus, CDX1 may function as an important regulator in the development of glioma.
In the present study, the authors investigated the expression patterns of miR-24 in glioma tissue and cell lines, and explored molecular mechanisms underlying the function of miR-24 on glioma progression and prognosis. In addition, they also explored the potential interaction between miR-24 and CDX1 in glioma, which might contribute to provide novel therapeutic targets and biomarkers for glioma.
Materials and Methods
Tissue samples and clinical data
A total of 40 primary glioma tissues and normal brain tissue samples were collected from Hanyang Hospital Affiliated to Wuhan University of Science and Technology, from those who received surgical treatment. All specimens were confirmed by two independent pathologists and did not receive any preoperative treatment. Specimens were immediately stored at −80°C for further study. The clinical features, including age, gender, WHO grade, and tumor size, were obtained from the recruited cases. Follow-up was conducted regularly every 3 months in the first 2 years after surgery, and from the third year reduced to once every 6 months. The last follow-up was carried out in July 2017. The study was permitted by the Ethics Committee of Hanyang Hospital Affiliated to Wuhan University of Science and Technology and all patients had signed the written informed consents.
Cell culture and transfection
Glioma cell line A172, U251, and SHG-44 were obtained from Shanghai Cell Bank of the Chinese Academy of Science (Shanghai, China), human glioma cell line HS683, and normal human astrocyte cell were purchased from the American Type Culture Collection (ATCC, Manassas, VA). All cells were cultured in Dulbecco's modified Eagle medium (DMEM; Gibco, Carlsbad, MD) containing with 10% fetal bovine serum (FBS; Gibco) and 100 μg/mL streptomycin/penicillin at 37° with 5% CO2.
Small interfering RNA (siRNA) targeting CDX1 (si-CDX1) (si-CDX1 sequence, 5′-GATCCGACTCGGACCAAGGACAAGTTCAAGAGACTTGTCCTTGGTCCGAGTCTTTTTTGGAAA-3′) and siRNA negative control (si-NC), empty vector (vector), and pcDNA3.1-CDX1 wild-type or mutant overexpression vector (CDX1 WT/MUT) were synthesized by Genepharma (Shanghai, China). The miR-24 mimic (miR-24), mimic negative control (miR-NC), miR-24 inhibitor, and inhibitor negative control (inhibitor-NC) were purchased from RiboBio (Guangzhou, China). All oligonucleotides or vectors were transfected into A172 and U251 cells using Lipofectamine™ 2000 reagent (Invitrogen, Carlsbad, CA), respectively. Then cells were harvested at 48 h for the subsequent analysis.
Quantitative real-time polymerase chain reaction
Total RNA was isolated from the tissues or cell lines with TRIzol reagent (Invitrogen). The cDNA was generated using M-MLV reverse transcriptase (Invitrogen). Then real-time polymerase chain reaction (PCR) was conducted using SYBR Green PCR Master Mix (Applied Biosystems, Foster City, CA) according to the manufacturer's introduction. GAPDH or U6 was as an endogenous control and the 2−ΔΔCt method was used to calculate the fold changes. The special primers for miR-24 or U6 were purchased from Qiagen and primers for CDX1 or GAPDH were listed as follows: CDX1 5′-GGCAGAAGATGAGCAGGATT-3′ (forward) and 5′-TCCAAAGACTGAATGCGAGA-3′ (reverse); GAPDH 5′-AACGGATTTGGTCGTATTGG-3′ (forward) and 5′-TTGATTTTGGAGGGATCTCG-3′ (reverse).
Cell proliferation assay
Cell proliferation capacity was quantitated by a Cell Counting Kit-8 (CCK-8) (Dojindo, Tokyo, Japan) following the standard protocol. In brief, transfected cells (5 × 103 cells/well) were seeded into 96-well plates with DMEM (with 10% FBS) and cultured for 1, 2, 3, 4, and 5 d, respectively, and then each well was added with 10 μL of CCK-8 solution at point time and incubated for another 1 h. The absorbance value was measured at 450 nm using a microplate reader (Bio-Rad, Hercules, CA).
Cell apoptosis analysis
Annexin V-FITC/PI apoptosis detection kit (Solarbio, Beijing, China) was used to analyze cell apoptosis. After transfected, cells were resuspended with binding buffer and then stained with the FITC-Annexin V and propidium iodide (PI) in the dark. The results were detected by flow cytometry (BD Biosciences, San Jose, CA), and the data were analyzed by the ModFit LT.
Luciferase reporter assay
The CDX1 3′-UTR containing wild-type or mutant binding sequence of miR-24 was cloned into the pGL3 basic vectors (Promega, Shanghai, China), respectively. Then, A172 and U251 cells were seeded in 24-well plates, following cotransfection with CDX1-WT or CDX1-MUT with miR-24 mimics or miR-NC using Lipofectamine 2000 (Invitrogen). After 48 h transfection, a dual luciferase assay kit (Promega) was utilized to analyze the luciferase activity following the manufacturer's instructions.
Western blot
Western blot assays were performed as described before. 21 Immunoblot assays were carried out using antibodies against CDX1 (1:3000, ab126748; Abcam, Cambridge, MA), phosphorylated (p-) PI3K (1:1000, ab182651; Abcam), PI3K (1:1000, ab40776; Abcam), p-AKT (1:1000, 9271; Cell Signaling Technology, Boston, MA), AKT (1:1000, 9272; Cell Signaling Technology), proliferating cell nuclear antigen (PCNA) (1:1000, ab18197; Abcam), Bcl-2 (1:500, ab692; Abcam), Bax (1:1000, ab32503; Abcam), cleaved caspase3 (Cleaved cas3) (1:1000, ab32042; Abcam), and GAPDH (1:10,000, ab181602; Abcam).
Xenograft experiments in vivo
BALB/c nude mice (4–6 weeks old, N = 6) were used to perform Xenograft experiments according to the guidelines permitted by the Animal Research Committee of Hanyang Hospital Affiliated to Wuhan University of Science and Technology and the Ethics Committee of the Animal Experimental Center of Hanyang Hospital Affiliated to Wuhan University of Science and Technology approved this study. Approximately 5 × 106 A172 cells stably injected with lentivirus-inhibitor NC or lentivirus-miR-24 inhibitor were subcutaneously inoculated into the flanks of the nude mice. The tumor size in mice was detected every 7 d. At day 28, all mice were sacrificed by inhaling with 2% methoxyflurane and the death of the mice was determined by continuous involuntary breathing (2–3 min) and no blink reflex. Then the tumor masses were weighted and harvested for further molecular analysis.
