
Abstract
Background:
Curcumin is the major component of turmeric, which has an anticancer property in multiple cancers, including hepatocellular carcinoma (HCC). However, the mechanisms are still largely unclear. This research aims to assess the pharmacological function of curcumin and explore the potential microRNA (miRNA)-mRNA regulatory mechanism in curcumin-mediated HCC progression.
Materials and Methods:
Hep3B and Huh-7 cells were used for in vitro experiments. Cells were exposed to various doses of curcumin, and transfection was conducted using Lipofectamine 2000. Cell proliferation, migration, and invasion were examined using 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide or transwell assay. The abundances of miR-21-5p and sex-determining region Y-related high-mobility group box 6 (SOX6) were examined using quantitative real-time polymerase chain reaction or Western blot. The relationship between miR-21-5p and SOX6 was analyzed through luciferase reporter analysis.
Results:
Curcumin repressed HCC cell proliferation, migration, and invasion. miR-21-5p level was decreased in curcumin-treated cells, and miR-21-5p overexpression reversed curcumin-mediated inhibition of HCC progression. SOX6 was targeted through miR-21-5p, and SOX6 restoration attenuated miR-21-5p–induced promotion of HCC progression. Moreover, curcumin exposure increased SOX6 expression through regulating miR-21-5p, and knockdown of SOX6 overturned curcumin-modulated suppression of HCC progression.
Conclusions:
Curcumin repressed proliferation, migration, and invasion of HCC cells by regulating miR-21-5p and SOX6, indicating the promisingly pharmacological effect of curcumin in HCC.
Introduction
Hepatocellular carcinoma (HCC) is an important medical disorder and is the leading cause of cancer-related death in patients. 1 Many strategies have been used for the diagnosis, prognosis, and treatment of HCC patients. 2 However, effective therapeutics for advanced HCC are limited. Hence, much hope is held out for exploring a novel target to therapy of patients with HCC. In recent years, multiple natural plant products have been found to be beneficial for the treatment of HCC. 3 Curcumin is extracted from rhizomes of curcuma longa, which displays anticancer role in many human cancers, such as oral squamous cell carcinoma, glioblastoma, and retinoblastoma.4–6 Moreover, curcumin is found to exhibit hepatoprotective effect on hepatic disorders, including HCC. 7 Nevertheless, the pharmacological effect of curcumin and its potential mechanism remain poorly understood.
microRNA (miRNA) is reported to function as a novel target of curcumin in therapy of cancer. 8 For example, miR-345 and miR-200a/b are associated with the therapeutic effect of curcumin on HCC.9,10 Accruing studies suggest that miR-21-5p can act as a key oncogene to promote progression of human cancers.11–13 More particularly, miR-21-5p level is increased in HCC and regarded as an important marker for the diagnosis of HCC. 14 sex-determining region Y-related high-mobility group box 6 (SOX6), one of the critical members of SOX family, plays an important regulatory role in vertebrates. 15 The emerging evidence suggests that SOX6 is lowly expressed and serves as a promising prognostic biomarker for HCC. 16 We hypothesized that miR-21-5p and SOX6 might be correlated with the underlying mechanism of curcumin in HCC.
In this research, we assessed the influence of curcumin on cell proliferation, migration, and invasion in HCC. Furthermore, we evaluated the role of miR-21-5p and SOX6 in HCC cells, and explored the potential association among curcumin, miR-21-5p, and SOX6.
Materials and Methods
Cell culture, transfection, and treatment of curcumin
Human HCC cell lines (Huh-7 and Hep3B) as well as control liver cell line (HL-7702) (Cell Biology of Chinese Academy of Science, Shanghai, China) were grown in RPMI-1640 medium (HyClone, Logan, UT) with 10% fetal bovine serum (FBS) (HyClone) and 1% penicillin/streptomycin (Beyotime, Shanghai, China) at 37°C with 5% CO2.
Hep3B and Huh-7 cells were added into six-well plates. Once reaching 60% confluence, cells were transfected with miR-21-5p mimic, mimic negative control (miR-NC), miR-21-5p inhibitor (anti-miR-21-5p), inhibitor negative control (anti-miR-NC), pcDNA-SOX6 overexpression vector (pcDNA-SOX6), pcDNA empty vector (pcDNA-control), short hairpin RNA (shRNA) against SOX6 (sh-SOX6), or shRNA negative control (sh-NC) (Genepharma, Shanghai, China) through Lipofectamine 2000 (Thermo Fisher, Waltham, MA) for 24 h. Transfection efficacy was assessed through quantitative real-time polymerase chain reaction (qRT-PCR) or Western blot.
