
Abstract
Abstract
Reprogramming of differentiated nuclei into a totipotent embryonic state following somatic cell nuclear transfer (SCNT) is not efficient. Previous studies in the hybrid B6D2F1 mouse strain revealed that a transient treatment of the SCNT embryos with the histone deacetylase inhibitor (HDACi) trichostatin A (TSA) significantly enhance the potential of the cloned embryos to develop in vitro and to term. Here, we compare two different SCNT protocols with TSA and explore, for the first time, the effect of another HDACi, valproic acid (VPA), on the in vitro development, blastocyst quality, and full-term development of mouse B6CBAF1 cloned embryos. Rates of blastocyst development in SCNT embryos treated with either 5 nM TSA during and after activation (31.8%) or with 100 nM TSA or 2 mM VPA before and during activation (34.5 and 38.3%, respectively) were clearly superior to those of nontreated SCNT embryos (22.9–25.1%). These increased in vitro development rates of the HDACi-treated embryos were correlated with an increased level of histone H3 lysine 14 acetylation and an improved blastocyst quality, as judged by the increased number of total and ICM cells in comparison to the nontreated embryos (30–35% increase). Treatment of SCNT embryos with TSA or VPA also allowed the obtention of viable cloned mice, whereas none could be produced from untreated SCNT embryos. In conclusion, we have demonstrated for the first time that VPA can improve the in vitro and full-term development of B6CBAF1 SCNT embryos, at a similar level as TSA. Our findings may open new opportunities to improve cloning efficiencies in other mouse strains or species.
Introduction
In an effort to improve cloning efficiency, several modifications to the original mouse cloning method (Wakayama et al., 1998) have been attempted, but none of them has resulted in a remarkable improvement (reviewed in Wakayama, 2007). More recently, however, two independent studies with the hybrid B6D2F1 mouse strain revealed that a transient treatment of the SCNT embryos with the histone deacetylase inhibitor (HDACi) trichostatin A (TSA), significantly enhances in vitro development up to the blastocyst stage and leads to a two- to fivefold increase in cloning efficiency (Kishigami et al., 2006a; Rybouchkin et al., 2006). Further, the TSA treatment led to the first success in the cloning of an adult ICR mouse, an outbred strain that had been considered as “unclonable” (Kishigami et al., 2007). Additional studies also revealed that TSA treatments decrease the expression of chromatin structure- and DNA methylation-related genes (Li et al., 2008) and improve constitutive heterochromatin remodeling in SCNT-derived embryos (Maalouf et al., 2009), all of which might be correlated with the improved rates of embryonic development observed in TSA-treated embryos.
Despite the obvious beneficial effects of TSA on mouse cloning efficiency, the optimal concentration and timing of the TSA treatment are not yet clear, as different groups have used slightly different protocols. In particular, Kishigami et al. (2006a) first reported an improved cloning efficiency when treating the SCNT embryos with 5 or 50 nM TSA during (6 h) and after (4 h) oocyte activation. Although Rybouchkin et al. (2006) obtained a similar result when using 100 nM TSA and applying the treatment before (2–3 h) and during (6 h) oocyte activation.
It has been speculated that induction of pluripotent stem (iPS) cells from somatic cells transfected with defined transcription factors, first demonstrated by Takahashi and Yamanaka in 2006, may share common mechanisms of somatic nuclei reprogramming with SCNT. In recent studies, it was demonstrated that induction of iPS cells by defined factors is greatly improved by small molecules that alter the epigenetic status of the somatic cell nuclei, including histone and DNA methyltransferases inhibitors and HDACis (reviewed in Feng et al., 2009). In particular, the HDACi valproic acid (VPA) has been shown to induce reprogramming of mouse fibroblasts with only three transcription factors, with a considerably higher efficiency than TSA (Huangfu et al., 2008a). Moreover, VPA enabled reprogramming of primary human fibroblasts with only two factors, Oct4 and Sox2, without the need for the oncogenes c-Myc or Klf4 (Huangfu et al., 2008b). The superior beneficial effects of VPA, in comparison to TSA, for the in vitro reprogramming of somatic cell nuclei using transcription factors prompted us to investigate whether a similar trend would be observed in the in vivo reprogramming of somatic cell nuclei during SCNT.
Thus, in the present study, we explore for the first time the effect of VPA on the in vitro and the in vivo development of SCNT mouse embryos and we compare it to the effect of TSA. In the first part of our study, treatment of SCNT embryos with the two HDACis before and during oocyte activation (as reported by Rybouchkin et al., 2006) or during and after activation (as reported by Kishigami et al., 2006a) was tested, and the acetylation status of histone H3 lysine 14 (H3K14) in the transferred nuclei and the in vitro development and quality of the cloned embryos were compared. In the second part of the study, the full-term developmental potential of the SCNT embryos produced with the most efficient HDACis treatments was assessed.
Material and Methods
Unless otherwise indicated, all reagents were purchased from Sigma (Spain).
