
Abstract
Abstract
Somatic cell nuclear transfer (SCNT) holds potential as a useful tool for agricultural and biomedical applications. In vitro development of marbled cat intergeneric SCNT reconstructed into domestic cat cytoplast revealed that cloned, marbled cat embryo development was blocked at the morula stage. No pregnancies resulted from the transfer of one- to eight-cell stage embryos into domestic cat surrogate mothers. This suggested that abnormalities occurred in the cloned marbled cat embryos, which may be associated with incomplete reprogramming during early embryo development. Two pregnancies were established in surrogate mothers that received cloned domestic cat embryos, but SCNT offspring developed abnormally. Some specific phenotypes that were observed included incomplete abdominal wall disclosure, improper fetal development. In addition, some of the fetuses were mummified or stillbirths. The two live births died within 5 days. Telomere lengths of cloned kittens as determined by qualtitative polymerase chain reaction (qPCR) were inconclusive: some were found to be shorter, longer, or the same as donor control cells. Our findings support the hypothesis that telomere lengths do not govern the health of these cloned animals. A lack of complete reprogramming may lead to developmental failure and the abnormalities observed in cloned offspring.
Introduction
Telomeres, the termini of eukaryotic chromosomes, are composed of long tandem repeated TTAGGG sequences. Telomere functions involve stabilization, replication, and segregation of the chromosomes during cell division (Blackburn, 1991). Each time a cell divides, it gradually loses its telomeric sequences due to the inability of DNA polymerase to completely replicate the 3′ end of the linear DNA. This process ultimately leads to cell cycle arrest and cellular senescence (Olovnikov, 1973). The first cloned sheep, Dolly, was derived from a cell of a 6-year-old sheep that exhibited shortened telomeres, which resulted in cellular senescence and further shortening of her own telomeres (Shiels et al., 1999). Since then, restoration of telomere length in cloned animals has been intensely studied. Questions regarding the relationship between telomere length and cloned animals' life spans remain to be elucidated. In SCNT, cloned embryos need reprogramming to convert differentiated donor nuclei into undifferentiated states to reestablish the correct pattern of gene expression, allowing for normal embryogenesis. Moreover, these embryos require appropriate cellular rejuvenation due to their generation from aged and cultured propagated somatic cells expressing shortened telomeres. Determination of telomere restoration processes and possible physiological effects of SCNT on the senescence of the cells is pertinent to current research on telomeres in cloned animals. Several studies have measured the telomere length of cloned animals compared to their age-matched control counterparts. These results reported that some telomeres were shorter (Clark et al., 2003; Miyashita et al., 2002; Shiels et al., 1999), comparable (Jiang et al., 2004; Kubota et al., 2004), or longer (Lanza et al., 2000) than their controls, suggesting that SCNT reprograms telomere length in different ways dependent on experimental methods employed. No telomere research has been reported on cloned feline species.
In our study, intergeneric SCNT (iSCNT) between two feline species of different genera, the marbled cat (Pardofelis marmorata) and domestic cat (Felis catus), were conducted. Male ear skin fibroblasts derived from either marbled or domestic cats were used as donor nuclei and transferred into enucleated domestic cat oocytes. To evaluate the in vivo developmental competence, reconstructed intergeneric cloned marbled cat and cloned domestic cat embryos were transplanted into domestic cat surrogate mothers. To gain knowledge of telomere length restoration dynamics in cloned feline species, we investigated whether nuclear reprogramming restored the cellular age of donor cells after SCNT by assessing the telomere lengths in the cloned newborn offspring using quantitative real-time polymerase chain reaction (qPCR). This method is simple, rapid, sensitive, quantitative, and has higher inherent reproducibility for measuring the number of telomere repeats than conventional methods (Cawthon, 2009; O'Callaghan et al., 2008).
Materials and Methods
All animal experiments were approved by and performed according to the guidelines of the Ethics Committee of the Laboratory Animal Care of Suranaree University of Technology. Unless stated otherwise, all reagents were purchased from Sigma Chemical Company (St. Louis, MO, USA). Donor cell preparation, in vitro maturation of oocytes, and in vitro production of cloned embryos in feline species were performed as previously described (Imsoonthornruksa et al., 2010). In vitro culture of oocytes and embryos was performed at 38°C in a humidified atmosphere of 5% CO2, 5% O2, and 90% N2.