Statistical analyses
All statistical analyses are presented as the mean ± standard deviation and analyzed using GraphPad Prism 7 (GraphPad, Inc., San Diego, CA). The significant group differences were evaluated using Student's t-test or one-way analysis of variance (ANOVA) followed by Tukey's post hoc test. The correlation between miR-24 and CDX1 was analyzed by Pearson's correlation analysis. The correlation between miR-24 and clinicopathological features of the patients was analyzed using a χ-square test. Kaplan–Meier survival curves were plotted and the difference in survival between two groups was analyzed by the log-rank test. The p-value <0.05 was considered as statistically significant.
Results
MiR-24 is elevated in glioma tissues and cell lines and highly expressed miR-24 predicts poor prognosis
The expression of miR-24 was measured and it was found that miR-24 was significantly increased in glioma tissues as well as in glioma cell lines, including A172, U251, SHG-44, and HS683, compared with that of the controls (Fig. 1A, C). Subsequently, patients were grouped as follows: a high miR-24 group possessing miR-24 expression higher than the median level (8.92) and a low miR-24 group possessing miR-24 expression lower than the median level, and results showed that compared with low miR-24 group, highly expressed miR-24 predicted worse prognosis in glioma patients (Fig. 1B). Furthermore, the correlation between the miR-24 and clinicopathological features in glioma patients was analyzed (Table 1). The table suggested that miR-24 expression was closely associated with glioma grade and tumor size, while not related to age and gender; besides that, miR-24 expression also was related to the malignancy of glioma. Therefore, it was verified that miR-24 was upregulated in glioma tissues and cell lines and highly expressed miR-24 predicted poor prognosis.
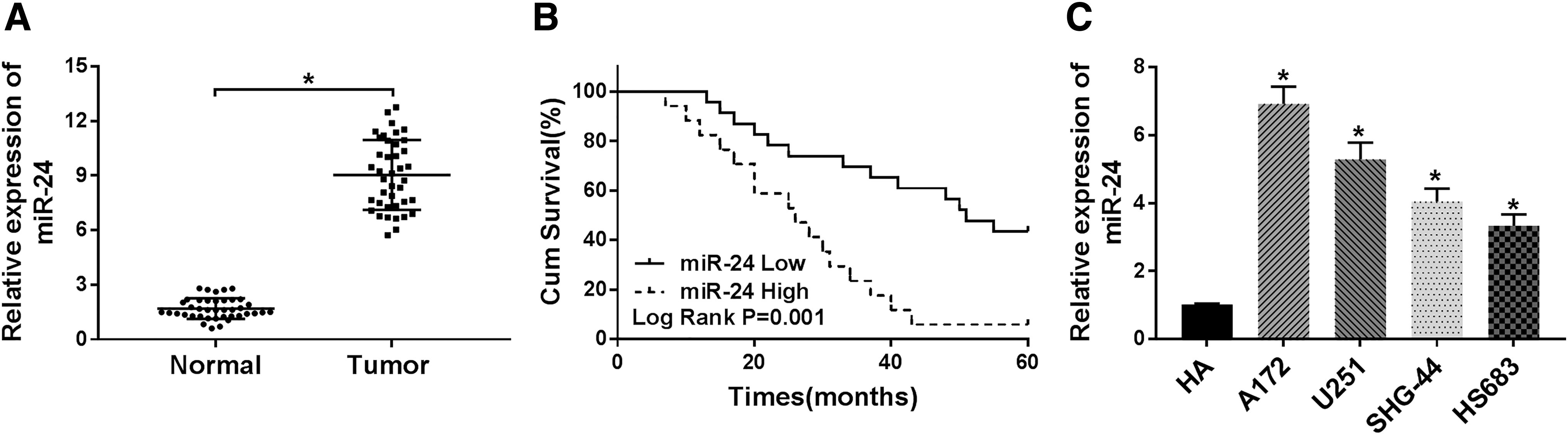
MiR-24 is elevated in glioma tissues and cell lines and highly expressed miR-24 predicts poor prognosis.
Relationship Between miR-24 Expression and Clinicopathological Features of Glioma (n = 40)
p < 0.05.
miR-24, microRNA-24.
MiR-24 inhibitor antagonizes proliferation and induces apoptosis in glioma cells
To explore the effects of miR-24 on the glioma progression, A172 and U251 cells were transfected with miR-24 inhibitor or inhibitor NC. After transfection, a decrease of miR-24 expression was measured in A172 and U251 cells, showing successful transfection (Fig. 2A). Then, cell proliferation was quantitated and the data indicated that miR-24 inhibitor suppressed proliferation of A172 and U251 cells (Fig. 2B, C). In the meanwhile, cell apoptosis rate was detected and it was found that miR-24 inhibition promoted cell apoptosis in glioma, especially inhibiting late apoptosis (Fig. 2D). Furthermore, the expression of PCNA, Bcl-2, Bax, and Cleaved-cas3 protein was detected, and Western blot analysis exhibited that miR-24 inhibition significantly decreased the level of PCNA and Bcl-2, but increased the level of Bax and Cleaved-cas3 in A172 and U251 cells (Fig. 2E, F), further suggesting the regulatory roles of miR-24 in glioma cell proliferation and apoptosis. Thus, these results suggested that miR-24 inhibition repressed glioma development by regulating cell proliferation and apoptosis.