Animals
Animal care and procedures were conducted according to the protocols approved by the Ethics Committee on Animal and Human Research of the Universitat Autònoma de Barcelona. Hybrid B6CBAF1 (C57BL/6J × CBA/J) female mice were used as oocyte and somatic cell (cumulus cell) donors and for the collection of in vivo fertilized embryos that were used as control in the embryo transfer procedures. Outbred CD1 females mated with normal or vasectomized males of the same strain were used as foster or surrogate mothers, respectively.
Collection of oocytes, embryos, and cumulus cells
Females were induced to superovulate by intraperitoneal injection of 5 IU of pregnant mare serum gonadotropin (PMSG; Intervet, Spain) followed 48 h later by 5 IU of human chorionic gonadotropin (hCG; Farma-Lepori, Spain). Cumulus–oocyte complexes were collected from the oviducts 13–14 h after hCG administration in HEPES-buffered CZB medium (H-CZB) (Chatot et al., 1989), and treated with hyaluronidase (300 U/mL) in H-CZB at 37°C until cumulus cells dispersed. The denuded oocytes were then washed twice in H-CZB and kept in KSOM culture medium (106-D, Millipore, Bedford, MA, USA) containing essential and nonessential amino acids, glucose, and 1 mg/mL of BSA under oil at 37°C in an atmosphere of 5% CO2 in air, until use. Cumulus cells were removed from the hyaluronidase drops and placed in an eppendorf with H-CZB medium to be washed by centrifugation at 250 × g. The cumulus cell pellet was then resuspended in a small volume of 3% polyvinyl pyrrolidone (PVP) in H-CZB and kept on ice until the moment of the nuclear injection.
For the obtention of in vivo fertilized pronuclear stage embryos, superovulated females were mated with B6CBAF1 males after hCG administration and the embryos were collected from the oviducts 20 h later in H-CZB. After treatment with hyaluronidase (300 U/mL) and several washes, denuded zygotes were cultured in KSOM at 37°C with 5% CO2 in air for 24 h before their transfer into recipient females.
Production of cloned embryos by somatic cell nuclear transfer
Nuclear transfer experiments were performed essentially as previously described (Wakayama et al., 1998). All micromanipulations were performed using an Eppendorf micromanipulation system installed on an Olympus IX71 microscope. After 5 min of pretreatment in H-CZB medium supplemented with 5 μg/mL cytochalasin B (CB), meiotic spindles of B6CBAF1 oocytes were removed using a Piezo-driven (Burleigh) pipette with an outer diameter of 9–10 μm. Enucleated oocytes were then washed extensively and returned to the incubator in KSOM medium for 1–2 h before injection. Somatic nuclei were isolated from cumulus cells by pipetting the cells with a 6–7-μm pipette and injected individually into the enucleated B6CBAF1 oocytes in H-CZB medium supplemented with 5 μg/mL of CB. Reconstructed oocytes were then washed extensively in KSOM and returned to the incubator for 1–3 h.
Parthenogenetic activation of oocytes
Oocytes reconstructed by SCNT were parthenogenetically activated 1–3 h after nuclear injection, in parallel to nonmanipulated oocytes. Activation was performed by incubation of the oocytes in 10 mM SrCl2 in Ca2+-free-CZB medium supplemented with 5 μg/mL CB for 6 h. After activation, oocytes showing two visible pronuclei were cultured in KSOM until the two-cell or blastocyst stages.
Treatments with histone deacetylase inhibitors (HDACis)
A stock solution of TSA was prepared in DMSO at a 500 nM concentration (Kishigami et al., 2006b). VPA was dissolved in tri-distillated water as a 0.2 M stock solution, filtered, and stored at room temperature for up to 2 months. The final concentrations of both HDACis were prepared by dilution of the stock solutions in the culture or activation media, depending on the experimental procedure. According to their reported effectiveness and toxicity (de Ruijter et al., 2003), the final concentrations used were on the nanomolar range for TSA (5 nM and 100 nM) and on the millimolar range for VPA (0.5 mM and 2 mM).
Two protocols were used for the treatment of reconstructed oocytes with the HDACis, based on previous experiments performed by others with TSA. In one protocol (based on Kishigami et al., 2006a), SCNT embryos were activated at 1–2 h post-NT in Ca2+-free-CZB medium (with 10 mM SrCl2 and 5 μg/mL CB) supplemented with either 0.5 mM VPA or 5 nM TSA. At 6 h postactivation (p.a.), the embryos were moved to KSOM for another 2 h of HDACi treatment (total HDACi treatment time = 8 h). In the other protocol (based on Rybouchkin et al., 2006), SCNT embryos were treated with 2 mM VPA or 100 nM TSA in KSOM for 2–3 h immediately after nuclear injection. Then, the SCNT embryos were activated for 6 h in the presence of the same concentration of HDACis (total HDACi treatment time = 8–9 h). At the end of each treatment the embryos were extensively washed, and those with visible pronuclei were then cultured in KSOM medium until the two-cell or blastocyst stages. SCNT embryos that were not exposed to HDACi were used as control.
