
Abstract
It is now well understood that many signaling pathways are vital in carrying out and controlling essential pro-survival and pro-growth cellular functions. The NOTCH signaling pathway, a highly conserved evolutionary signaling pathway, has been thoroughly studied since the discovery of NOTCH phenotypes about 100 years ago in Drosophila melanogaster. Abnormal NOTCH signaling has been linked to the pathophysiology of several diseases, notably cancer. In tumorigenesis, NOTCH plays the role of a “double-edged sword,” that is, it may act as an oncogene or as a tumor suppressor gene depending on the nature of the context. However, its involvement in several cancers and inhibition of the same provides targeted therapy for the management of cancer. The use of gamma (γ)-secretase inhibitors and monoclonal antibodies for cancer treatment involved NOTCH receptors inhibition, leading to the possibility of a targeted approach for cancer treatment. Likewise, several natural compounds, including curcumin, resveratrol, diallyl sulfide, and genistein, also play a dynamic role in the management of cancer by inhibition of NOTCH receptors. This review outlines the functions and structure of NOTCH receptors and their associated ligands with the mechanism of the signaling pathway. In addition, it also emphasizes the role of NOTCH-targeted nanomedicine in various cancer treatment strategies.
Introduction
The NOTCH signaling pathway was initially discovered through genetic studies in the Drosophila melanogaster in 1910s. It is a cell-to-cell contact mechanism between adjacent cells that serves a crucial role in the development of mammalian species. NOTCH is a juxtracrine signaling mechanism in which membrane-bound NOTCH ligands activate NOTCH receptors on adjacent cells. 1 They play an important role in the regulation of embryonic development as well as the transmission of cell-to-cell signals. They govern what occurs to cells through managing cell differentiation, death, and proliferation. 2
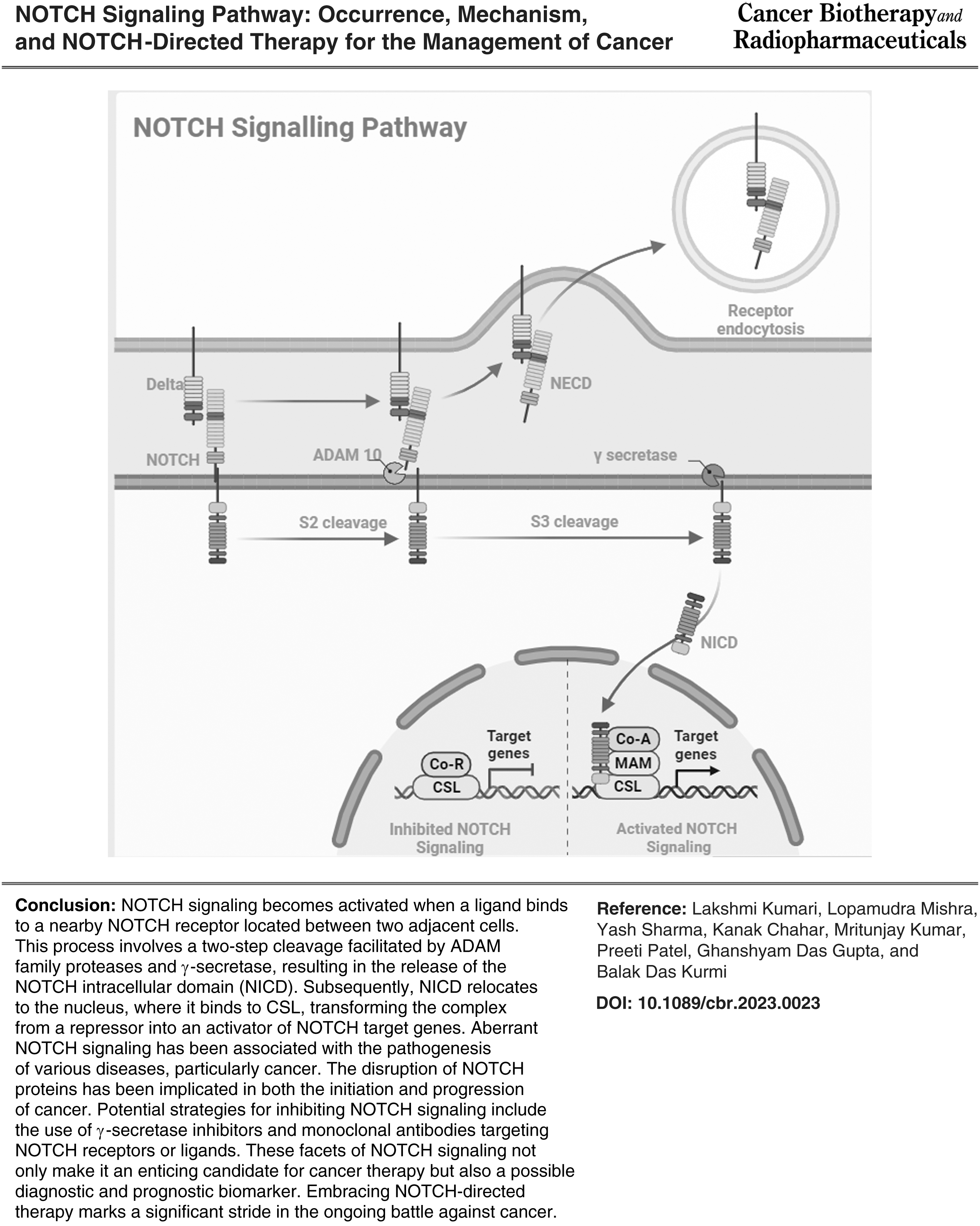
The NOTCH receptor is composed of an intracellular portion that carries the signal, a transmembrane region, and an extracellular portion (with numerous tandem epidermal growth factors (EGF)-like repeats) that binds to ligands produced on a neighboring cell as shown in Figure 1. Despite the fact that the majority of members of the NOTCH receptor family are often implicated in many malignancies, targeting particular NOTCH receptors offers several advantages, including decreased toxicity and higher impact on specific receptors that are involved in several kinds of cancer.