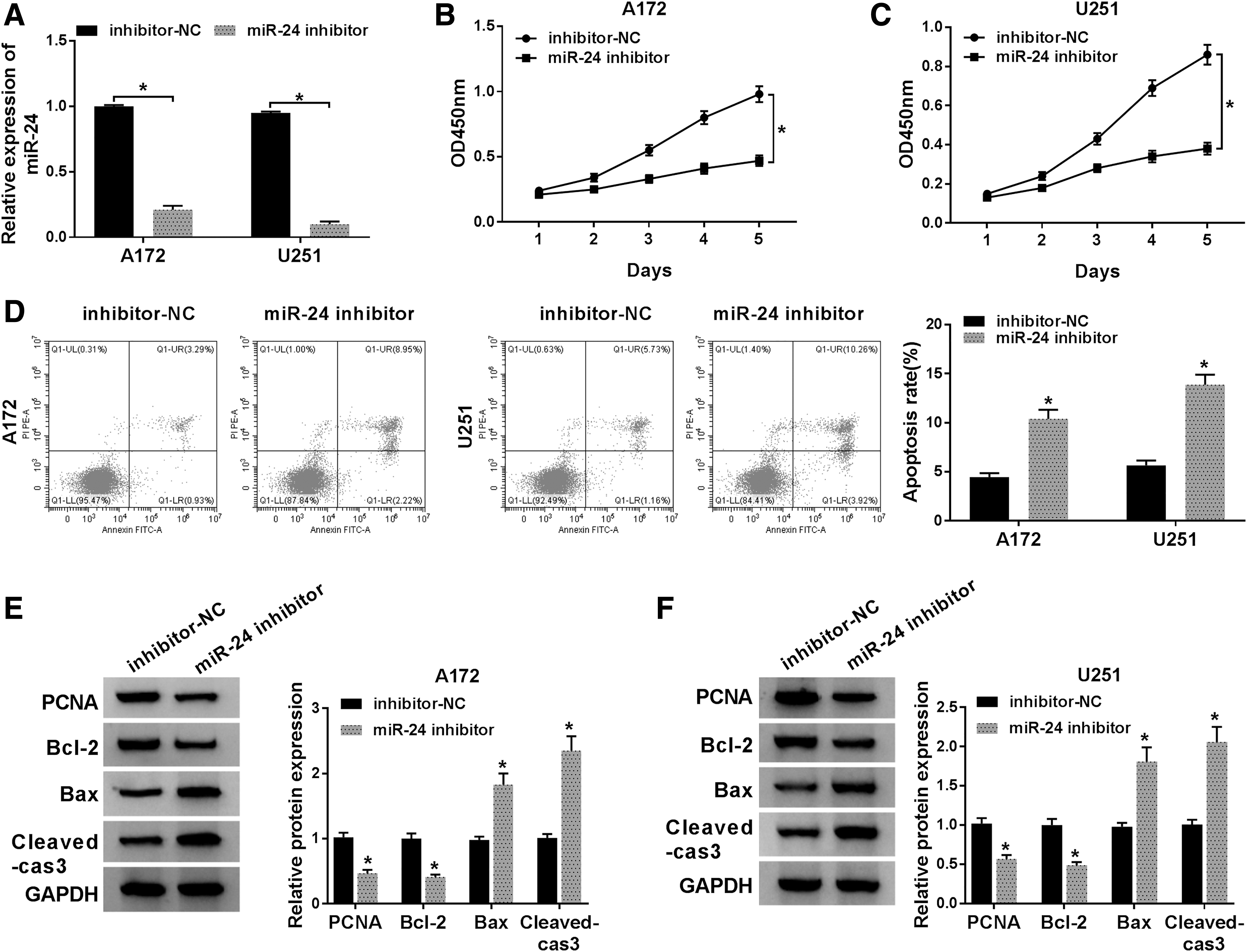
MiR-24 inhibitor antagonizes proliferation and induces apoptosis in glioma cells. A172 and U251 cells were transfected with miR-24 inhibitor or inhibitor-NC.
MiR-24 overexpression promotes proliferation and suppresses apoptosis in glioma cells
The authors further investigated the influence of overexpressed miR-24 on glioma cell proliferation and apoptosis. A172 and U251 cells were transfected with miRNA mimics or miR-24 mimic. As expected, miR-24 was significantly elevated in A172 and U251 cells transfected with miR-24 mimic (Fig. 3A). After that, it was found miR-24 overexpression promoted proliferation of A172 and U251 cells, reflected by the increased proliferating cell and PCNA expression (Fig. 3B, C, E, F). Meanwhile, miR-24 overexpression also inhibited apoptosis of A172 and U251 cells, as illustrated by the decreased apoptotic cells and Bax, Cleaved-cas3 expression, as well as the increase of Bcl-2 expression (Fig. 3D–F). Therefore, the authors demonstrated that miR-24 overexpression promoted glioma development.
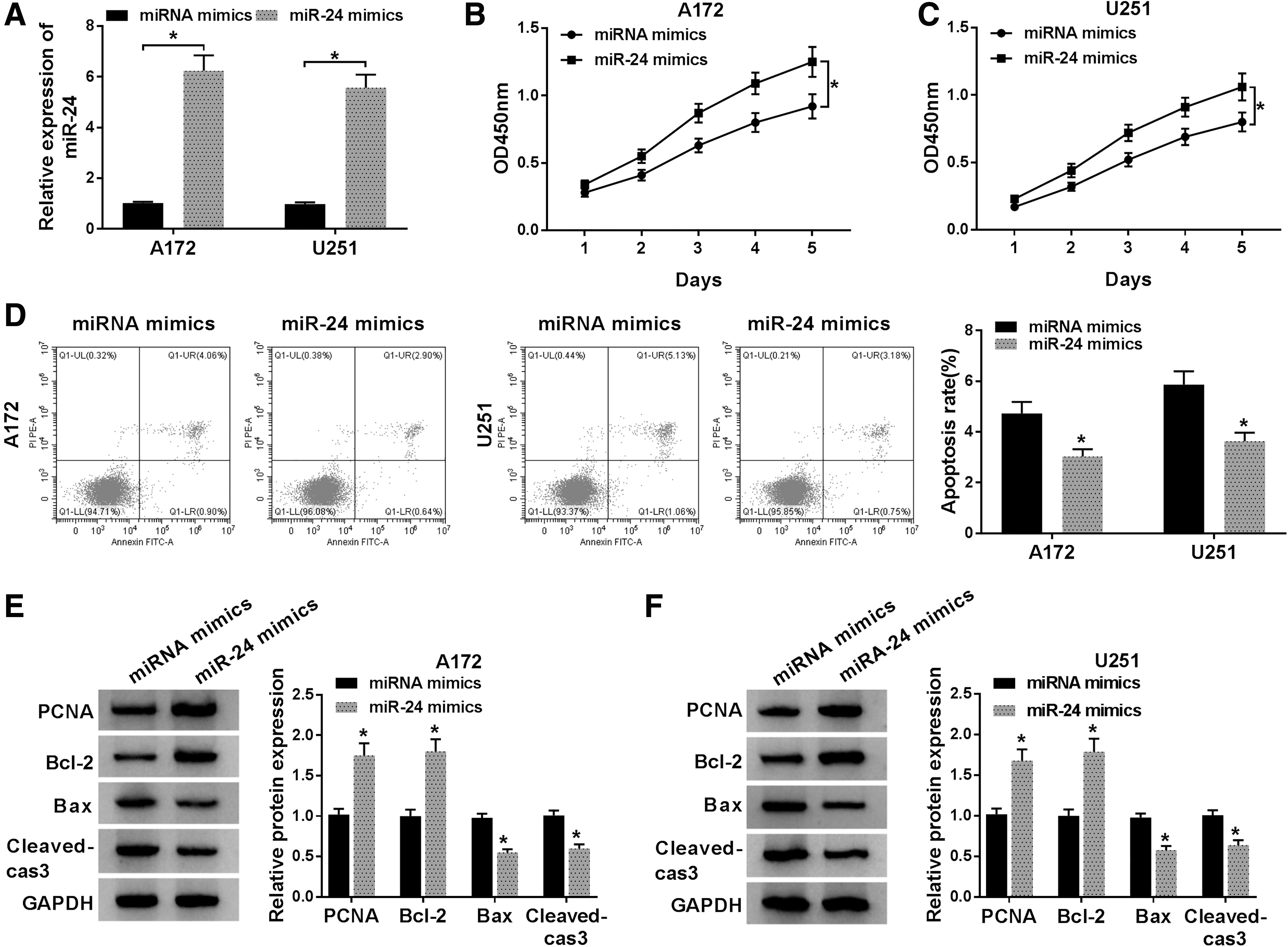
MiR-24 overexpression promotes proliferation and suppresses apoptosis in glioma cells. A172 and U251 cells were transfected with miR-24 mimics or miRNA mimics.
MiR-24 directly targets CDX1 and suppresses its expression