For treatment of curcumin, Hep3B and Huh-7 cells were exposed to different doses (10, 20, and 30 μM) of curcumin (Sigma, St. Louis, MO) before functional analyses. Curcumin was dissolved in dimethyl sulfoxide (DMSO), and cells in corresponding control group were incubated with equal amounts of DMSO.
Cell proliferation, migration and invasion
Cell proliferation was measured through 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2-H-tetrazolium bromide (MTT). Hep3B and Huh-7 cells (5 × 103 per well) were added into 96-well plates, cultured overnight, and exposed to various doses of curcumin for 0, 24, 48, or 72 h, followed by incubating with 1 mg/mL MTT (Sigma) for another 4 h. Subsequently, formazan was solubilized by DMSO, and the absorbance was detected at 490 nm with a microplate reader (BioTek, Winooski, VT).
Cell migration and invasion were detected using 24-well transwell chambers. The transwell chambers were coated with Matrigel (Solarbio, Beijing, China) for invasion analysis, whereas the chambers without Matrigel were used for migration analysis. Hep3B and Huh-7 cells (1 × 104 cells) in medium without serum were added in the upper chambers with or without Matrigel. The lower chambers were added with medium containing 10% FBS. After treatment of curcumin for 24 h, the cells that passed the membranes were dyed with 0.1% crystal violet (Sigma) and observed under a microscope (Nikon, Tokyo, Japan) with three randomly selected fields.
qRT-PCR and Western blot
Cells were incubated with TRIzol reagent (Thermo Fisher) for total RNA isolation. The RNA was employed to synthesize cDNA through TaqMan microRNA reverse transcription kit (Thermo Fisher), and qRT-PCR was conducted on Real-Time PCR Detection System (Bio-Rad, Hercules, CA). The relative RNA level was measured with U6 or GAPDH as reference by the 2−ΔΔCt method. 17 The specific primers for miR-21-5p or U6 were provided through RiboBio (Guangzhou, China), and the primers for SOX6 or GAPDH are listed as follows: SOX6 forward primer: 5′-TTACCAGTGATTTCTGGGTGCT-3′, reverse primer: 5′-GCTGCTGTTGTCTCGCAATC-3′; GAPDH forward primer: 5′-GAAAGCCTGCCGGTGACTAA-3′, reverse primer: 5′-TTCCCGTTCTCAGCCTTGAC-3′.
For detection of protein level, total protein was isolated using RIPA buffer and quantified by BCA kit (Solarbio). Sodium dodecyl sulfate polyacrylamide gel electrophoresis was performed to separate protein, which was then transferred to nitrocellulose membranes (Bio-Rad). After blocking in 5% skim milk, membranes were interacted with primary antibodies for SOX6 (ab64946; Abcam, Cambridge, MA) or GAPDH (ab181602; Abcam) overnight and then incubated with IgG (ab97051; Abcam). After exposing to BeyoECL Plus (Beyotime), relative expression of SOX6 was measured through Image Lab software (Bio-Rad) with GAPDH as a reference.
Luciferase reporter analysis
The 3′ untranslated region (3′ UTR) fragments of SOX6 having binding sites of miR-21-5p predicted through TargetScan were amplified and inserting into psiCHECK-2 vectors (Promega, Madison, WI) to form wild-type luciferase reporter construct SOX6 3′ UTR-WT. The mutant-type SOX6 3′ UTR-MUT with the mutated seed sites was formed through a site-directed mutagenesis kit (Thermo Fisher). Hep3B and Huh-7 cells were cotransfected with anti-miR-21-5p or anti-miR-NC and corresponding luciferase reporter constructs for 24 h. A dual-luciferase assay system (Promega) was used for detection of luciferase activity.
Statistical analysis
Statistical analysis was conducted through GraphPad Prism 7 (GraphPad, Inc., San Diego, CA). All results are shown as the means ± standard deviation based on three independent experiments. The difference was compared through Student's t-test for two groups or analysis of variance with Dunnett's test for multiple groups. The difference was statistically significant at p < 0.05.
Results
Curcumin inhibits proliferation, migration, and invasion of HCC cells
To probe the potential role of curcumin in HCC progression, Hep3B and Huh-7 cells were stimulated by different doses of curcumin for 24, 48, or 72 h. The data of MTT assay displayed that exposure to curcumin suppressed cell proliferation in a concentration-dependent manner (Fig. 1A, B). In addition, analysis of transwell showed that the migration ability was markedly impaired in Hep3B and Huh-7 cells after treatment of curcumin for 24 h (Fig. 1C, D). Similarly, insult of curcumin also led to progressive loss of invasive ability in the two cell lines (Fig. 1E, F).