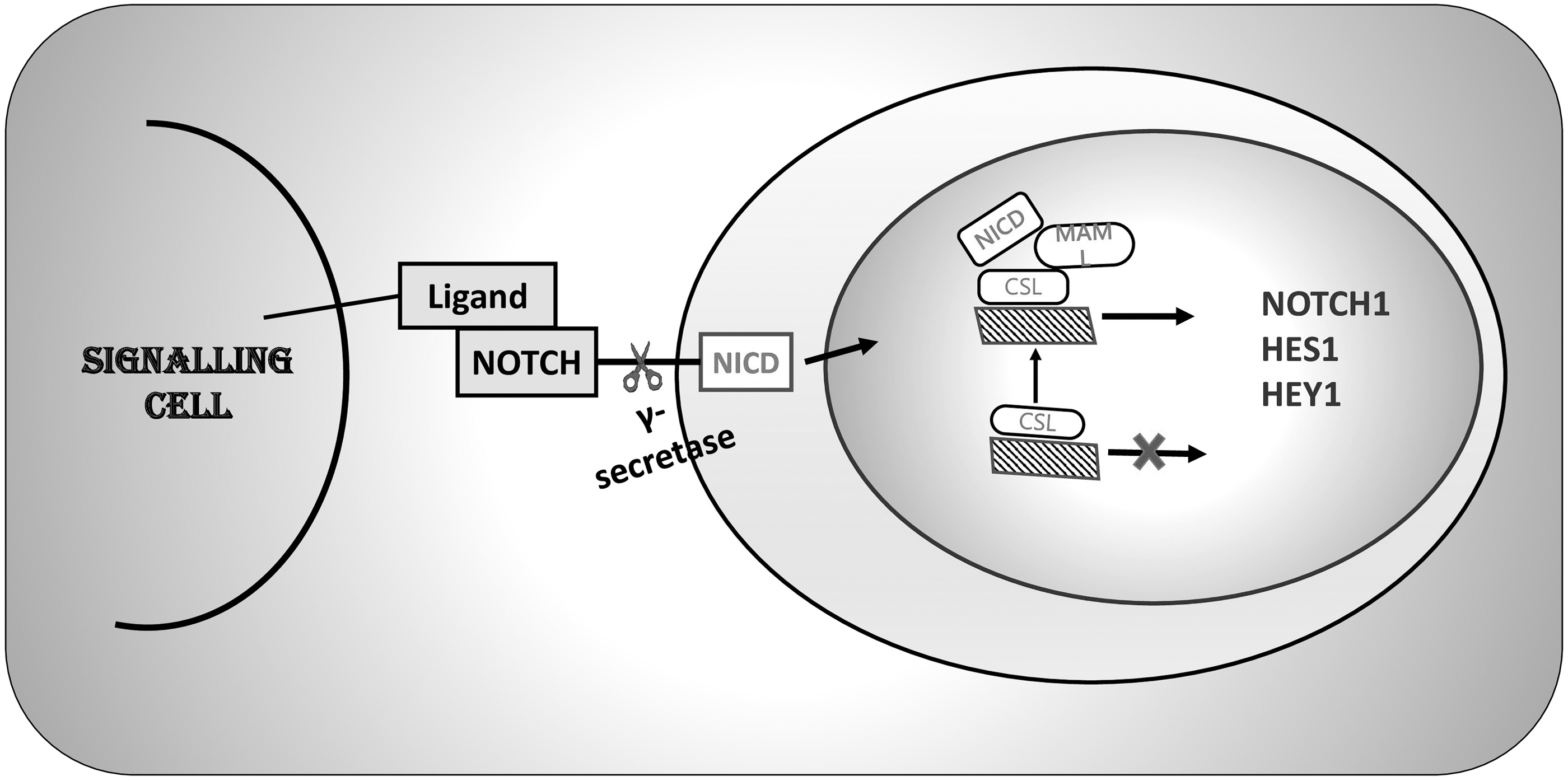
Basic elements of NOTCH Signaling pathway. NICD, NOTCH intracellular domain; CSL, C protein binding factor 1/Suppressor of Hairless/Lag-1; MAML1, mastermind-like 1; HES1, hairy and enhancer of split-1.
Numerous malignancies, such as T cell acute lymphoblastic leukemia (T-ALL) and triple-negative breast cancer (TNBC), have been linked to NOTCH1 activating mutations. 3 The NOTCH pathway has a substantial effect on various kinds of cancer, and abnormalities in NOTCH signaling lead to many of the tumor's distinguishing characteristics. The NOTCH signaling pathway's function in cancer ranges from tumor-suppressive to malignant. 4
History of NOTCH Signaling Pathway
For the very first time, in 1913, investigation of mutant D. melanogaster with NOTCH wings, the NOTCH gene was originally identified by Thomas Hunt Morgan. D. melanogaster developed NOTCH at the tips of its wings due to the haploinsufficiency of NOTCH. 5,6 The latter suggested that nomenclature was motivated by the identification of this trait.
The first allele of NOTCH was established in 1917. 6 After the isolation and sequencing of the D. melanogaster NOTCH gene in the 1980s, it was discovered that the putative NOTCH protein spans the membrane and has several repetitions that are like the EGFR. Moreover, as research on NOTCH signaling in D. melanogaster raised, the entire signaling pathway came into focus.
Further, in Caenorhabditis elegans (C. elegans), LIN-12 and GLP-1 were discovered as NOTCH homologs in 1988 and 1989, and they are believed to be linked to C. elegans development. Around 1990, “XOTCH,” a Xenopus molecule remarkably like Drosophila NOTCH in both structure and developmental expression, was found. Ever since, interest to research NOTCH in other species has risen and NOTCH signaling has been better understood, and as a result, it is now acknowledged as a highly classic and conserved signaling pathway. 5,7 NOTCH signaling was connected to cancer for the first time in 1991, and human T-ALL was one of the first examples in which a correlation between the NOTCH gene and cancer was discovered, where the t(7;9) (q34;q34.3) chromosomal translocation was related to the NOTCH-1 gene. 8
Here, we briefly review the occurrence and evolution of the NOTCH signaling pathway, structure, as well as the functions of its receptors and ligands, and the key steps involved in the mechanism and then turn to the roles of NOTCH in several cancers, including breast cancer. The present review additionally places focus on various targeted therapies that block the NOTCH pathway to treat cancer.
The Architecture of NOTCH Signaling
Structure and function of NOTCH receptors and ligands
From sea urchins to humans, the NOTCH receptor has evolved into a single pass trans-membrane protein. Originally, NOTCH receptors were recognized as causing a particular “NOTCH” shaped phenotype on the D. melanogaster's wing structure. 9 Vis a vis to Drosophila, which only expresses one NOTCH receptor, the mammalian NOTCH family contains four members (NOTCH1, 2, 3, and 4), 10,11 which are around 60% identical to each other and to Drosophila NOTCH.
There are notable differences across NOTCH receptors, even though their general structure is similar. NOTCH receptors are made up of three domains: an extracellular domain (NECD), a transmembrane domain (NTM), and an intracellular domain (NICD). 12,13 The extracellular subunit of NOTCH is endowed with many interchangeable structures containing numerous Ca2+ binding EGF-like repeats that are required for ligand interactions and a negative regulatory region (NRR). 14,15
Three cysteine-rich Lin12/NOTCH repeats (LN), each with a Ca2+ binding site, comprise an NRR. 16,17 The transmembrane domain that is conjectured to be involved in heterodimerizations are significant for S2 cleavage, including a short extracellular area encompassing a pair of preserved cysteines. 18 The NICD includes an RBP-jk association molecule domain, two nuclear localization signals, seven ankyrin (ANK) repeats, a trans-activation domain, which ends with a polyglutamine region (OPA), and a C-terminal “PEST sequence” (a region rich in proline (P), glutamic acid (E), serine (S), and threonine (T) residues that comprise degradation signals and are, therefore, responsible for the stability of the NICD and subsequently become a reason for its ubiquitination). 14
The extracellular domains of the NOTCH1 and NOTCH2 receptors feature 36 EGF-like repeats, similar to Drosophila, but NOTCH3 and NOTCH4 have 34 and 29 repeats, respectively. 14,19
In contrast to Drosophila who expresses two canonical NOTCH ligands named as Delta and Serrate, mammals have five NOTCH ligands: three ligands, namely Delta-like-1, -3, and -4 (DLL1, DLL3, and DLL4) that are homologous to Delta ligand and others two are Jagged1 and Jagged2 (JAG1 and JAG2) which are homologous to Serrate ligand. 20,21 In the context of NOTCH activities, DLL1 regulates cell-to-cell communication and cell differentiation, whereas DLL3 suppresses cell proliferation via activating receptors. 22 Figure 2 displays the structural elucidation of receptors and ligands of NOTCH pathway.