Immunofluorescence analysis of histone acetylation levels in SCNT embryos
All steps were performed at room temperature, unless otherwise mentioned. Control and HDACi-treated SCNT embryos were fixed in 4% paraformaldehyde in H-CZB medium for 30 min at different time points after reconstruction (10 min and 3 h post-SCNT) and activation (3 h and 6 h postactivation). Once fixed, they were permeabilized in PBS containing 0.5% Triton X-100, 3% goat serum, and 0.2% sodium azide. The samples were then washed and stored overnight at 4°C in a phosphate-buffered saline (PBS) blocking solution composed of 1% (w/v) bovine serum albumin (BSA), 0.2% (w/v) powdered milk, 2% (v/v) normal goat serum, 0.1 M glycine, 0.2% (w/v) sodium azide, and 0.01% Triton X-100 (Wickramasinghe and Albertini, 1992). Afterward, they were incubated for 1 h with a 1:500 dilution of a rabbit anti-acetyl-H3K14 primary antibody (Millipore 06-911), washed four times in blocking solution for 10 min each, and incubated for 1 h with Alexa Fluor 594 goat antirabbit IgG (6 μg/mL, Molecular Probes, Eugene, OR). Finally, after several washes, the samples were stained with 10 μg/mL Hoechst 33258 (Molecular Probes) for 10 min and mounted on slides in 50% (v/v) glycerol/PBS containing 25 mg/mL of sodium azide. Samples were examined using an epifluorescence microscope (Olympus Bx60) equipped with specific filters for Hoechst and Texas Red and a 50-W mercury lamp. Digital images of the acetylated H3K14 (Ac-H3K14) and DNA signals were acquired on Genus Software (version 3.0; Olympus, Melville, NY, USA) using the same contrast, brightness, and exposure settings for all embryos. The mean values of the fluorescence intensities were measured on the Ac-H3K14 and DNA channels separately with Image J Software in at least eight SCNT embryos for each time and treatment group. The average profiles represented as the ratio of Ac-H3K14/DNA signals were then used for the comparison between the different groups.
Blastocyst differential staining
A simplified technique, previously described by Thouas et al. (2001), was used for the differential staining and counting of inner cell mass (ICM) and trophectoderm (TE) cells in cloned and parthenogenetically activated blastocysts. Briefly, embryos that reached the blastocyst stage 96 h p.a. were first incubated in 500 μL of solution 1 (H-CZB medium with 1% Triton X-100 and 100 μg/mL propidium iodide) for up to 10–15 sec. They were then immediately transferred into 500 μL of solution 2 (fixative solution of 100% ethanol with 25 μg/mL Hoechst 33258) and stored at 4°C overnight. Fixed and stained blastocysts were then washed in a drop of glycerol and mounted on a glass microscope slide in a 3 μL drop of glycerol and flattened with a coverslip. Cell counting was performed from digital images obtained on an inverted microscope (Olympus IX71) fitted with an ultraviolet lamp and an excitation filter that allows the visualization of the red and blue fluorochromes simultaneously (U-MNV, Olympus).
Embryo transfer
CD-1 females mated with vasectomized males of the same strain were used as recipients. All embryos were transferred at the two-cell stage into the oviducts of 0.5 day postcoitum (dpc) recipients. Between 9 and 23 SCNT embryos were cotransferred with one to three parthenogenetic embryos (10–15% of the total of embryos transferred) (Meng et al., 2008) to each female. As control of the embryo transfer procedure, 8–10 in vivo fertilized embryos were also transferred to other CD-1 recipients. Females were sacrificed at 19.5 dpc and pups were delivered by Cesarean section. The pups and their corresponding placentas were weighted in an analytic scale and the cloned pups were fostered to a CD-1 mother.
Statistical analysis
All experiments were repeated 4 to 12 times on separate days and the results obtained in the replicated experiments were pooled. Data were analyzed by chi-square test or Fisher's exact test to compare embryonic development between the different groups. Data on mean number of cells in blastocysts, fluorescence intensity ratios, and body and placenta weights were analyzed by one-way ANOVA. A probability value of p < 0.05 was considered to be statistically significant.
Experimental design
To determine the effect of VPA on the efficiency of mouse cloning by SCNT and compare it to the effect of TSA, two series of experiments were performed, as summarized in Figure 1. In the first series of experiments, the in vitro developmental potential and the quality of the HDACi-treated SCNT embryos as well as the levels of H3K14 acetylation in the transferred nucleus were investigated. With this aim, mouse oocytes reconstructed with cumulus cell nuclei were treated with different concentrations of one of the two HDACis either before and during oocyte activation or during and after activation. Reconstructed oocytes that were not treated with the HDACis and that were parthenogenetically activated at the same times as the HDACi-treated oocytes were used as controls. Some oocytes were fixed at several time points after reconstruction (10 min and 3 h) or activation (3 and 6 h) and processed for immunofluorescence detection of acetylated H3K14. The remaining SCNT embryos were cultured in vitro until the blastocyst stage and their development was assessed every 24 h. By 96 h p.a. the control and HDACi-treated SCNT blastocysts together with a group of nonreconstructed parthenogenetically activated oocytes were processed for differential staining and counting of ICM and TE cells.