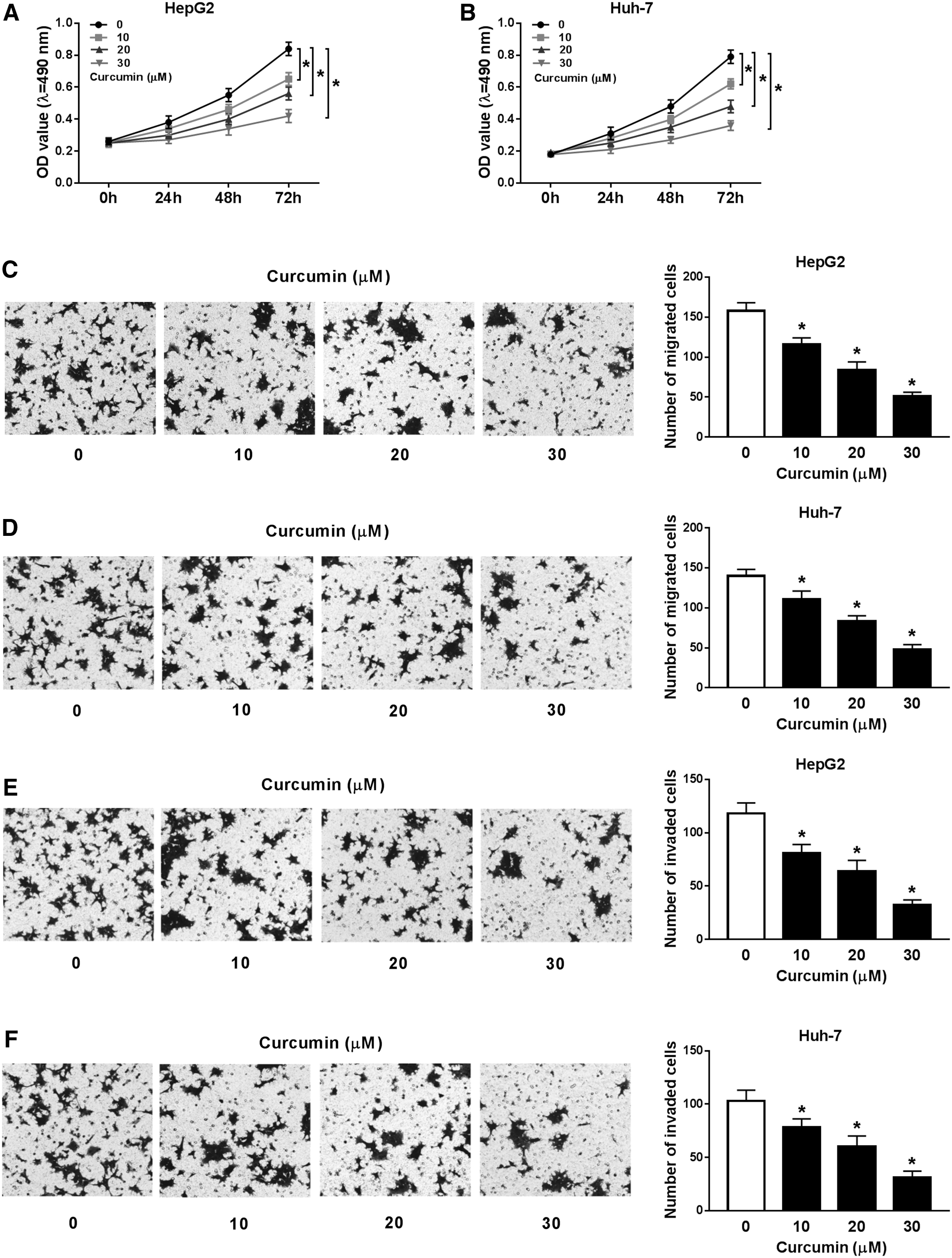
Influence of curcumin on proliferation, migration, and invasion of HCC cells.
Overexpression of miR-21-5p reverses curcumin-mediated inhibition of HCC progression
To study the function of miR-21-5p, its level was detected in HCC cells. Results showed that miR-21-5p expression was obviously enhanced in Hep3B and Huh-7 cells compared with that in HL-7702 cells (Fig. 2A). However, after treatment of curcumin, miR-21-5p abundance was progressively reduced in a concentration-dependent manner (Fig. 2B). To assess whether miR-21-5p was responsible for curcumin-mediated progression, Hep3B and Huh-7 cells were transfected with miR-21-5p mimic or miR-NC before exposure to 20 μM of curcumin. Transfection efficacy was identified with the results of upregulated miR-21-5p level in miR-21-5p mimic-transfected cells (Fig. 2C). Furthermore, the data of MTT and transwell analyses demonstrated that addition of miR-21-5p weakened curcumin-modulated inhibition of proliferation, migration, and invasion in Hep3B and Huh-7 cells (Fig. 2D–G).