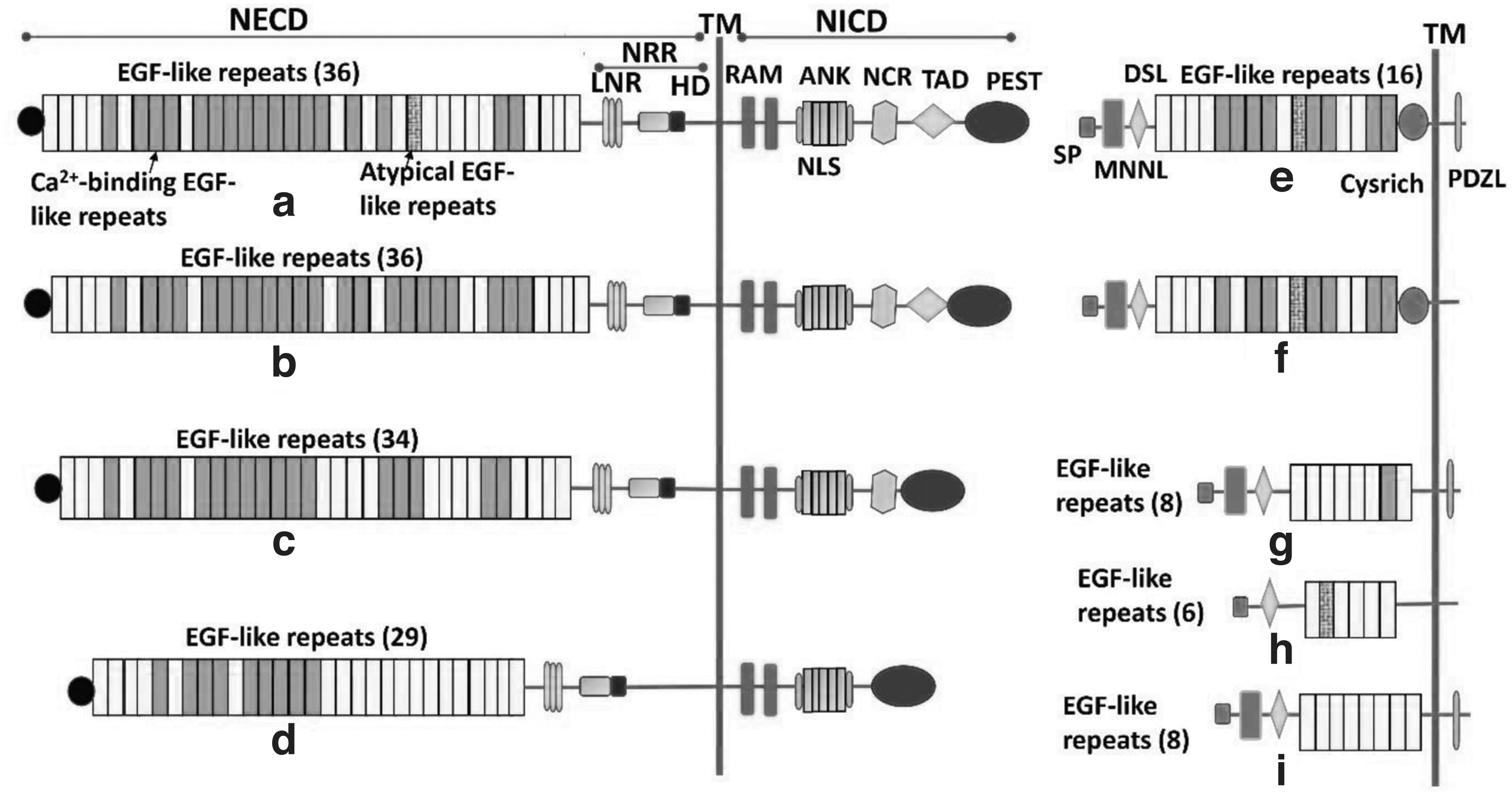
Schematic illustration of receptors and ligands of NOTCH signaling pathway.
The Canonical NOTCH Signaling Pathway
NOTCH precursors are generated in the endoplasmic reticulum (ER) and then transported to the Golgi apparatus after being translated and transcriptionally activated. 23 The EGF-like repeat region of the ER serves as the initial region where the NOTCH precursors are glycosylated. Further, the enzymes POFUT1, POGLUT1, and EOGT1, respectively, catalyze the glycosylation of O-fucosylation, O-glycosylation, and O-GlcNAcylation. 24
Then, in the Golgi apparatus, the xylosyl transferases GXYLT1/2 and XXYLT1 extend O-glucose, whereas the Fringe family of GlcNAc transferases extend O-fucose. 25,26 The Golgi apparatus performs S1 cleavage on the glycosylated NOTCH precursors before delivering them to the cell membrane. A furin-like protease catalyzes the cleavage, which turns NOTCH into a heterodimer at a conserved site (the heterodimerization domain). 27
The heterodimerization domain of the mature NOTCH receptors found on the cell membrane is cleaved in the Golgi apparatus (S1 cleavage). For instance, ligands can start endocytosis by attaching to NOTCH receptor's extracellular domains. When receptors undergo such endocytosis, their conformation changes, revealing the enzymatic site for S2 cleavage. 28,29
Receptors then undergo S3 cleavage, transforming into the effector form: NICD. To control the transcription of targeted genes, NICD is either destroyed in the cytoplasm or trafficked into the nucleus. Since S2 cleavage is the initial step that binds the ligand, it is required for signal initiation. The LNR domain conceals the S2 site (metalloprotease site) in the quiescent phase, also known as the “autoinhibited conformation.” 16 When the receptor is ligand-bound, it lengthens the LNR domain, making the S2 site a prominent one for cleavage. Disintegrin and metalloprotease 10 (ADAM 10), as well as its isoforms ADAM 17 and ADAMTS1, are the principal enzymes for S2 cleavage. 30
The transmembrane domain as well as the intracellular domain constitutes the significant portion of the S2 cleavage products, commonly known as the “NOTCH extracellular truncation” (NEXT). Subsequently, cleavage of NEXT occurs at the S3 region, generating NICD, a transcription factor that may be translocated into the nucleus. 31 The catalytic subunits presenilin1 or presenilin2 (PS1 or PS2), APH-1, PEN-2, and nicastrin (NCT) are found in the enzyme secretase, which is responsible for S3 cleavage.
In addition, when NEXT is endocytosed, S3 cleavage can take place either in the endosome or on the cell membrane, referred to as the endocytic activation model and the endocytosis-independent model, respectively. 32 NICD is translocated into the nucleus to control gene transcription after being released from the cell membrane; the mechanism behind this action might be associated to the NICD's nuclear localization sequences. 33 Figure 3 illustrated the schematic presentation of the Canonical NOTCH pathway.