To investigate the underlying molecular mechanism of miR-24 in glioma progression, the potential target genes were predicted according to the microRNA database, and CDX1 was identified that it might be a target of miR-24 with a putative binding site (Fig. 4A). The results of quantitative real-time PCR analysis showed that CDX1 mRNA was markedly downregulated in glioma tissues and cell lines, including A172 and U251 compared to the controls (Fig. 4B, D). Besides, a negative correlation between miR-24 and CDX1 mRNA in glioma also was investigated (Fig. 4C). Then to ascertain whether the same binding site on miR-24 bound to the CDX1 3′-UTR, first, A172 and U251 cells were transfected with vector or CDX1 MUT/WT, and as shown in Supplementary Figure S1, the expression level of CDX1 was significantly elevated in cells, suggesting the transfection of CDX1 MUT/WT was successful. Next, luciferase reporter assay was performed and it was found that the miR-24 mimics reduced the luciferase activities of the CDX1 WT reporter vector but not the CDX1 MUT reporter vector in A172 and U251 cells (Fig. 4E, F). Actually, miR-24 mimic transfection inhibited CDX1 protein expression (Fig. 4G), while miR-24 inhibition promoted CDX1 expression, which was also demonstrated (Fig. 4H). In all, these data showed that miR-24 directly targeted CDX1 and suppressed its expression.
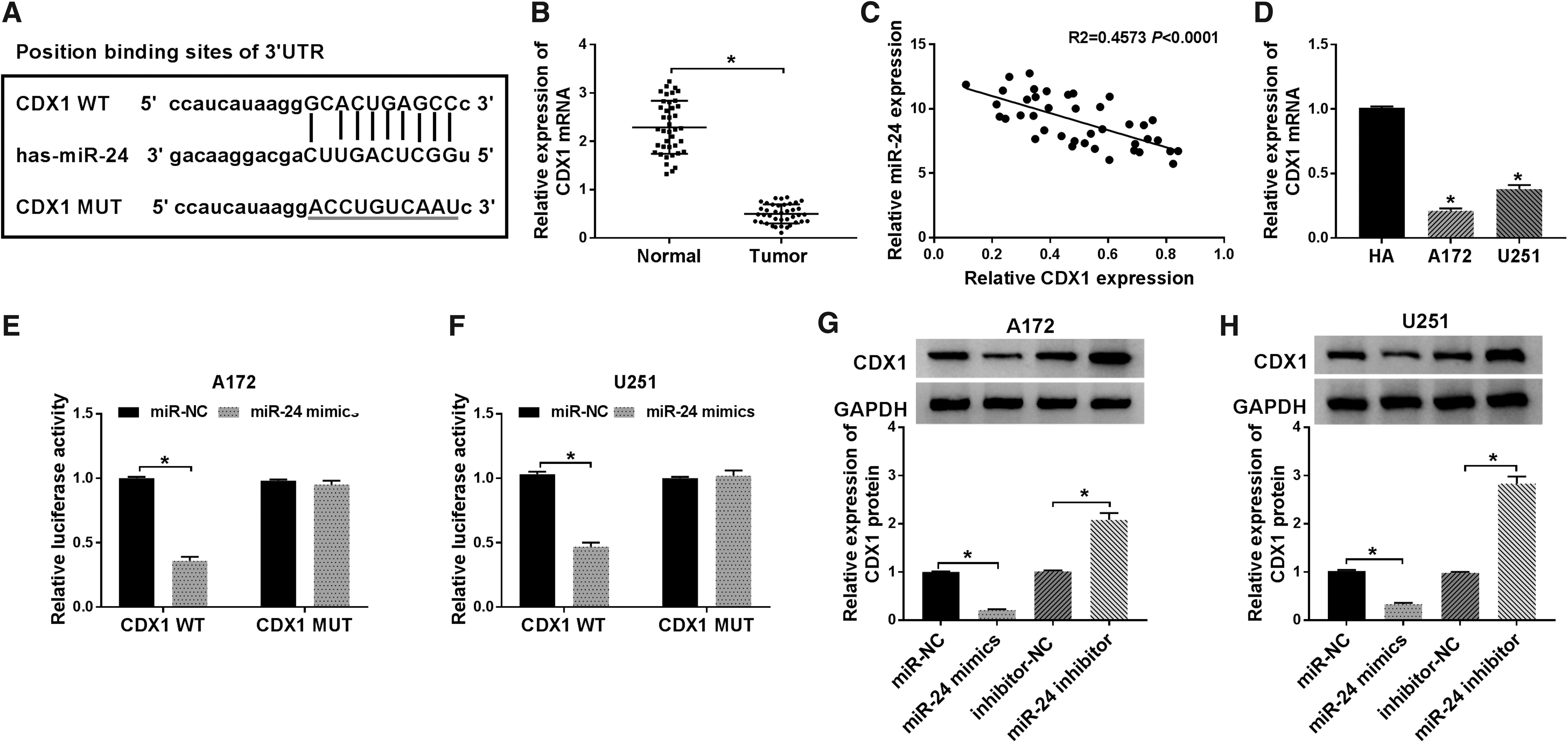
MiR-24 directly targets CDX1 and suppresses its expression.