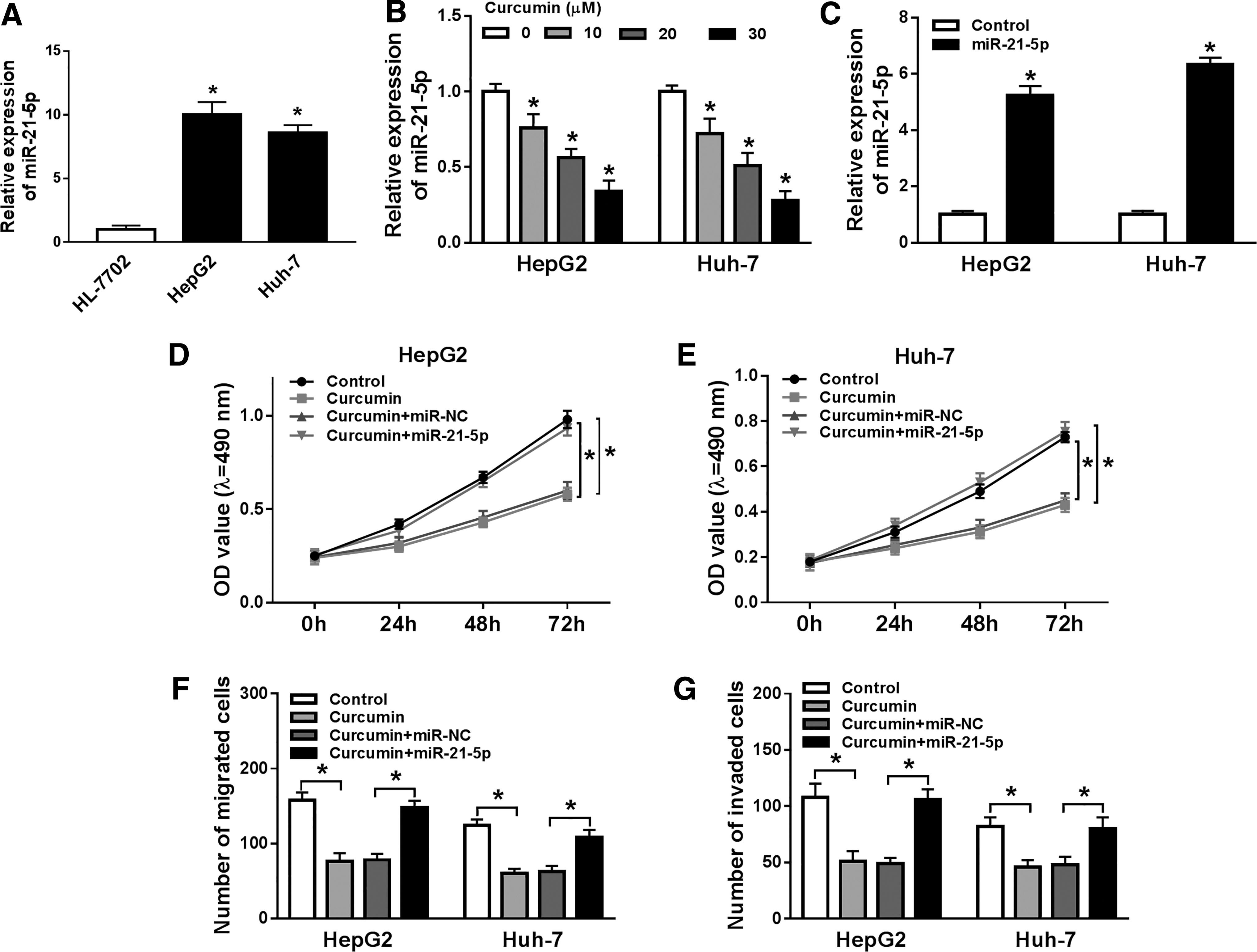
Effect of miR-21-5p on curcumin-mediated HCC progression.