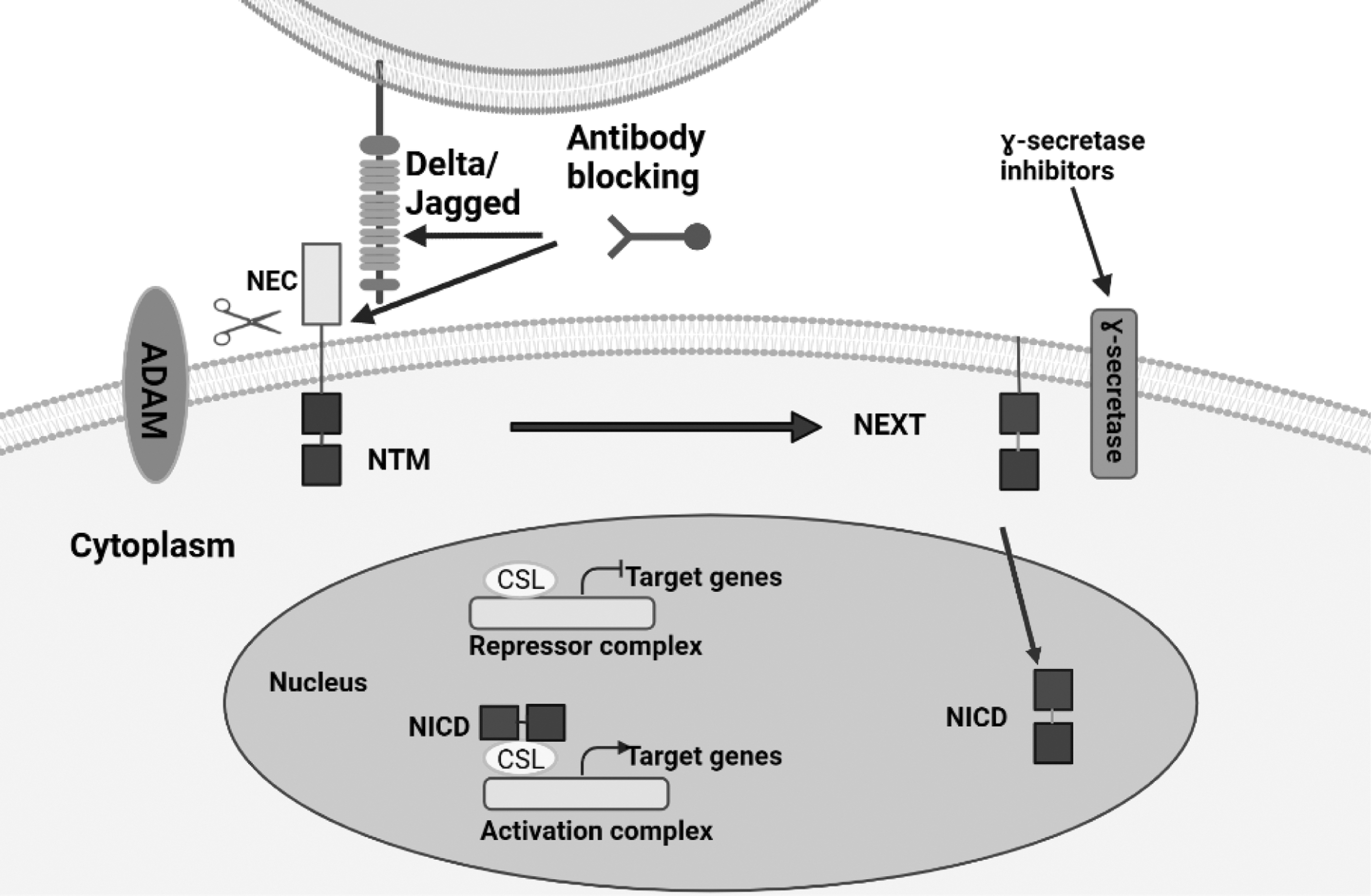
Schematic illustration of the Canonical NOTCH pathway. NOTCH signaling is activated when a ligand binds to a nearby NOTCH receptor between two adjacent cells. The metalloprotease and γ-secretase complex cleaves the NOTCH receptors in a sequence of proteolytic reactions once they have been activated. The NICD is dispersed by the cleavage into the cytoplasm, where it might go on to the nucleus. A repressor complex regulated by the CSL prevents the transcription of NOTCH target genes in the absence of NICD. The CSL complex is transformed from a transcriptional repressor into a transcription activator complex by NICD when it is inside the nucleus. This activation of NOTCH target genes results from the displacement of the histone deacetylase-corepressor complex and the recruitment of the protein MAML1 and histone acetyltransferases to the CSL complex. Theoretically, NOTCH signaling could be suppressed by preventing interactions between (i) NOTCH and its ligands, (ii) blocking receptor activation, (iii) promoting NOTCH ubiquitination and degradation, (iv) inhibiting NOTCH from translocating to the nuclear compartment, and (v) suppressing the formation of the NICD nuclear complex.
Functions of NOTCH in Tumorigenesis
The NOTCH signaling pathway is critical for angiogenesis and tumor growth in pathological settings, and both ligands and receptors have been discovered to be overexpressed and activated in a variety of tumors, including the tumor stroma. 34 The NOTCH family is also crucial in controlling the interaction between tumor cells and surrounding cells in the tumor stroma.
Such crosstalk entails cis-signaling, where the same cell co-expresses a ligand and receptor, as well as reciprocal trans-signaling connections between tumor cells and surrounding cells. As a result, the NOTCH signaling pathway can control more intricate and diverse interactions between tumor cells and their stromal neighbors as well as the opposite.
It has been demonstrated that these interactions regulate a number of tumor pathogenesis-related processes, including angiogenesis, cancer stem cell (CSC) maintenance, and resistance to cancer treatment. 35
The function and sprouting of nearby tumor endothelial cells may be impacted by NOTCH ligands or receptors produced in cancer cells. 36 In addition, NOTCH ligands or receptors that are expressed in cancer cells function as a crucial signaling mechanism to sustain CSCs in a number of malignancies, which can increase cancer stemness, metastasis, heterogeneity, and resistance against conventional cancer therapy. 37
Depending on the kind of cancer, the NOTCH signaling pathway can have either oncogenic or tumor-suppressive activities due to the intricacy of crosstalk mediated by NOTCH signaling in tumors and their neighboring cells.
NOTCH as an oncogene
Acute lymphoblastic leukemia (ALL) of the human T cell line first established the oncogenic nature of abnormal NOTCH signaling. A further investigation found that activating NOTCH1 mutations were present in 56% of T-ALL patients, proving that NOTCH signaling is definitely oncogenic in the development of human T-ALL. 38 After T-ALL, it was identified that the NOTCH signaling pathway had an oncogenic function in a number of solid tumors, including breast, lung, colorectal, and pancreatic cancer. 39 Additional evidence from mice research demonstrated the functioning of the oncogenic NOTCH pathway in solid tumors.
The investigation further proved that the fusion of the mouse mammary tumor virus into NOTCH4 resulted in the development of a condensed dominant active version of NOTCH4 that produces mammary tumours. 40 Enhanced NOTCH activity has been associated with overexpression of ligands and triggering alterations in solid tumors. The ultimate result of this dysregulation, and the main mechanism through which NOTCH signaling appears to promote cancer, is the acceleration of cell cycle progression and the suppression of apoptosis via overexpression of the target gene. 5
NOTCH as tumor suppressor gene
Research on mouse and human keratinocytes supported the hypothesis that the NOTCH pathway serves as a tumor suppressor. 41 Recent data suggested that NOTCH signaling acts as a tumor suppressor in several human organs, including neuroendocrine tumors, skin cancer, and liver cancer. 42,43 Although the precise mechanism by which NOTCH exerts its tumor suppressor activity is still poorly understood, two potential roles have been noted: (1) modulating cell growth through the expression of transcriptional repressor target genes (HES1 and HEY1), and (2) interacting with other signaling pathways. 44
Moreover, it was discovered that head and neck squamous cell cancer (HNSCC) had NOTCH-inactivating mutations and were found in two distinct cohorts of HNSCC patients, according to two independent whole-genome sequencing studies. A more recent and thorough genomic analysis of HNSCC that discovered NOTCH mutations in about 20% of the patients further corroborated these findings. 45,46