MiR-24 inhibitor performs antitumor effects on glioma cells by regulating CDX1 expression
Based on the relationship between miR-24 and CDX1, further studies were done whether the effects of miR-24 on glioma cell proliferation and apoptosis were mediated by CDX1. First, A172 and U251 cells were transfected with si-CDX1 or si-NC, and as expected, si-CDX1 transfection significantly reduced the level of CDX1 in A172 and U251 cells (Fig. 5A). Subsequently, A172 and U251 cells were transfected with miR-24 inhibitor, inhibitor NC, si-CDX1+miR-24 inhibitor, or si-NC+miR-24 inhibitor, and it was found that miR-24 inhibitor promoted CDX1, while this promotion was attenuated by CDX1 silence, indicating successful transfection (Fig. 5B). Then cell proliferation and apoptosis rate were evaluated and it was identified that the repression of proliferation and enhancement of apoptosis induced by miR-24 inhibition were significantly attenuated by the knockdown of CDX1 in A172 and U251 cells (Fig. 5C–E). Moreover, Western blot analysis showed CDX1 deletion markedly abated miR-24 inhibition mediated inhibition on PCNA and Bcl-2 expression, and promotion on Bax and Cleaved-cas3 expression in A172 and U251 cells (Fig. 5F). Taken together, the authors confirmed that miR-24 might regulate glioma cell progression by targeting CDX.
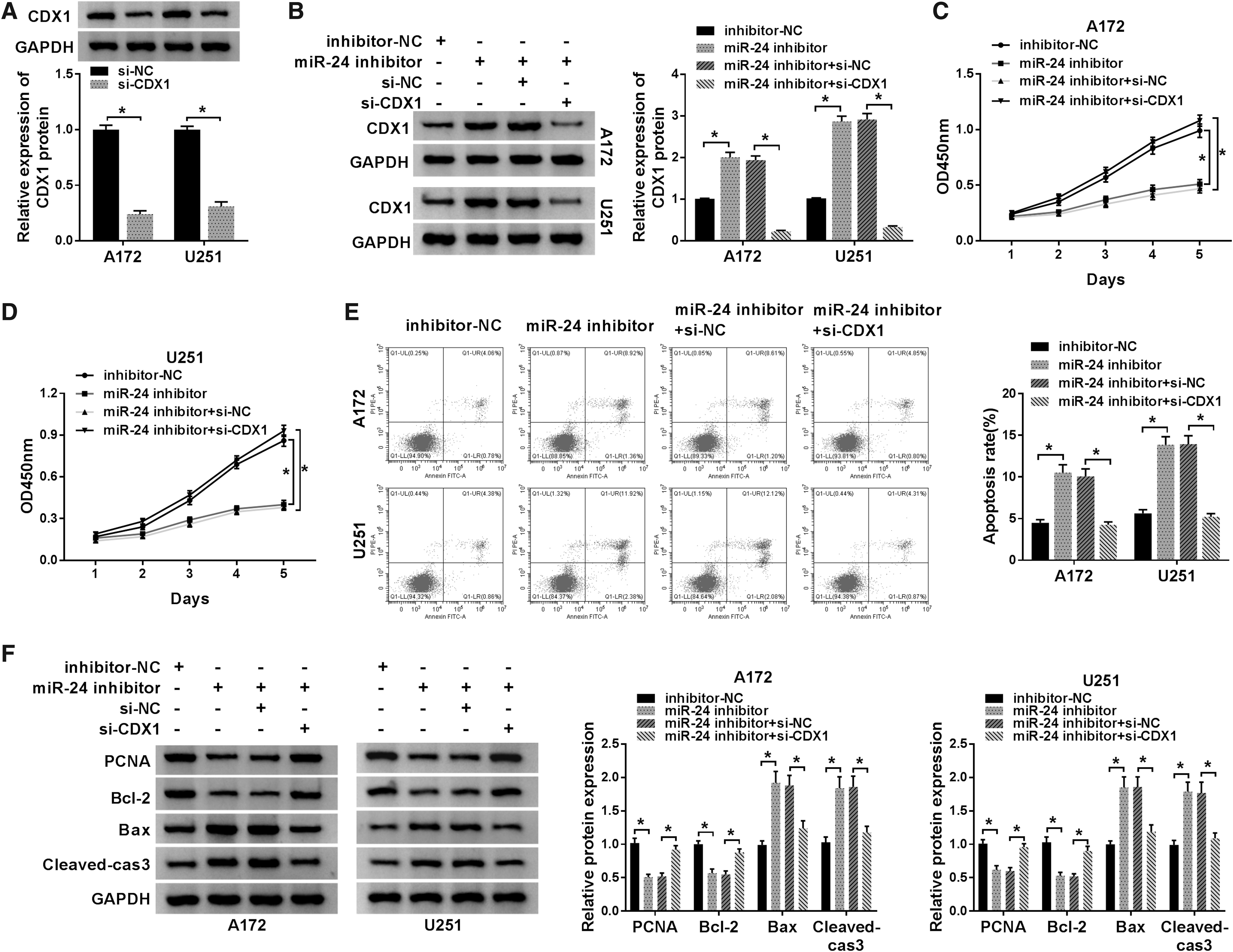
MiR-24 inhibitor performs antitumor effects on glioma by regulating CDX1 expression.