SOX-6 is targeted and regulated through miR-21-5p in HCC cells
Bioinformatics analysis displayed the predicted complementary sites of miR-21-5p and SOX6 (Fig. 3A), suggesting that SOX6 might be targeted by miR-21-5p. To confirm this relationship, wild-type or mutant luciferase reporter vector of SOX6 was constructed. Luciferase reporter assay exhibited that luciferase activity was markedly enhanced in Hep3B and Huh-7 cells transfected with SOX6 3′ UTR-WT and anti-miR-21-5p in comparison with the miR-NC-transfected group, whereas it was not affected relative to SOX6 3′ UTR-MUT–treated cells (Fig. 3B, C). Moreover, the influence of miR-21-5p on SOX6 abundance was assessed in the two cell lines with transfection of anti-miR-21-5p or anti-miR-NC. The data displayed that SOX6 abundance was evidently elevated at mRNA and protein levels by miR-21-5p inhibition (Fig. 3D, E).
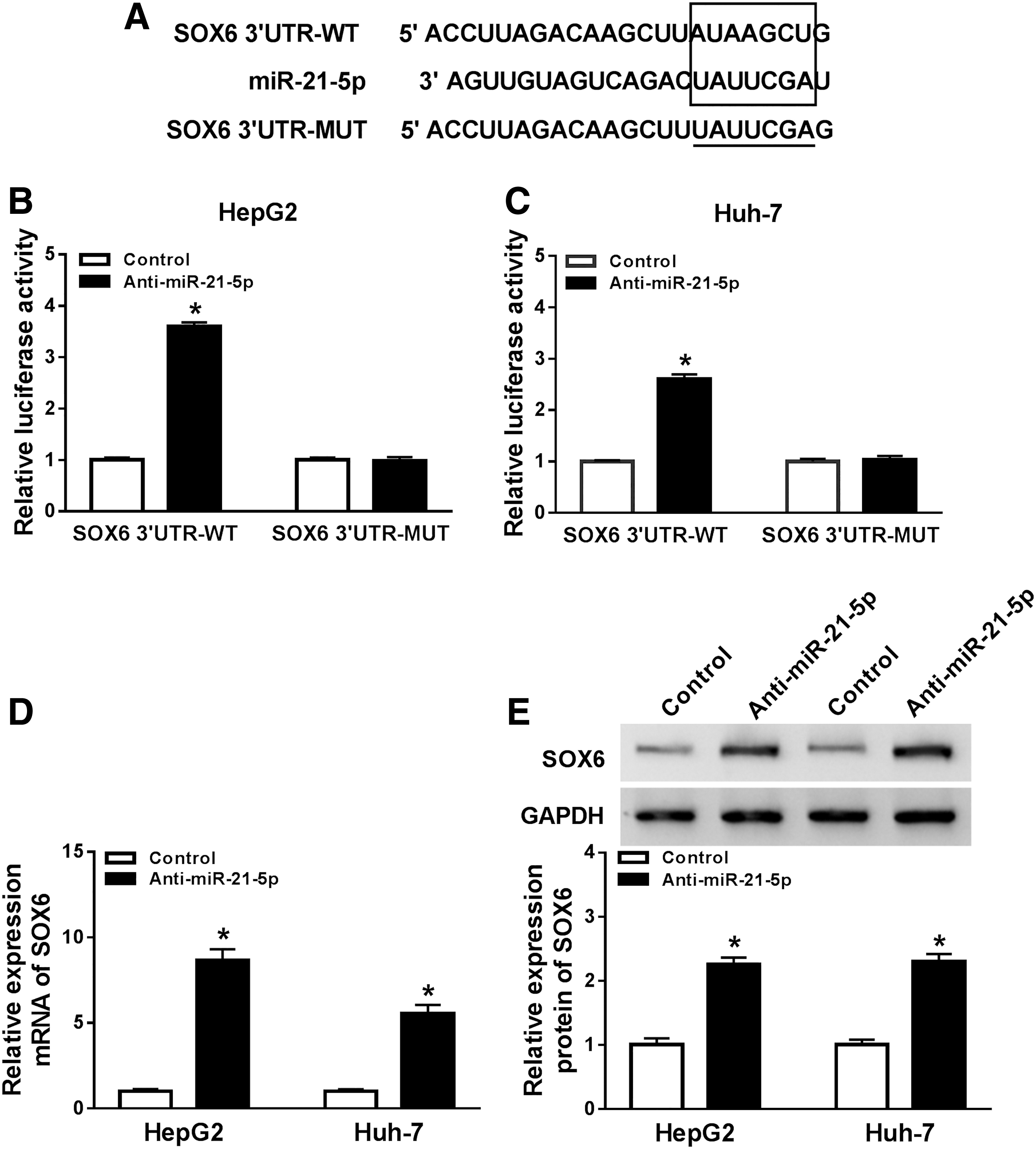
Association between miR-21-5p and SOX6.