Crosstalk with other oncogenic signaling pathways is about how NOTCH activates its tumor suppressor function. 47 Figure 4 demonstrates the mechanism via which the NOTCH pathway crosstalks with other oncogenic signaling pathways through which NOTCH acts as a tumor suppressor gene. Given that Hh and Wnt are recognized BCC mediators, NOTCH inhibition of Hh and/or Wnt signaling in skin might be a technique for cell proliferation suppression. 48
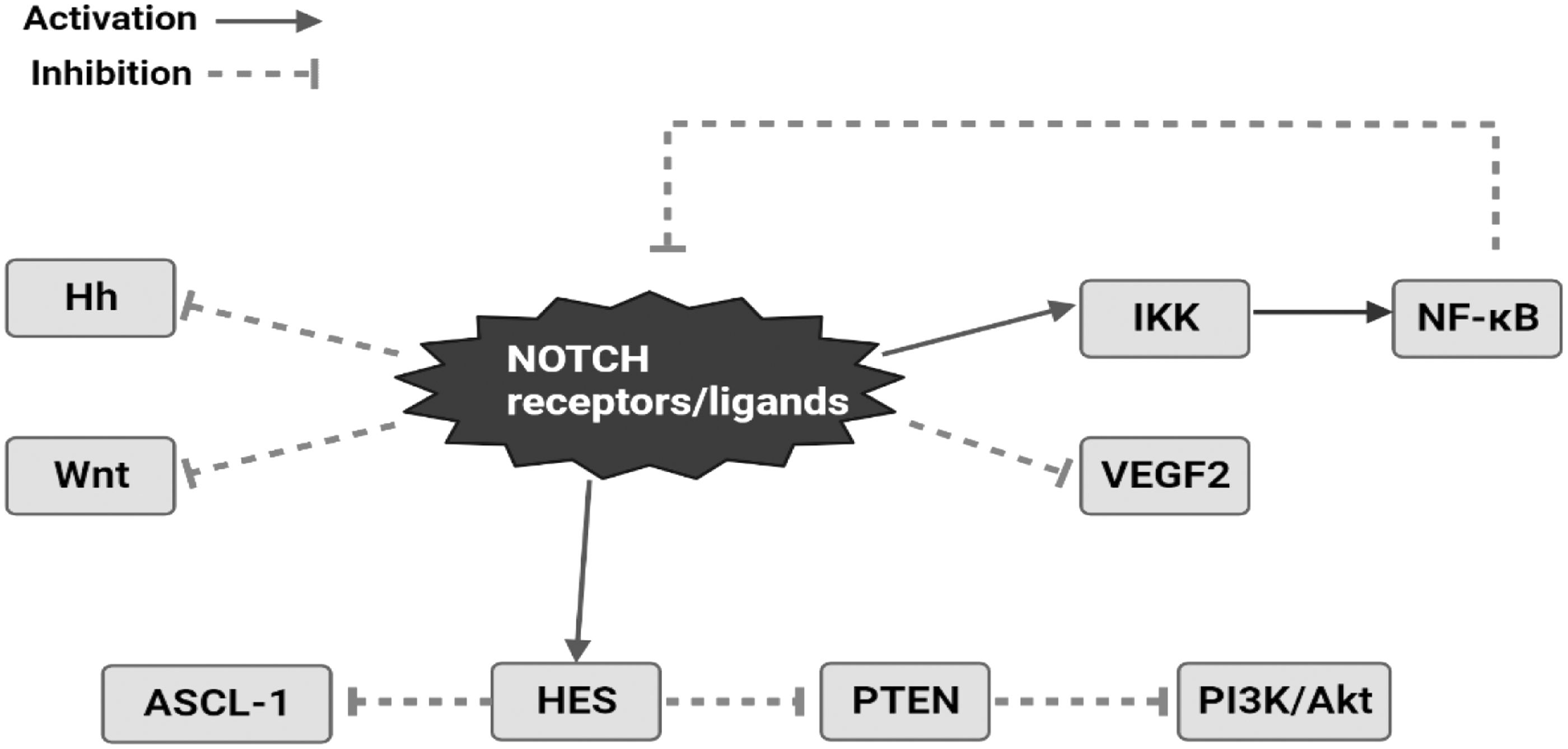
Crosstalk between NOTCH and other oncogenic pathways for tumor suppressor activity of the NOTCH signaling pathway.
By regulating a transcriptional network that controls PTEN expression and PI3K/Akt signaling activity in healthy thymocytes and leukemic T cells, the NOTCH pathway has also been linked to the regulation of the PI3K/Akt signaling Pathway. 49 Likewise, vascular endothelial growth factor (VEGF) regulates angiogenesis by raising the amount of delta-like 4 to promote vessel development. Vascular tumors were produced when this crosstalk was disrupted using delta-like 4-specific antibodies, revealing a tumor suppressor function for NOTCH. 50
Crosstalk between NOTCH and other conserved stem cell pathways
Crosstalk between NOTCH and Hedgehog pathways
Two significant signaling pathways, the NOTCH and Hedgehog pathways, are essential for several cellular and developmental events. Crosstalk between these two pathways has been established in a number of events, such as cancer progression, stem cell maintenance, and embryonic development. In general, the Hedgehog pathway promotes cell proliferation and inhibits cell differentiation, 51 whereas the NOTCH pathway encourages cell differentiation and inhibits cell proliferation. 52
Therefore, the determination of cell fate and tissue homeostasis may be significantly impacted by the interaction between these two pathways. Through the regulation of the expression of each other's components, the NOTCH and Hedgehog pathways interact. For instance, under some circumstances, Hedgehog pathway activation can cause the production of NOTCH ligands, increasing NOTCH signaling. On the contrary, NOTCH pathway activation can cause the production of Hedgehog ligands, increasing Hedgehog signaling. The control of downstream effectors is another method of crosstalk. 53
For instance, NOTCH signaling occasionally can trigger the production of Gli1, a gene that belongs to the Hedgehog pathway and promotes cell proliferation. The NOTCH pathway's target genes, such as Hes1 and Hey1, can also be activated by Hedgehog signaling to promote cell differentiation. 54
Research has shown that the Hh and NOTCH pathways interact, promoting tumor growth in a synergistic manner. First and foremost, it is well known that NOTCH signaling can alter Hh. Second, Hh signaling enables either direct or indirect regulation of NOTCH through subsequent actions. The following are examples of Hh's modification of the NOTCH pathway: (1) The transcription of ligands and regulators in the NOTCH pathway, such as Fringe proteins, is controlled by Hh's downstream effectors. (2) Gli proteins provide immediate, NOTCH-independent transcriptional control of several NOTCH target genes. 55
NOTCH signaling affects Hh in the following ways: (1) Hh components (such as Ptch and Smo) are transported to PC under the regulation of downstream effectors of NOTCH. (2) NOTCH downstream effectors control Gli levels independently of transcription. (3) NICD directly regulates the transcription of the Gli gene. 55,56
In addition, NUMB blocks NOTCH and Hh signaling and aids in the ubiquitination and degradation of NOTCH1 by directly interacting with the WW structure domain of Itchy E3 Ubiquitin Ligase (ITCH) through its polypyrimidine tract-binding protein (PTB) structural domain. 57 The Hedgehog pathway is also directly inhibited by NUMB, whereas ITCH stimulates Glis1 to be ubiquitinated. 58
Stem cell maintenance and tissue homeostasis depend on the interplay of the NOTCH and Hedgehog pathways. For instance, in the skin, NOTCH signaling is essential for the maintenance and differentiation of epidermal stem cells into various skin cell types, but Hedgehog signaling is required for the activation of these cells. 55 Similar to this, NOTCH signaling facilitates the preservation of hair follicle stem cells in the hair follicle whereas Hedgehog signaling is required for their activation and proliferation.
The interaction between the NOTCH and Hedgehog pathways is intricate and context-specific overall. Cell fate determination and tissue homeostasis can be significantly impacted by the interaction between these two pathways, and disruption of this crosstalk has been linked to a number of diseases, including cancer. To completely comprehend the processes of crosstalk and its physiological and pathological effects, further research needs to be done.
Crosstalk between NOTCH and WNT signaling pathways
It has been determined that the NOTCH and WNT signaling pathways interact in a variety of developmental processes, including cancer. Through the direct or indirect regulation of one another's vital components, these crosstalks can result in either feedforward or feedback loops. It has been demonstrated that NOTCH signaling suppresses Wnt signaling in skin cancer, acting as a tumor suppressor.