MiR-24/CDX1 can activate the PI3K/Akt signaling pathway in glioma
The PI3K/AKT signaling pathway is a well-known pathway in the regulation of tumorigenesis, and is significantly activated in glioma. 22 To examine if PI3K/Akt signaling implicated in miR-24 mediated glioma cell progression, PI3K, Akt, p-PI3K, p-Akt, Cyclin D1, and p27 levels were analyzed by Western blot and the results indicated miR-24 inhibitor obviously suppressed the level of p-PI3K, p-Akt, and Cyclin D1, but promoted the level of p27 in A172 and U251 cells, while these effects were partially attenuated by CDX1 deletion (Fig. 6A, B), indicating miR-24/CDX1 could activate the PI3K/Akt signaling pathway in glioma.
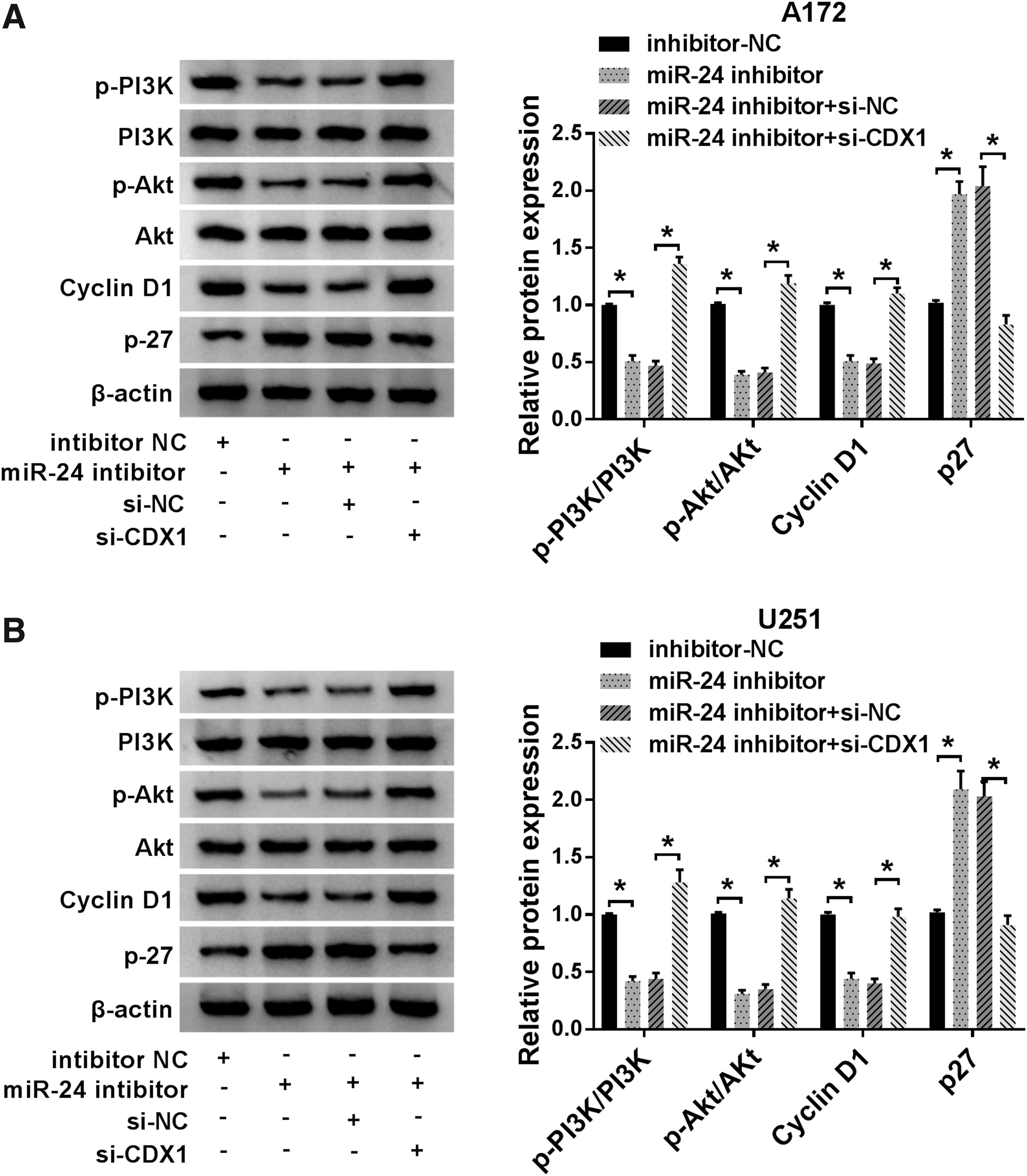
MiR-24 may regulate cell progression by CDX1/PI3K/Akt pathway in glioma.
CDX1 overexpression antagonizes proliferation and induces apoptosis in glioma cells by activating PI3K/Akt signaling pathway
Based on the above results, the authors further detected whether PI3K/AKT pathway involved in the regulation of glioma cell proliferation and apoptosis. Seven hundred forty Y-P, the activator of PI3K, was treated with A172 and U251 cells, as expected, 740 Y-P increased the level of p-PI3K, p-Akt, and Cyclin D1 and decreased the level of p27 (Fig. 7A). After that, A172 and U251 cells were transfected with vector or CDX1, and a notably elevation of CDX1 protein expression was observed (Fig. 7B). Subsequently, rescue assay showed that the activation of PI3K/AKT pathway could reverse CDX1 overexpression induced inhibition on cell proliferation (Fig. 7C, D, F) and enhancement on cell apoptosis (Fig. 7E, F) in glioma. Thus, the authors confirmed that CDX1 overexpression inhibited glioma cell progression by activating PI3K/Akt signaling pathway.