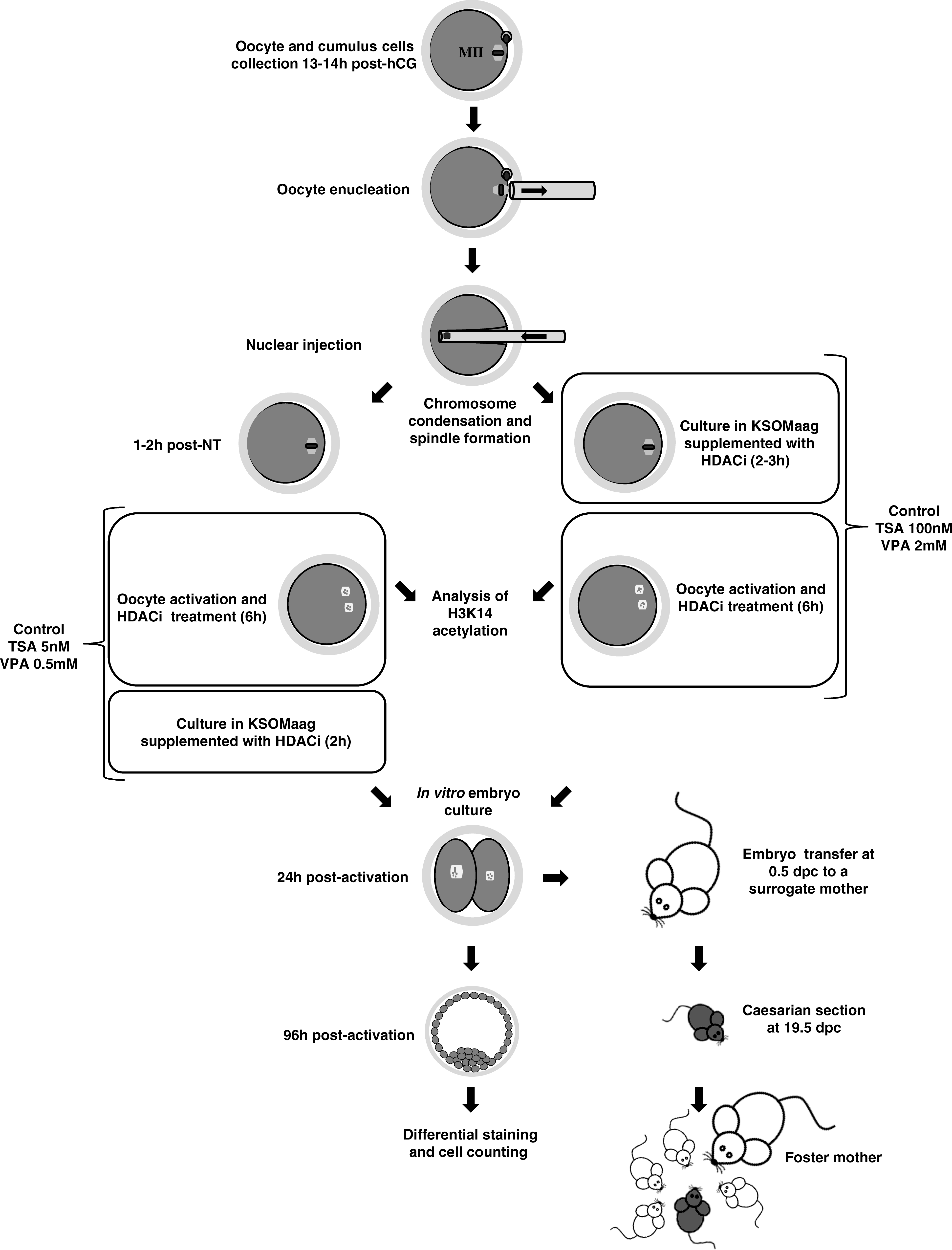
Schematic view of the experiments performed. See text for details.
In the second series of experiments, the full-term developmental potential of the SCNT embryos produced with the most efficient HDACis treatments was assessed. In this case, HDACi-treated and nontreated SCNT embryos were cultured in vitro for 24 h and those that cleaved were transferred into the oviducts of 0.5 dpc pseudopregnant females. The pups were delivered by Cesarean section at 19.5 dpc and fostered to a lactating mother (Fig. 1).
Results
Histone acetylation levels in VPA-, TSA-, and nontreated SCNT embryos
The Ac-H3K14/DNA ratio 10 min after the injection of the cumulus cell nucleus into enucleated oocytes was 0.97. In control SCNT embryos that were not treated with HDACis, this ratio significantly decreased by 3 h after SCNT (p < 0.0001), when acetylation of H3K14 was practically undetectable, only to raise again to about half of the original ratio (0.40) by 6 h postactivation (Table 1 and Fig. 2).