Addition of miR-21-5p facilitates proliferation, migration, and invasion of HCC cells through regulating SOX6
To analyze whether miR-21-5p–mediated HCC progression was modulated by SOX6, Hep3B and Huh-7 cells were transfected with miR-NC, miR-21-5p mimic, miR-21-5 mimic + pcDNA-control, or pcDNA-SOX6. Results showed that SOX6 expression was obviously enhanced at mRNA and protein levels by introduction of pcDNA-SOX6 (Fig. 4A, B). Moreover, MTT assay revealed miR-21-5p upregulation promoted cell proliferation, which was reversed by accumulation of SOX6 (Fig. 4C, D). Furthermore, the data of transwell analysis demonstrated that miR-21-5p overexpression obviously promoted migration and invasion of Hep3B and Huh-7 cells, whereas this effect was alleviated by restoration of SOX6 (Fig. 4E, F).
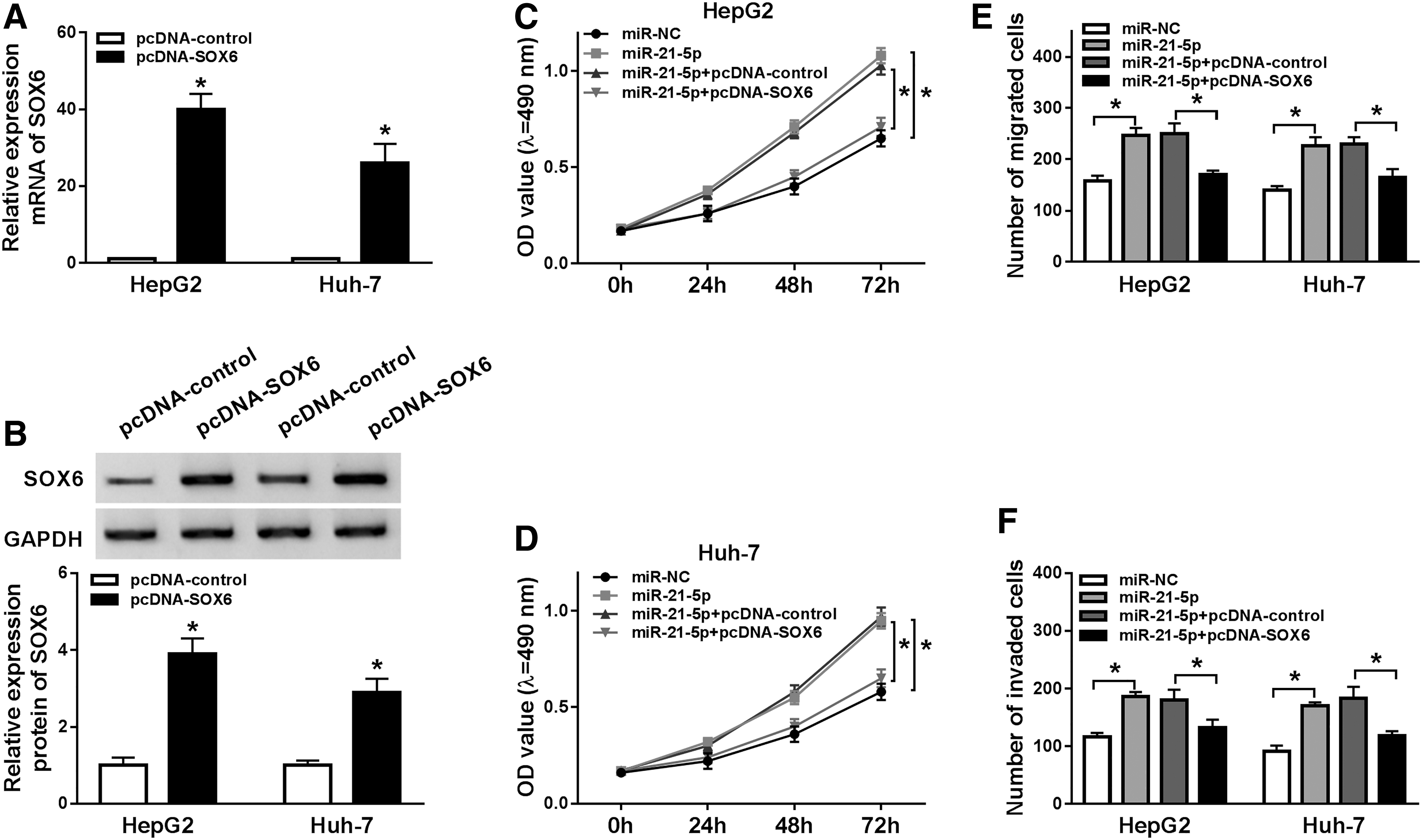
Effect of SOX6 restoration on miR-21-5p–mediated HCC progression.