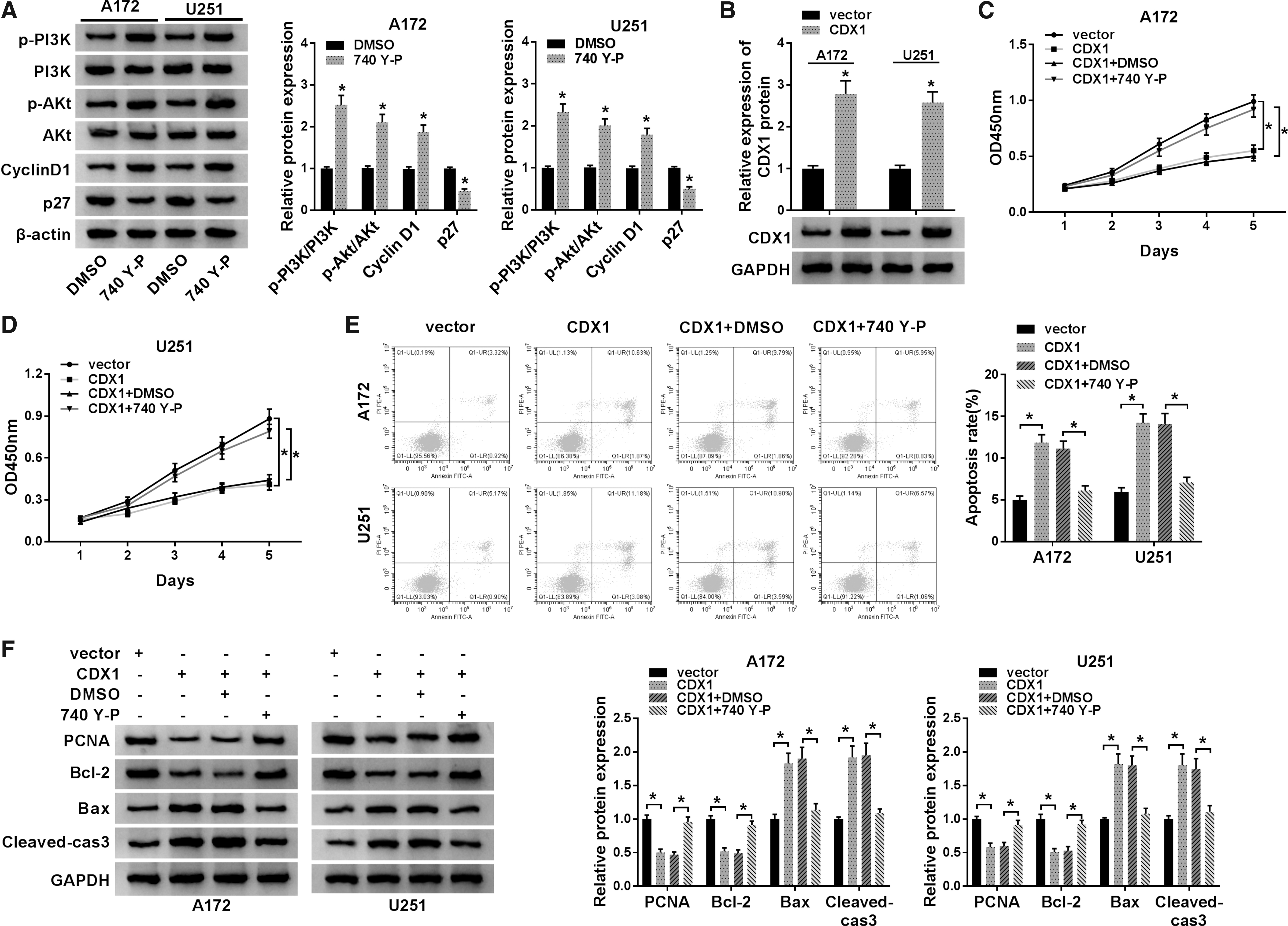
CDX1 overexpression antagonizes proliferation and induces apoptosis in glioma cells by activating PI3K/Akt signaling pathway.
MiR-24 inhibition inhibits glioma tumor growth in vivo
The carcinogenesis roles of miR-24 in vivo were elucidated, and it was found that miR-24 inhibition suppressed NSCLC tumor growth, reflected by the inhibition of tumor volume and weight in miR-24 inhibitor group (Fig. 8A, B). After that, molecular analysis showed that miR-24 inhibition decreased the expression of miR-24, but increased the expression of CDX1 in vivo (Fig. 8C, D). Collectively, these results revealed that miR-24 inhibition hindered glioma tumor growth in vivo through partially regulating CDX1 expression.
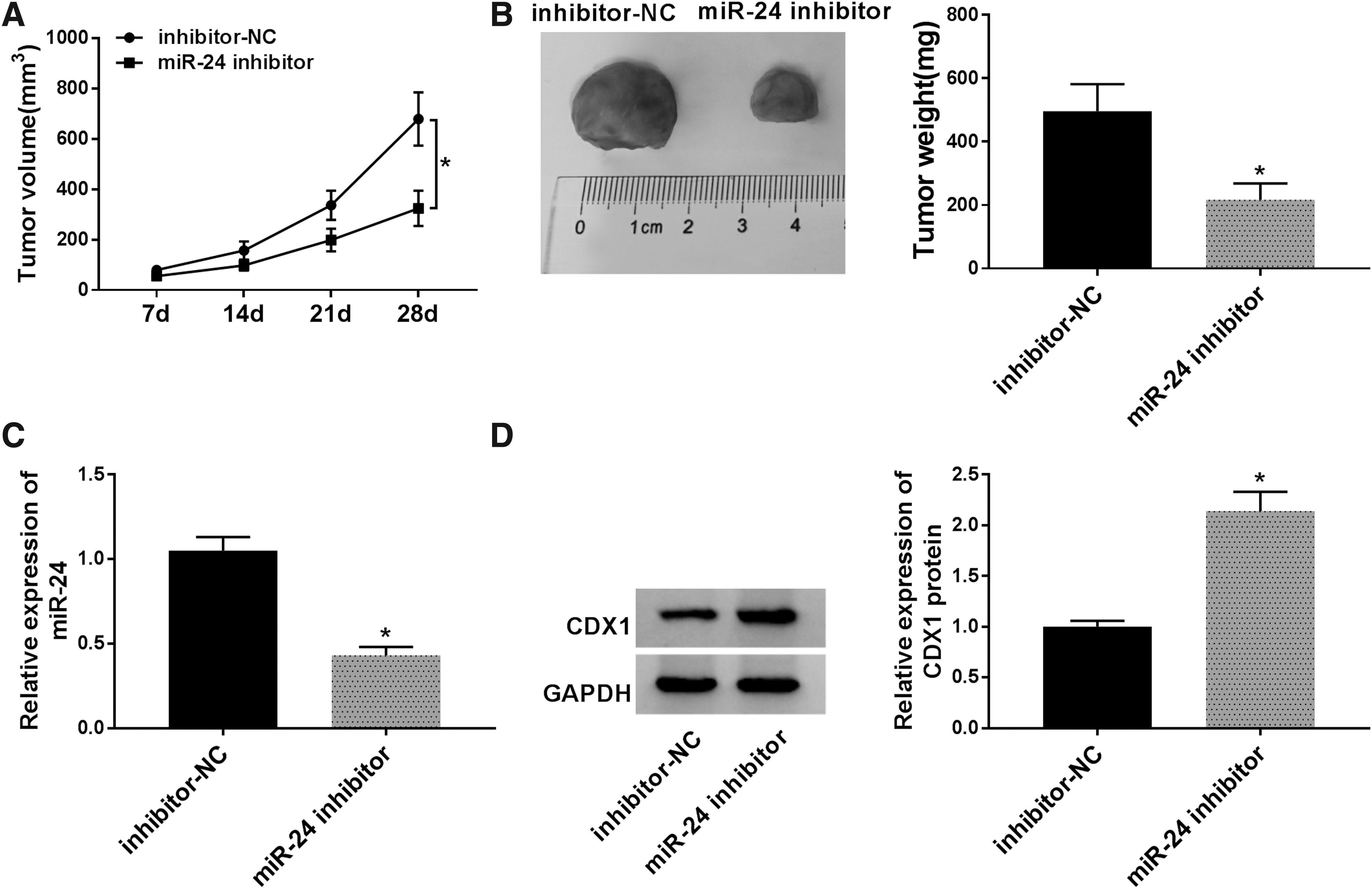
MiR-24 inhibition inhibits glioma tumor growth in vivo.