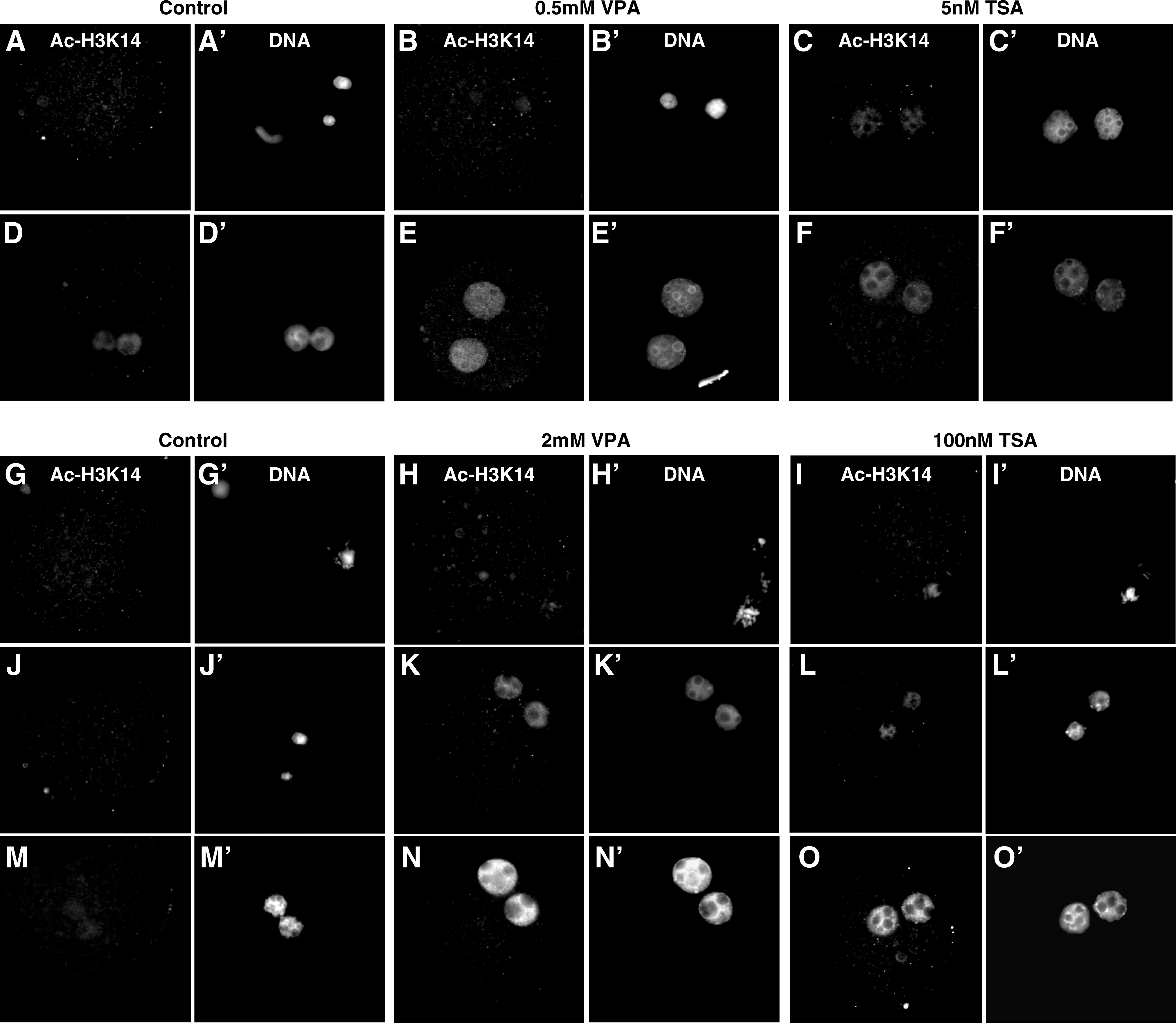
Acetylation level of H3K14 in HDACi-treated and nontreated SCNT embryos at different time points after reconstruction and activation. Upper panel: control SCNT embryo and SCNT embryos treated for 8 h with 0.5 mM VPA or 5 mM TSA during and after activation, fixed at 3 h (
Values with different superscripts within the same column differ significantly between treatments (P < 0.05).
When SCNT embryos were cultured in the presence of 0.5 mM VPA or 5 nM TSA during and after activation, higher Ac-H3K14/DNA ratios were observed by 3 h postactivation when compared to the control SCNT group, although the difference was only significant for the TSA-treated embryos (p < 0.001). However, by 6 h postactivation both HDACi treatments resulted in a similar and significant increase in H3K14 acetylation levels when compared to the control nontreated SCNT embryos (p < 0.01 for VPA and p < 0.05 for TSA). Because these two HDACi treatments were initiated 1–2 h post-SCNT, no analysis of histone acteylation levels were performed in these groups by 3 h post-SCNT.