Curcumin inhibits HCC progression by regulating miR-21-5p and SOX6
To explore the association between SOX6 and curcumin-mediated HCC progression, the level of SOX6 was detected in Hep3B and Huh-7 cells with transfection of miR-21-5p mimic or miR-NC and treatment of curcumin. The results displayed that SOX6 mRNA and protein levels were remarkably enhanced through exposure to curcumin in the two cell lines, which was notably weakened by miR-21-5p overexpression (Fig. 5A–D). In addition, Hep3B and Huh-7 cells were transfected with sh-SOX6 or sh-NC and exposed to curcumin. Silence of SOX6 overturned the suppressive role of curcumin in cell proliferation, migration, and invasion (Fig. 5E–H).
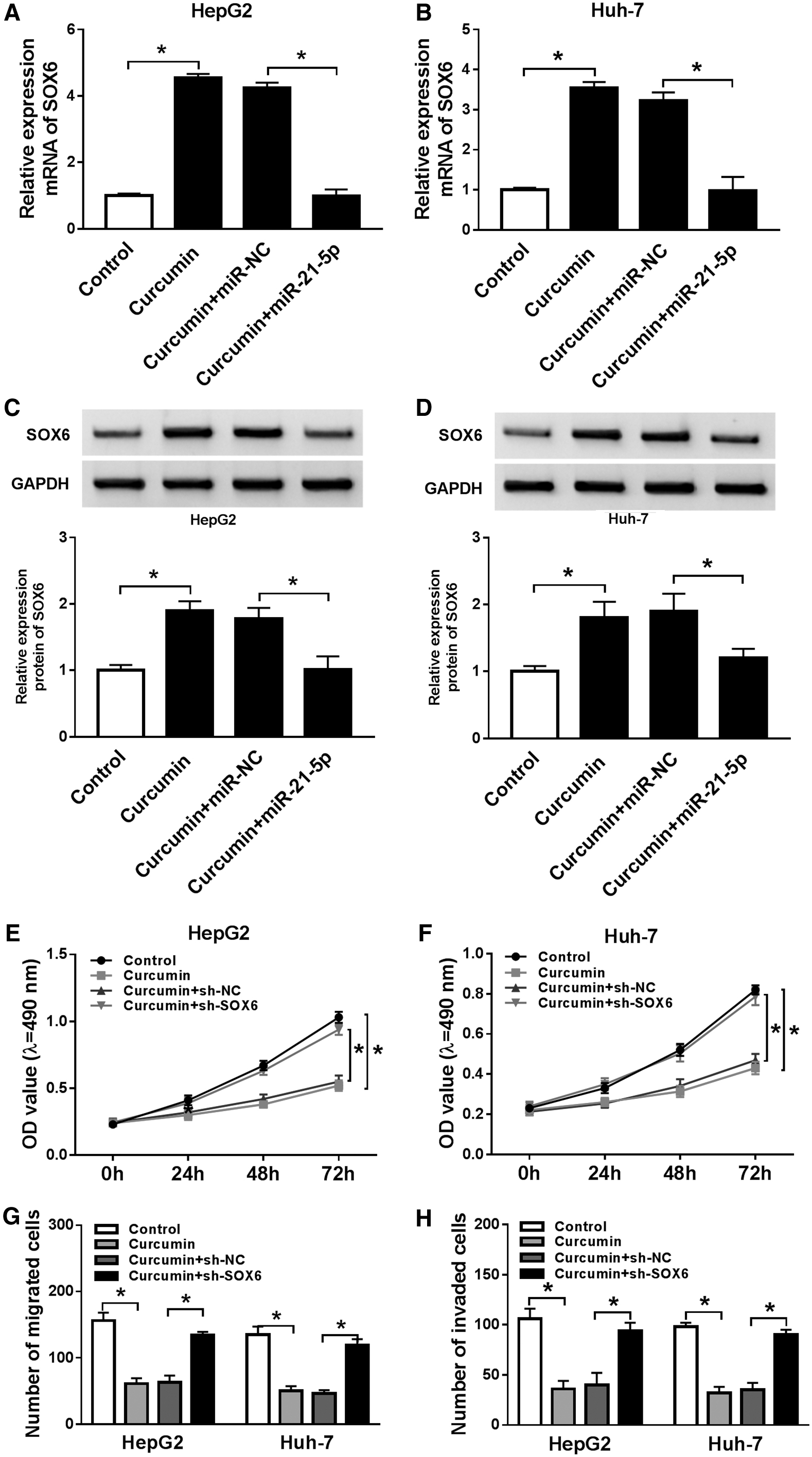
Effect of SOX6 on curcumin-mediated HCC progression.