Similar to this, NOTCH1 signaling continued to limit the expression of Wnt target genes in colorectal cancer even after B-Catenin degradation by the APC complex was impaired. In addition, NOTCH may bind to B-catenin and influence its stability; as a result, the loss of NOTCH1 activity results in the activation of B-catenin. 59 In addition, NOTCH1 target gene NOTCH-regulated ankyrin repeat protein 1 (NRARP) and WNT target genes have been found to be negatively correlated in human colorectal cancer. 60
However, by stabilizing the transcriptional factor LEF1 and hence raising the LEF1 dependent promoter activity, NARP may function as a positive regulator of Wnt signaling. Recently, the function of this crosstalk in initiating the early stages of myeloid regeneration and in myeloid cancers has been thoroughly characterized. 59 On the other hand, Wnt signaling has the ability to directly control the expression of many NOTCH signaling elements, including Delta-like1 (Dll1), Hes1, NOTCH2, Jag1, etc. Direct interactions between B-catenin and NOTCH1 can lead to NOTCH1's decreased ubiquitination, which, in turn, affects NOTCH1's stability and function. 61
The nuclear localization, stability, and transcriptional activity of NICD-1 are similarly impacted by GSK3B's direct interaction and phosphorylation of its serine and threonine residues. Further, NICD-2 may also be phosphorylated by GSK3B; however, this has the effect of decreasing transcriptional activity. The promoter activity of a NOTCH-sensitive reporter construct shows that Dvl, another element of Wnt signaling, physically interacts with RBPJ to lower its transcriptional activity. 54
Targeted therapies by inhibition of NOTCH signaling pathway for cancer treatment
Studies on the impact of inhibiting NOTCH signaling on various cancer types employ a variety of methodologies. Cancer incidence and death in individuals might be significantly decreased by preventing the progression of the cancer with natural or synthetic compounds.
γ-Secretase inhibitors
The ground-breaking research done to target the γ-secretase (GS) complex in Alzheimer's disease led to the development of the first drug used to target NOTCH in the clinical field. GSIs opened the way to preclinical and early clinical research of NOTCH signaling pathway inhibition in cancer, despite the fact that they have not yet been successful in treating Alzheimer's disease. 62
An intriguing multi-subunit protein complex called γ-secretase has a catalytic core that is found in the intramembrane space and is frequently triggered by foreign factors. This catalytic center facilitates the cleavage of single-pass transmembrane receptors. Presenilin (PSEN1 and PSEN2), a catalytic component, interacts with Nicastrin, anterior pharynx defective 1 (APH1), and presenilin enhancer 2 (PEN2) to form the γ-secretase complex. 63
The Delta, Jagged, and Lag2 family of cell surface proteins are among the canonical and noncanonical ligands that serve as receptors for NOTCH. NOTCH isoforms are reliant on ADAM metalloproteases and GS for cleavage after ligand binding, which further enables the NICD to translocate to the nucleus, where it along with CSL and coactivators of the MAML family generates a transcriptional activation complex. 64,65 Clinical GSIs each have a unique profile of NOTCH inhibition and are pharmacologically distinctive. Further, cleavage of certain NOTCH receptors can be induced by low GSIs. 66
A potent and broad-spectrum GSI, MK-0752 (cis-3-[4-[(4-chlorophenyl)sulfonyl]-4-(2,5-difluorophenyl) cyclohexyl]propanoic acid), inhibits NICD cleavage and its associated nuclear translocation was developed initially to treat Alzheimer's disease but was later repurposed to treat T-ALL and in a limited number of individuals, it showed modest effectiveness in preventing the growth of T-ALL and reverting back the condition. 67,68
In Alzheimer's disease, LY411,575, a GSI that binds to presenilin 1 (PS1), has been successfully utilized to diminish amyloid-peptide accumulation. LY 411,575 therapy enhanced apoptosis and re-sensitized resistant HER2+ cells to trastuzumab in the HER2+ breast cancer cell line BT474. 69
GSIs are frequently used with other treatments. Dong Wang and his colleagues, to inhibit breast cancer development, combined the γ-secretase inhibitors MK-0752 and RO4929097 with interleukin (IL)-6. They discovered that breast CSCs have higher NOTCH3 expression and subsequently, by reducing IL-6 production in breast CSCs expressing NOTCH3, γ-secretase inhibitors were utilized in conjunction with IL-6 antagonists to suppress tumor formation. 70
In another study, based on NOTCH1 and mitochondrial co-localization in TNBC cells, Hossain et al. combined the protein kinase B (AKT) inhibitor MK-2206 with a γ-secretase inhibitor (PF-03084014) and to inhibit breast cancer cell multiplication, they suppressed NOTCH pathway activation. 71
In addition to their direct effects on tumor cells, the majority of GSIs have indications of anti-angiogenic actions in vivo. Probably, the suppression of the crucial NOTCH-VEGF crosstalk for angiogenesis is to blame. 39,72 GSIs are expected to cause several off-target effects in vivo because of the diverse range of substrates that γ-secretase might bind to. However, their toxicity appears to be virtually entirely NOTCH-mediated. 73
The most severe side effect associated with GSIs is diarrhea (dose-limited) that is brought on by goblet cell metaplasia of the small intestine, caused by NOTCH suppression in intestinal epithelial stem cells. 74 A list of GSIs is tabulated in Table 1, which displays various GSIs and its utilization for cancer treatment along with its clinical status.
Lists of Gamma-Secretase Inhibitors for Several Cancers in Clinical Trials
Monoclonal antibodies
One of the effective ways to target the NOTCH signaling pathway is with monoclonal antibodies directed against specific receptor regions that could also distinguish between different NOTCH receptors and ligands. Further, the monoclonal antibody (mAb) reduced cell growth and brought on apoptotic cell death. Also, exposure to this antibody diminished the MDA-MB-231 cell's CD44Hi/CD24. A low subpopulation implied a decline in the cancer stem-like cell subpopulation. 75 Numerous investigations demonstrated the potential of monoclonal antibodies to inhibit NOTCH signaling and significantly diminish CSC burden.
Silva et al. demonstrated the potential of innovative monoclonal antibodies against NOTCH signaling ligand DLL1. They discovered that the Dl1.72 (an anti-DLL1 antibody) significantly reduced the expression of NOTCH target genes and DLL1-NOTCH signaling that expressed the estrogen receptor-positive in breast cancer cells. In vitro exposure with Dl1.72 decreased MCF-7 cell motility, proliferation, and the ability to generate mammospheres and endothelial tubes.
In vivo, Dl1.72 dramatically reduced liver metastases and tumor cell proliferation in a xenograft mouse model without exhibiting any obvious hazard. These results imply that anti-DLL1 Dl1.72 would be a desirable treatment for estrogen receptor-positive in breast cancer cells and would urge further preclinical research. 76 Liu et al. explored the role of IL-6 in the drug-resistant multiple myeloma (MM) cell line's chemosensitivity to bortezomib.
The result showed that anti-IL-6 and BTZ have the ability to halt the cell cycle, incite apoptosis, and decrease the proliferation of KM3/BTZ cells. The coupled effect of anti-IL-6 and BTZ was more apparent on KM3/BTZ cells than with either drug alone, and it vastly reduced the expression of IL-6, NOTCH1, and STAT3 in KM3/BTZ cells. They were able to conclude that IL-6 can affect MM cell's chemosensitivity to BTZ and that IL-6 might lessen this sensitivity through the STAT3/NOTCH signaling pathway. 77
Brontictuzumab (OMP-52M51) is a humanized mAb that selectively targets NOTCH1 juxta membrane NRR and thus inhibits NOTCH signaling. In a phase II trial of brontictuzumab, six of the total 36 (17%) patients with refractory solid tumors demonstrated clinical benefits, with four patients displaying prolonged disease stabilization (NCT01778439). 78
Hu et al. conducted a phase II trial on a fully human IgG2 antibody called tarextumab that blocks NOTCH2/3 receptors and has demonstrated promising anticancer effectiveness in small cell lung cancer (NCT01277146). Patients receiving tarextumab along with etoposide and platinum-based treatments experienced an overall response rate of 84% in the phase I/II trial. In addition, tarextumab-induced diarrhea was dose-limiting above 2.5 mg weekly and 7.5 mg/kg every 3rd week, and tarextumab causes significant inhibition of NOTCH signaling.