In embryos exposed to 100 nM TSA immediately after SCNT, a much smaller decrease in the Ac-H3K14/DNA ratio was observed by 3 h post-SCNT than in the control group (0.47 vs. 0.02; p < 0.05). Although deacetylation of H3K14 occurred to a greater extend in 2 mM VPA than in 100 nM TSA-treated embryos during the first 3 h after SCNT (p < 0.05), by 3 h after activation the levels of acetylated H3K14 were similar in both groups (ratios of 0.53 and 0.59, respectively) and significantly higher than in the control (p < 0.001) and 0.5 mM VPA-treated (p < 0.01) groups. By 6 h postactivation, and similar to the results obtained for 0.5 mM VPA and 5 nM TSA treatments, both groups of oocytes treated with HDACis immediately after reconstruction had approximately recovered the initial Ac-H3K14/DNA ratios detected by 10 min after SCNT.
Effect of the VPA and TSA treatments on the in vitro development of SCNT embryos
Reconstructed oocytes treated with 5 nM TSA or 0.5 mM VPA during (6 h) and after (2 h) oocyte activation showed similar rates of embryonic development until the morula stage (Table 2). In terms of blastocyst formation, the 5 nM TSA treatment (31.8%) was statistically equivalent to the 0.5 mM VPA treatment (27.3%), but significantly higher than the control group (22.9%; p = 0.028). When treatments with 100 nM TSA or 2 mM VPA began immediately after the nuclear transfer (for 2–3 h) and continued during the oocyte activation period (for 6 h), the morula and blastocyst rates for the 2 mM VPA treatment were significantly increased when compared to the 0.5 mM VPA treatment used in the other set of experiments (63.6 vs. 50.6% and 38.3 vs. 27.3%; p = 0.005). In contrast, no significant differences were observed in the rates of morula or blastocyst formation between the two TSA treatments. Both the 2 mM VPA (38.3%) and the 100 nM TSA (34.5%) treatments resulted in a blastocyst development rate significantly higher than the obtained in the corresponding control group (25.1%; p = 0.001 and p = 0.035, respectively).
Values with different superscripts within the same column and category differ significantly between treatments (P < 0.05).
Embryo quality in blastocysts produced by SCNT after treatments with VPA, TSA, or control
The total and ICM mean cell numbers obtained for TSA- and VPA-treated and for control SCNT blastocysts are shown in Table 3. Blastocysts obtained with 0.5 mM VPA and 5 nM TSA treatments had a similar number of cells in the ICM (12 ± 4 and 14 ± 3, respectively), which were in both cases significantly higher than the mean in untreated SCNT-control embryos (8 ± 4; p < 0.001 for both comparisons). However, although in the 5 nM TSA treatment group the average number of cells in the ICM (14 ± 3) and in total (54 ± 14) were very close to those observed in parthenogenetically-activated controls (16 ± 4 and 60 ± 10, respectively), in the 0.5 mM VPA group they were significantly lower (12 ± 4 and 46 ± 12; p < 0.001 and p < 0.01, respectively). Blastocysts produced from either 2 mM VPA or 100 nM TSA had a higher total number of cells (57 ± 15 and 55 ± 15 vs. 41 ± 9; p < 0.001) and a higher number of ICM cells (14 ± 4 and 13 ± 4 vs. 9 ± 5; p < 0.001) than the SCNT control group. Analysis of the parthenogenetic blastocysts obtained in this series of experiments showed an average of total and ICM cell numbers (62 ± 13 and 16 ± 4, respectively) equivalent to the 2 mM VPA and 100 nM TSA groups.
Values with different superscripts within the same column and category differ significantly between treatments (p < 0.05).
Blastocysts produced from SCNT embryos treated with either 5 nM or 100 nM TSA had a similar number of total and ICM cells. The two treatments with VPA also resulted in blastocysts with equivalent ICM cell numbers, but in this case blastocysts produced with the 2 mM VPA treatment had a significantly higher total number of cells than those produced with the 0.5 mM VPA treatment (p < 0.05).
Full-term development of VPA, TSA, and nontreated SCNT embryos
Based on the previous results on the in vitro development and embryo quality, we selected 2 mM VPA and 100 nM TSA as the best treatments and the full-term development of the resulting SCNT embryos was next analyzed and compared to that of the control nontreated SCNT embryos. As shown in Table 4, we transferred a total of 190 two-cell stage embryos that had been treated with 100 nM TSA into the oviducts of 12 surrogate females and, as a result, 3 cloned pups developed to term and were born alive (1.6%). We also obtained, for the first time, full-term development from 2 mM VPA-treated embryos (2 cloned mice/270 transferred embryos into 15 surrogate females, 0.7%). No significant differences were observed between the VPA and TSA treatment groups. No full-term development resulted from nontreated control SCNT-embryos, despite the higher number of embryos that were transferred in this group (328 embryos into 18 surrogate females). As expected, the highest rates of full-term development were achieved when in vivo fertilized embryos were transferred (8 pups/18 transferred embryos, 44.4%).
Values with different superscripts within the same column and category differ significantly between treatments (p < 0.05).
All cloned pups obtained from VPA and TSA treatment groups had bodyweights that were very similar to those of noncloned pups (Table 4). The placenta weights of clones from VPA- and TSA-treated embryos were equivalent between each other (415.8 ± 108.7 and 377.8 ± 15.9 mg, respectively), but significantly heavier than those of noncloned pups produced from in vivo fertilized embryos (164.8 ± 21.4 mg; p < 0.001).