Discussion
HCC is the main type of liver malignancies with high incidence worldwide. Curcumin, the main active compound of turmeric, has been widely regarded as promisingly traditional medicine in several cancers. A previous study implied the hepatoprotective effect of curcumin on HCC by regulating autophagy and apoptosis. 18 However, further study is warranted to better define the underlying mechanism of curcumin. In this research, the pharmacological function of curcumin in cell proliferation, migration, and invasion was assessed, and the crosstalk between curcumin and miR-21-5p/SOX6 axis in HCC cells was first addressed.
This study revealed that curcumin treatment inhibited HCC cell proliferation, which is also consistent with a previous finding that suggested the antiproliferative role of curcumin. 19 Hosseini et al. reported that curcumin could inhibit HCC cell viability through decreasing cyclin D1 expression. 20 Moreover, You et al. and Pan et al. also showed that curcumin could suppress HCC growth by regulating sterol regulatory element binding transcription factor 1 or vascular endothelial growth factor.21,22 Interestingly, curcumin derivative WZ35 and its analogue diphenyl difluoroketone have been shown to repress invasion and migration of HCC cells,23,24 suggesting the antimetastatic role of curcumin. Unsurprisingly, it was also found that curcumin blocked migration and invasion of HCC cells. Collectively, curcumin could be used as an optional chemotherapeutic agent for HCC treatment.
A previous study reported that curcumin could play an anticancer effect by regulating miRNA expression in many caners. 25 Multiple pieces evidence described that curcumin could decrease miR-21 abundance in colorectal cancer, nonsmall cell lung cancer, breast cancer, and gastric cancer.26–29 Nevertheless, it could not indicate the relationship between curcumin and miR-21-5p in HCC due to the alteration of tumor microenvironment. Previous studies suggested that miR-21-5p abundance was enhanced in serum or tissues of HCC patients.14,30 Consistent with these studies, this study also showed that miR-21-5p was highly expressed in HCC cells and miR-21-5p overexpression promoted HCC progression, suggesting that miR-21-5p played as an oncogene in HCC development. Moreover, miR-21-5p level was decreased through curcumin and accumulation of miR-21-5p counteracted the inhibitive role of curcumin, indicating downregulation of miR-21-5p summarized a new mechanism for the antitumor role of curcumin in HCC. One of the underlying mechanisms allows the function of miRNA is known to regulate its potential target. In this study it was found that depletion of miR-21-5p could increase the luciferase activity of SOX6 3′ UTR-WT constructs and SOX6 abundance in HCC cells, revealing that SOX6 was directly targeted by miR-21-5p, which was also consistent with a previous study. 31
Xie et al. demonstrated that miR-155 promoted tumorigenesis of HCC by targeting SOX6. 32 Li et al. showed that miR-96 increased cell proliferation, migration, and invasion through inhibiting SOX6 in HCC. 33 These studies suggested that SOX6 might serve as a tumor suppressor in HCC development. Similarly, this study uncovered that SOX6 restoration mitigated miR-21-5p–mediated promotion of proliferation, migration, and invasion, implying the anticancer role of SOX6, and that miR-21-5p contributed to HCC progression by targeting SOX6. Moreover, SOX6 expression was enhanced by curcumin through regulating miR-21-5p, and knockdown of SOX6 reversed the anticancer effect of curcumin on HCC, indicating the importance of miR-21-5p/SOX6 axis on the mechanism of curcumin. This study indicated curcumin as a promisingly chemopreventive and therapeutic agent in HCC in vitro. However, the pharmacologic effect of curcumin is limited because of the poor solubility in water and low bioavailability in vivo. 34 Some drug delivery systems, such as nanoparticle, 35 might contribute to improving bioavailability of curcumin. Hence, multidrug combination therapy should be analyzed for better clinical application of curcumin in future.
In conclusion, the data of this study revealed that curcumin impeded malignancy of HCC in vitro by inhibiting cell proliferation, migration, and invasion. Moreover, it was first confirmed that this antitumor role was associated with miR-21-5p/SOX6 axis. This indicated the pharmacological effect of curcumin in HCC cells, suggesting a promising avenue for therapeutics of HCC.
Footnotes
Funding Information
This study was supported by the Natural Science Foundation of Zhejiang Province (Grant No. LY15H030002).
Disclosure Statement
There are no existing financial conflicts.