When tarextumab was combined with gemcitabine and nab-paclitaxel, tumor development and the frequency of tumor-initiating cells were more inhibited vis a vis tarextumab has coupled alone with gemcitabine. 79
Natural compounds
Diallyl trisulfide
Diallyl trisulfide (DATS, allitridin), a bioactive component of Allium sativum commonly called garlic, has been shown to have potent anticancer and chemo-preventive effects. Chemically, it is an organosulfur molecule that is synthesized by the hydrolysis of allicin. DATS have been proven to regulate tumorigenesis, angiogenesis, and metastasis in a variety of cancer types. 80,81 DATS was utilized to exhibit suppression of osteosarcoma (OS) cell growth by inducing cell cycle arrest as well as apoptosis in vitro.
Through reduced production of NOTCH-1 downstream genes such as VEGF and matrix metalloproteinases, DATS has also been shown to inhibit cell survival, wound-healing capacity, invasion, and angiogenesis in OS cells. 82 Kiesel and Stan had demonstrated in their studies that diallyl trisulfide targets breast cancer that overexpresses NOTCH pathway components and may perform as an important nutraceutical for preventing breast cancer. Their studies showed that treatment with 40 mM DATS decreased Jagged-1 expression in MDA-MB-231 and MCF7 cells by around 40% and 60%, respectively, for 24 h. 83
Genistein
Genistein (Gen), one of the major soy isoflavones in soy products, is consumed regularly by Asian populations, and it was reported as a protective factor against breast cancer. This phytoestrogen may be one of the leading compounds as its safe and anticancer activities have already been tested in several in vitro and preclinical models. 84 NF-κB activity and the NOTCH-1 signaling pathway were examined by Pan et al. in relation to the effects of genistein.
The findings showed that Gen may reduce the proliferation of TNBC by a new mechanism that involves inhibiting NF-κB activity via the NOTCH-1 pathway. By reducing NF-κB activity via the Nocth-1 signaling pathway in a dose-dependent manner, Gen reduced the development of MDA-MB-231 TNBC cell lines. Exposure of MDA-MB-231 cells with 0, 5, 10, or 20 μM Gen resulted in apoptotic rates of 6.78%, 18.98%, 30.45%, and 60.64%, respectively. 85
Curcumin
The phenolic compound curcumin, which is derived from the plant Curcuma longa (Linn), has exhibited antitumor efficacy in a variety of cancers. 86 Curcumin's potential to target a myriad of cancer-related signaling pathways has led to extensive research on its anti-proliferative effects against a variety of cancer forms. 87 Curcumin also targets the NOTCH-1 signaling pathway. Curcumin has been observed to impair NOTCH-1 signaling inside the NICD in the HEP3B, SK-Hep-1, and SNU449 cell lines. 88,89
A potential therapeutic target for hepatocellular cancer is the regulation of the NOTCH1 signaling system by MSCs and/or NCD. 90 Increases in curcumin dosage resulted in a reduction in NOTCH1 mRNA and protein expression. As a result, curcumin could modify the NOTCH signaling pathway to prevent proliferation and trigger apoptosis in hepatoma cells. 91
Liu et al. demonstrated that increased inhibition of hepatoma cells, that is, SMMC-7721 in culture, was observed in response to increasing dosages of curcumin, and these changes were associated with a reduction in NOTCH-1 mRNA as well as protein expression. 92 Likewise, Abdel Aziz et al. and his colleagues showed in his studies that curcumin triggered the apoptosis of hepatoma cells by suppressing NOTCH-1 as well as its target genes CyclinD1 and HES1. 93
The photosensitizing ability of curcumin has been proven in the activation of apoptosis, suppression of cell growth, and modulation of NOTCH signaling pertaining to cervical cancer. A study conducted by He et al. in the cervical cancer cell line Me180 showed that the curcumin along with DAPT, a NOTCH receptor blocker, exhibited an increase in apoptosis rate and a notable suppression of NOTCH1 mRNA expression of 79.27%. 94
Resveratrol
Resveratrol, a dietary polyphenolic phytoalexin found in grape and peanut peels, is a widely researched phytochemical with extraordinary pharmacological properties and has the ability to change a variety of pathways in different malignancies. 95 Numerous studies demonstrate that resveratrol being a double-edged sword suppresses NOTCH signaling because it exhibits both oncogenic and tumor-suppressive effects on tumor cells. 96
Resveratrol has been observed to primarily activate NOTCH1 to trigger differentiation-associated genes in anaplastic thyroid cancer (ATC). 97 In medullary thyroid carcinoma (MTC), resveratrol elevated NOTCH-2 mRNA, caused apoptosis, and decreased the neuroendocrine marker ASCL-1. Meanwhile, it enhanced NOTCH-1 signaling in ATC and inhibited the development of ATC cells both in vitro and in vivo. Together, these findings suggest that NOTCH inhibits tumor growth in MTC as well as ATC. 98
Nanotechnological Approaches for NOTCH Pathway Inhibition
Nanoparticles (NPs) have tremendous potential as carriers for enhancing the delivery of small molecules and antibodies to tumor cells to regulate NOTCH signaling and interrupt disease development (Table 2). Encapsulating these molecules inside NPs or mounting them on the surface of NPs can increase their pharmacokinetics, stability, biodistribution, and site-specific delivery. This allows the nano-formulations to be far more effective than their freely supplied counterparts. 99
Lists of Nano-Formulations for Anti-NOTCH Directed Therapies
CSC, cancer stem cell; TNBC, triple-negative breast cancer.
GSIs, which impede NICD cleavage and restrict its translocation to the nucleus, are the most thoroughly researched class of NOTCH signaling inhibitors. 9 To present, neither ADAM inhibitors nor MAML1 inhibitors have been integrated into NP formulations, although GSIs and monoclonal antibodies have begun to be investigated in nano-formulations. 100,101
GSIs are effective against NOTCH-dependent tumors; they frequently cause significant gastrointestinal adverse effects when administered orally due to their limited water solubility. Encapsulating GSIs into NPs is one effective way to reduce these toxic off-target effects. 100,102 Mamaeva et al. developed mesoporous silica NPs to deliver the GSI, DAPT, to breast cancer patients in intriguing research.
They discovered that NP-mediated administration of DAPT efficiently blocked NOTCH signaling to diminish the CSC population in MDA-MB-231 breast cancer cells implanted on chorioallantoic membranes. 103 Likewise, Chandra et al., formulated α-mangostin encapsulated PLGA NPs (Mang-NPs) and investigated the molecular pathways by which Mang-NPs suppress colorectal cancer cell viability, colony formation, epithelial-mesenchymal transition, and cause apoptosis.