The two cloned females produced from VPA-treated embryos survived to adulthood and have proved to be fertile when mated to normal B6CBAF1 males, giving birth to several litters of 8–10 pups each. Of the three cloned females produced from TSA-treated embryos, one died unexpectedly at 3 weeks of age due to unknown reasons and a second one died at 3 months of age presumably due to complications during labor (she was pregnant with nine phenotypically normal pups). The third female in the TSA group has produced several litters and is still alive at 5 months of age.
Discussion
Reprogramming of differentiated somatic nuclei into a totipotent embryonic state by SCNT is not efficient. Although a considerable number of reconstructed oocytes might reach the blastocyst stage, their potential for full-term development is very limited (Wakayama et al., 2008). The reprogramming events following nuclear transfer occur mainly at an epigenetic level and one of the epigenetic pathways related to chromatin structure is the global level of acetylation of the nuclear histones (Yang et al., 2007). In this sense, and to facilitate nuclear reprogramming, SCNT embryos have been treated with the HDACi TSA to relax the epigenetic marks of the differentiated somatic cells, and this has resulted in a remarkable and significant increase in cloning efficiency in the hybrid B6D2F1 and the outbred ICR mouse strains (Kishigami et al., 2006a, 2007; Rybouchkin et al., 2006). More recently, improvement of mouse cloning efficiency by TSA has also been reported in the hybrid B6CBAF1 strain (Maalouf et al., 2009) and another HDCAi, scriptaid, has allowed the successful cloning of several inbred mouse strains (Van Thuan et al., 2009).
The HDCAi VPA has recently been shown to induce reprogramming of mouse fibroblasts transfected with transcription factors into iPS with a considerable higher efficiency than TSA (Huangfu et al., 2008a). Here we demonstrate that the HDACi VPA can also improve the in vitro developmental potential and the blastocyst quality of hybrid B6CBAF1 SCNT embryos, and that the VPA-treated SCNT embryos can give rise to healthy fertile cloned mice. However, contrary to the in vitro reprogramming results in iPS, VPA has a similar effect than TSA on cloning efficiency. To the best of our knowledge, this is the first report on the successful use of VPA for mouse cloning.
Two different protocols were used for the treatment of SCNT embryos with the two HDACis, based on previous work with TSA by Kishigami et al. (2006a) and Rybouchkin et al. (2006). Our results show that no significant differences exist between the two protocols in terms of in vitro development and embryo quality for SCNT embryos treated with TSA. In addition, the dynamics and levels of H3K14 acetylation, a marker for epigenetic reprogramming (Wang et al., 2007), were also similar in nuclei of embryos from the two TSA groups after activation. In contrast, when the two VPA protocols are compared, reacetylation of H3K14 after activation occurred faster in embryos subjected to the protocol in which the highest VPA concentration (2 mM) was used and the treatment was initiated immediately after nuclear injection, even though the final levels of H3K14 acetylation by 6 h after activation were equivalent in the two protocols. SCNT embryos treated with 2 mM VPA immediately after reconstruction also showed a higher in vitro developmental potential and a higher blastocyst cell number than those treated with the lowest VPA concentration (0.5 mM) starting 1–2 h after nuclear injection. Whether the faster kinetics of histone reacetylation and the increased developmental potential are due to the higher VPA concentration or to the different timing of the VPA treatment with regard to oocyte activation should be further investigated. However, the lack of significant differences between the two TSA protocols together with the limited increased in developmental potential and embryo quality induced by the 0.5 mM VPA treatment when compared to the control nontreated group, seem to indicate that a concentration of 0.5 mM VPA may be too low to exert beneficial effects. In this study, we chose 2 mM as the highest VPA concentration, as this was the concentration used by Huangfu et al. (2008a) to reprogram mouse embryonic fibroblasts to iPS. However, VPA concentrations higher than 2 mM could be tested in SCNT embryos.
Rates of blastocyst development in SCNT embryos treated with either 5 nM TSA during and after activation (31.8%) or 100 nM TSA or 2 mM VPA before and during activation (34.5 and 38.3%, respectively) were clearly superior to those of nontreated SCNT embryos (22.9–25.1%), confirming the beneficial effects of the TSA treatment on the in vitro developmental of cloned embryos reported by others (Kishigami et al., 2006a; Rybouchkin et al., 2006) and demonstrating the potential of VPA in mouse cloning experiments. In addition, these increased in vitro development rates in the HDACi-treated embryos were accompanied by increased levels of H3K14 acetylation and by an improved blastocyst quality, as judged by the increased number of total and ICM cells in comparison to the nontreated SCNT embryos (30–35% increase). In fact, increased histone acetylation has been associated with a more effective formation of DNA replication complexes (de Ruijter et al., 2003), which can facilitate cell proliferation and could explain these results. Differences in the level of H3K14 acetylation between the 2 mM VPA and the 100 nM TSA treatments by 3 h post-SCNT and between the 0.5 mM VPA and the 5 nM TSA treatments by 3 h postactivation, but not by 6 h postactivation for any of the HDACi groups, suggest that although both HDACis are equally effective in increasing histone acetylation levels, VPA has a slower kinetics than TSA independently of the concentration and protocol used.