Discussion
In this study, the authors found that miR-24 was upregulated, and high miR-24 expression independently predicted poor prognosis of glioma patients and was related to the malignancy of glioma. Meanwhile, they also demonstrated that miR-24 functioned as an oncogene to promote tumor growth in vitro and in vivo. In addition, CDX1 was a target of miR-24, and miR-24/CDX1 could activate the PI3K/Akt signaling pathway. Rescue assay indicated that miR-24 could promote glioma progression by facilitating cell proliferation and inducing cell apoptosis through CDX1/PI3K/Akt signaling pathway.
The progression of glioma is a multistep process involving multiple factors. To date, increasing evidence has identified that miRNAs extensively affect the function of genes implicated in gliomagenesis, tumor growth, proliferation, migration, invasion, apoptosis, blood vessel formation around the residual tumor, and posttranscriptional regulation of antioncogenes. As a result, it is suggestive that miRNA profiling may be an effective way to investigate glioma origin, prognosis, and response to therapy. 23,24 Aberrant expression of miR-24 has been determined in numerous tumors, including breast cancer, lung cancer, acute myeloid leukemia, prostate, gastric, colorectal cancer and glioma, and deregulated miR-24 significantly involved in the tumor development. 14,15,25 –30 MiR-24 is a member of the miR-23b cluster, which has been shown to target DNA repair genes H2AX to cause genomic instability and reduce DNA repair. 31 The p53 signaling pathway is an important regulated pathway in the modulation of the cell cycle arrest, differentiation, and DNA repair, and replication. Chen et al. revealed that miR-24 induced cell metastasis and invasion through interacting with p53 in hepatocellular carcinoma. 32 Lu et al. found that miR-24 enhanced lens epithelial cell apoptosis by miR-24-p53 pathway in cataracts. 33 Mori et al. showed that miR-24 could enhance cell proliferation and induce migration of cancer cells by targeting p38, p53, PML, and H2AX simultaneously. 34 Consequently, it seems likely that miR-24 functioned as an oncogene to regulate cell proliferation and apoptosis in glioma.
CDX1 gene belongs to homeobox transcription factor gene family, the tumor-related role of CDX1 has been investigated in various cancers. For example, long noncoding RNA XIST inhibited breast cancer cell growth, migration, and invasion via miR-155/CDX1 axis. 18 CDX2/CDX1 could induce growth inhibition on CDX-deficient gastric cancer by rapid intestinal differentiation. 19 Low-expressed CDX1 was associated with poor prognosis in hepatocellular carcinoma patients after hepatectomy. 35 Therefore, it is of great clinical significance to reveal the role of CDX1 in glioma progression. The PI3K/Akt/PTEN signaling has been validated to be a crucial oncogenic pathway among the dysregulated signaling pathways in glioma. 36 Emerging evidence has shown that the aberrant activity of PI3K/Akt contributes to the cell proliferation, apoptosis, and drug-resistant in glioma, and has been reported to be among the most responsive pathways in association with the pharmacological intervention for glioma treatment. 36 –38 In the current study, p-PI3K, p-Akt, Cyclin D1, and p27 levels were analyzed by using Western blot to explore whether miR-24 exerts biological functions through PI3K/Akt signaling. Subsequently, the results indicated miR-24 inhibitor obviously suppressed the level of p-PI3K, p-Akt, and Cyclin D1, but promoted the level of p27 in glioma cells, while these effects were partially attenuated by CDX1 deletion. In all, these findings indicted that miR-24 could affect the activity of PI3K/Akt pathway by regulating CDX1 in glioma.
Gliomas are generally classified using the WHO system that is largely based on histopathology. However, many recent studies have categorized tumors into multiple molecular classes, and several established molecular markers (MGMT, 1p/19q, IDH, EGFR, and TERT) have been identified to be associated with glioma. These classifications were better able to predict prognosis, survival time, and response to treatment. Although some interesting results were demonstrated in this study, the data of this work presented are based on pathological features. Considering the shortcomings of this study, the molecular analyses are essential to verify these conclusions.
In conclusion, the authors' findings demonstrated that miR-24 was upregulated in glioma, and high miR-24 expression predicted poor prognosis. Besides that, miR-24 could promote cell proliferation and inhibit cell apoptosis in glioma by regulating CDX1/PI3K/Akt pathway in glioma, revealing a novel pathway underlying progression of glioma cells and providing potential targets for glioma treatment. However, there is a limitation of the authors' study, the examination of tumors in vivo should be performed every 3 d, which will make the results more accurate.
Footnotes
Authors' Contributions
D.L. designed the study. H.S. and B.Z. analyzed and interpreted the results. D.L. wrote the article. All authors read and approved the final article.
Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.