They observed that the Mang-NPs suppressed NOTCH signaling by inhibiting the expression of NOTCH receptors (NOTCH1 and NOTCH2), their ligands (Jagged 1 and DLL4), the γ-secretase complex protein (Nicastrin), and downstream targets (Hes-1). 104 Lu and colleagues used polylactic acid-based NPs to administer DAPT in conjunction with the EGFR inhibitor erlotinib to TNBC. When the NPs were coated with both CREKA, a tumor-homing peptide, and F3, a cell-penetrating peptide, they achieved stronger antitumor effects in vivo than NPs coated with either external functional agent alone or no coating. 105
GSIs are pan-NOTCH inhibitors that target NOTCH 1–4 altogether, which can increase toxicity when compared with targeting the receptors separately. Further, because particular receptors serve diverse roles in the development of various types of cancer, inhibiting these receptors individually might be beneficial. 106 Rather than seeking to inhibit downstream pathway mediators, RNA interference (RNAi) directed at NOTCH signaling specifically affects the NOTCH receptors and their ligands. 100
Deng et al., used chitosan-based NPs to deliver the tumor inhibitory microRNA miR-34a to TNBC. They revealed that delivering miR-34a to MDA-MB-231 TNBC cells through these NPs hindered cell migration and decreased NOTCH-1 protein expression. 107 Similarly, Yang and his colleagues synthesized iron oxide-silica NPs that efficiently deliver shRNA targeting NOTCH-1 to TNBC. They showed that these NPs significantly inhibit cell proliferation and NOTCH-1 expression while inducing apoptosis in MDA-MB-231 TNBC cells. 108
Trends in cancer models system in the nano-medicine field
There is ongoing research and development in the field of nanomedicine, and the choice of model systems used can vary depending on the specific research question, the type of nanomedicine being tested, and the available resources. However, there has been a growing interest in the use of translational relevant disease models and assays in nanomedicine research, particularly in recent years.
The use of model systems in nanotechnology research is crucial for the development of new therapies and technologies. Traditional model systems include cell cultures and animal models, such as patient-derived xenografts (PDX), which involve the transplantation of human tumors into immunocompromised mice. PDX have been widely used as preclinical models for testing nanomedicines, as they allow for the evaluation of efficacy and toxicity in a tumor environment that more closely mimics human disease.
However, there are limitations to the use of PDX models, such as the variability in tumor growth rate and response to treatment between individual patient-derived tumors, as well as the ethical and logistical considerations involved in using animal models. 109
However, in recent years, there has been a growing focus on the development of alternative model systems that are more representative of human disease and can reduce the reliance on animal models. One such approach is the use of 3R (Replacement, Reduction, Refinement) principles, which aim at replacing animal models with non-animal alternatives, reducing the number of animals used in experiments, and refining experimental protocols to minimize pain and distress to animals.
The three Rs, which Russell and Burch (1959) proposed, were techniques to make animal studies safer. This strategy encourages the utilization of the fewest possible animals, or a “reduction,” in the overall number of animals used during an experiment. It is crucial to carefully organize and “refine” the usage of animals so that the amount of suffering they experience throughout the experiment is kept to a minimum.
Further, smaller organisms and alternate methods should “replace” higher animals wherever possible. 110 Numerous strategies have been put up to prevent the use of animals in testing. Up to a point, these techniques provide an alternate way to conduct chemical and drug testing. These approaches have the advantages of being time-efficient, using fewer manpower, and being affordable. These methods are: computer models, cells and tissue cultures, and alternative organisms.
To predict the numerous biological and toxic effects of a chemical therapeutic candidate without involving the dissection of animals, specialized computer models and software programs are utilized. For instance, in vivo research is required to determine a drug's receptor binding location.
The receptor binding site for a prospective therapeutic molecule is predicted using software called Computer Aided therapeutic Design (CADD). By locating a likely binding site, CADD prevents testing of undesired compounds with little biological activity. In addition, with the use of such software tools, we may custom-make a new medicine for a particular binding site. Finally, animal testing is carried out to get conclusive results.
Consequently, fewer experimental animals are used overall, and the 3 Rs' goals are satisfied. Likewise, Structure Activity Relationship computer programs are another well-liked resource. On the basis of the existence of chemical moieties connected to the parent substance, it predicts the biological activity of a therapeutic candidate. In vitro cell and tissue cultures, which include growing cells outside the body in a lab setting, can be a significant replacement for animal testing.
The organs' cells and tissues—liver, kidney, brain, skin, etc.—are taken from the animal and stored outside the body in a proper growth medium for a short while, a few weeks, or even a few years. A monolayer of isolated animal or human cells is grown on the surface of culture plates or flasks during in vitro cultivation. 110
Other translational relevant disease models' assays are also gaining acceptance in the nanotechnology field. These models use patient-derived cells or tissues to better represent human disease and can provide more accurate information about drug efficacy and toxicity.
Organ-on-a-chip models
A device called an “organ-on-a-chip” combines microfabrication with tissue engineering to mimic the vital physiological conditions and functions of human organs. These models can be used to study the effects of nanomaterials on specific organs, such as the liver, lung, or heart, in a more realistic and controlled manner than traditional animal models. As a result, it may be utilized to forecast drugs responses and the impact of the environment on organs.
High-precision microfluidic technology can regulate reagents at the microscale. Microfluidics has, therefore, been extensively used in organ-on-chip devices to simulate a single organ or a number of organs in vivo. These sensor-integrated models have a lot of promise for replicating the human environment. 111
Three-dimensional cell culture models
Three-dimensional cell culture models involve growing cells in a three-dimensional environment that more closely mimics the structure and function of tissues in the body. These models can be used to study the interactions between nanomaterials and cells in a more physiologically relevant context than traditional 2D cell cultures. Due to its obvious advantages in giving more physiologically appropriate information and more predictive data for in vivo studies, three-dimensional (3D) cell culture techniques have attracted growing interest in drug development and tissue engineering. 112
Three-dimensional cell culture platforms, which simulate in vivo microenvironmental elements and are therefore anticipated to provide outcomes with greater predictive value for clinical outcome, have recently gained prominence in the drug development process. In addition, human cells used in realistic 3D cell culture models can avoid the limitations of mice models, which, in addition to being expensive and unethical, are occasionally unable to effectively mimic human disease or capture adverse effects of drugs such liver damage. 113
Conclusion and Future Prospects
Since the discovery of the NOTCH signaling pathway almost 100 years ago in drosophila, extensive efforts have been made from then and its relationship with various inherited diseases including cancer has been established. Deregulation of NOTCH proteins has been linked to the initiation and spread of cancer. These aspects of NOTCH signaling make it a tempting prospect for cancer treatment as well as a potential diagnostic and prognostic biomarker.
A group of NOTCH pathway inhibitors have been developed, utilizing the molecular structure of the NOTCH receptor and NOTCH ligands. Therefore, targeting NOTCH signaling pathway provides a better insight for the management of several kinds of cancer. Repurposing of gamma secretase inhibitors (which was initially used for Alzheimer's diseases) and monoclonal antibodies is currently the most exploited NOTCH-targeted therapies for cancer treatment.
Moreover, the discovery of natural compounds with inhibitory effects on the NOTCH pathway provides additional hope for cancer management. This opens up new possibilities for the development of novel therapeutic agents with fewer side effects and enhanced efficacy. With ongoing research and innovative therapeutic approaches, we can expect further advancements in utilizing the NOTCH pathway as a potential target for cancer therapy and as a valuable tool for cancer diagnosis and prognosis.
Footnotes
Acknowledgment
Balak Das Kurmi and Preeti Patel would like to thank Shivani Rai Paliwal and Harish Rajak from the Institute of Pharmaceutical Sciences, Guru Ghasidas Vishwavidyalaya (A-Central University), Bilaspur, CG, India, for their guidance.
Authors' Contributions
L.K.: Writing original drafts and data curation; L.M.: Data curation; Yash Sharma: Writing, review, and editing; K.C.: Writing, review, and editing; M.K.: Writing, review, and editing; P.P.: Supervision and review correction; G.D.G.: Project administration; B.D.K.: Conceptualization, Supervision, and Funding acquisition.
Disclosure Statement
The author(s) declared no conflict of interest.
Funding Information
B.D.K. and G.D.G. are thankful to the Indian Council of Medical Research (ICMR), New Delhi, India for providing financial assistance in the form of ICMR - Adhoc Research Project (File No.: 67/05/2022-DDI/BMS; RFC No.: BMS/Adhoc/121/2022–23; IRIS ID: 2021–10997).