Treatment of SCNT embryos with 100 nM TSA or 2 mM VPA before and during oocyte activation also allowed the obtention of viable cloned mice with a success rate of 1.6 and 0.7%, respectively. In contrast, we were unable to obtain any cloned mouse from the untreated SCNT embryos, despite the higher number of embryos that were transferred from this group. Although the cloning efficiencies obtained are probably too low to reveal statistically significant differences among the three groups of SCNT embryos, we believe that the lack of cloned pups in the control group (no HDACi treatment) is very relevant and indicates a positive effect of both TSA and VPA treatments on the full-term development of the cloned embryos. These results agree with the idea that inhibition of histone deacetylation might increase the number of cloned embryos that are able to correctly reprogram the somatic nucleus, reach full-term development, and give rise to healthy cloned mice (Wakayama et al., 2008).
Kishigami et al. (2006a) reported a rate of blastocyst development of 75% (from two-cell embryos) and a full-term development rate of 6% in SCNT embryos treated with 5 nM TSA for 10 h after activation. Rybouchkin et al. (2006) obtained 63% of blastocysts and a cloning efficiency of 2.8% from SCNT embryos treated with 100 nM TSA for 8 h immediately after nuclear injection and during activation. These rates of in vitro and full-term development of the SCNT embryos are higher than those obtained in the present study with either TSA or VPA. However, it is important to point out that the mouse strain used in our study is the hybrid B6CBAF1, whereas both aforementioned studies were performed with the hybrid B6D2F1 strain. Because of the well-known effect of the mouse strain on the efficiency of the SCNT technique (Gao et al., 2003; Van Thuan et al., 2009), it is possible that the use of B6CBAF1 mice in our study explains the differences observed. In fact, although the B6D2F1 strain has been widely used in cloning experiments, the use of B6CBAF1 mice has been much more limited and, therefore, fewer results on this strain are available. To our knowledge, only one study on the effects of TSA on cloning efficiencies has been published using B6CBAF1 mice (Maalouf et al., 2009). In this study, B6CBAF1 embryos reconstructed with cumulus cell nuclei were treated with 5 nM TSA for 10 h starting after activation and 34.9% of them developed to the blastocyst stage, a rate very similar to those obtained in our study with the best HDACi treatments (31.8–38.3%). The rates of full-term development of nontreated and treated SCNT embryos (0.3 and 3.1%, respectively) were also lower than those regularly obtained in the B6D2F1 strain by the group of Wakayama (Kishigami et al., 2006a; Van Thuan et al., 2009) and more similar to the rates obtained in the present study.
Although the bodyweights of the five cloned pups obtained in our study were very similar to those of pups produced from fertilized embryos, the placentas of all SCNT derived pups were significantly heavier. This indicates that neither TSA nor VPA are capable of normalizing placental overgrowth, a typical feature of cloned fetuses (Eggan et al., 2001). Despite this, the two clones derived from VPA-treated embryos and one out of three clones derived from TSA-treated embryos survived to adulthood, showing a normal phenotype and a normal reproductive ability. Even though the causes for the death of the two cloned females in the TSA group could not be determined, the fact that two of the three clones in the TSA group died prematurely while all the clones in the VPA group are still alive at 5–6 months of age is noteworthy. To our knowledge, no detrimental effects of the TSA treatment on the postnatal viability of the cloned mice have been indicated in the published studies. However, Meng et al. (2009) reported in rabbits that all the seven pups produced from TSA-treated embryos died of unknown reasons within 1 h to 19 days after birth.
In conclusion, we have demonstrated for the first time that VPA can improve the in vitro and full-term development of mouse SCNT embryos, at least in the hybrid B6CBAF1 strain, to the same level as TSA. Because cloning efficiencies are clearly influenced by the species or mouse strains used as nuclear donors, and because not all HDACis are equally effective in all genetic backgrounds (Van Thuan et al., 2009), further studies should be performed to analyze the effect of VPA on the cloning success in other species/strains. Our results support previous findings with other HDACis that suggest that the regulation of histone deacetylases is key for the efficient reprogramming of the somatic nucleus in SCNT. Further, the results presented here also demonstrate that VPA may improve nuclear reprogramming during SCNT, in addition to its previously reported beneficial effect on the nuclear reprogramming of somatic cells into iPS (Huangfu et al., 2008a, 2008b). Our findings may open new opportunities to understand the mechanisms of nuclear reprogramming during SCNT and to improve cloning efficiencies.
Footnotes
Acknowledgments
The authors thank Marc Puigcerver and Jonatan Lucas for technical assistance and the staff at the Servei d'Estabulari for taking care of the animals. This work was supported by the Spanish Ministerio de Educación y Ciencia (BIO 2006-11792) and the Generalitat de Catalunya (2005-SGR00437). N.C.B. is a fellow of the Portuguese Fundação para a Ciência e Tecnologia.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.