Preparation of donor cells
Adult fibroblasts were generated from the ear skin tissues of 2-year-old domestic cats and a 12-year-old marbled cat from Nakhon Ratchasima Zoo, Thailand. The tissues were cut into small pieces and cultured in alpha modified minimum essential medium (αMEM) supplemented with 10% fetal bovine serum (FBS; Gibco-BRL, Grand Island, NY, USA) at 37°C in a humidified atmosphere of 5% CO2 in air. Tissues were cultured until fibroblast cell outgrowth was apparent. The primary culture was harvested and subcultured to multipassages. Fibroblasts were frozen at −80°C in αMEM +10% FBS and 10% (v/v) dimethylsulfoxide (DMSO) for 24 h before storage in liquid nitrogen for future use as donor cells. Only passage 3 donor cells were used for SCNT. Prior to nuclear transfer, frozen fibroblasts were thawed and cultured in αMEM + 10% FBS for 2–3 days. Nonquiescent fibroblasts were trypsinized at subconfluence and used as donor cells.
Maturation of cat oocytes
Healthy female domestic cats (9–36 months of age) were superstimulated by 200 IU equine chorionic gonadotropin (eCG, Intervet, Booxmer, The Netherlands) treatment. The superstimulated cats were ovariectomised 5 days after treatment. The ovaries were repeatedly sliced with a 21-gauge needle to release the cumulus oocyte complexes (COCs). These COCs were collected and eventually matured in TCM 199 supplemented with 0.36 mM sodium pyruvate, 2.2 mM calcium lactate, 2 mM L-glutamine, 1.13 mM cysteine, 0.3 mg/mL bovine serum albumin (BSA), 0.5 IU/mL eCG, and 1 IU/mL human chorionic gonadotropin (hCG, Intervet) for 24 h.
SCNT, iSCNT, parthenogenetic activation (PA), and embryo culture
Cumulus cells were removed from COCs by gentle pipetting in 0.2% hyaluronidase. Mature oocytes with a noted first polar body (PB) were incubated in Emcare holding medium (ICP Bio, Auckland, New Zealand) supplemented with 5 μg/mL cytochalasin B. Zonae pellucidae above the first PB of oocytes were cut using a fine glass needle. Further, the PB and metaphase plate were squeezed out with a small volume of surrounding cytoplasm. Successful enucleation of each oocyte was confirmed by 5 μg/mL Hoechst 33342 staining of the removed cytoplasm and by observation under ultraviolet light to ensure complete removal of the oocyte nuclei. After enucleation, a single domestic or marbled cat fibroblast was inserted into the perivitelline space of the enucleated oocyte. Fusion of cytoplast–cell couplet was performed in mannitol fusion medium between the tips of the fusion electrode. This was followed by electrostimulation using double DC pulses of 30 V for 30 μsec delivered by an electrofusion machine (Voltrain EP-1; Cryologic, Mulgrave,Vic., Australia). Fused oocytes and matured ones (for PA as control group) were activated with 7% ethanol for 5 min, followed by a 5-h incubation in 10 μg/mL cyclohexymide and 1.25 μg/mL cytochalasin D.
SCNT, iSCNT, and PA embryos were cultured in Tyrode's medium supplemented with 1% MEM nonessential amino acids (NEAA), 0.3 mg/mL BSA, 0.36 mM sodium pyruvate, 2.2 mM calcium lactate, and 2 mM L-glutamine. All eight-cell stage embryos on day 2 of culture were selected and cultured in the same medium without BSA, but were supplemented with 2% MEM essential amino acid (EAA) and 10% FBS until day 8. Half of the medium was changed and embryo development was assessed daily.
Embryo transfer
Female domestic cats age 12–18 months were hormonally stimulated by 200 IU eCG intramuscularly. Five days after administration, 200 IU hCG was intravenously administered to induce ovulation. Approximately 48 h after the hCG injection, the SCNT or iSCNT embryos at one- to eight-cell (Fig. 1A and B) stages were loaded into the embryo transfer catheter (K-JITS-303525, COOK, Queensland, Australia) (Fig. 1C) and gently inserted into the oviduct of the stimulated cat to deposit the embryos. The pregnancies of surrogate mothers were determined by palpation 45 days after embryo transfer and confirmed by X-ray photography on day 50 after embryo transfer. Pregnant mothers were allowed to complete gestation and obstetrical assistance was available if needed.
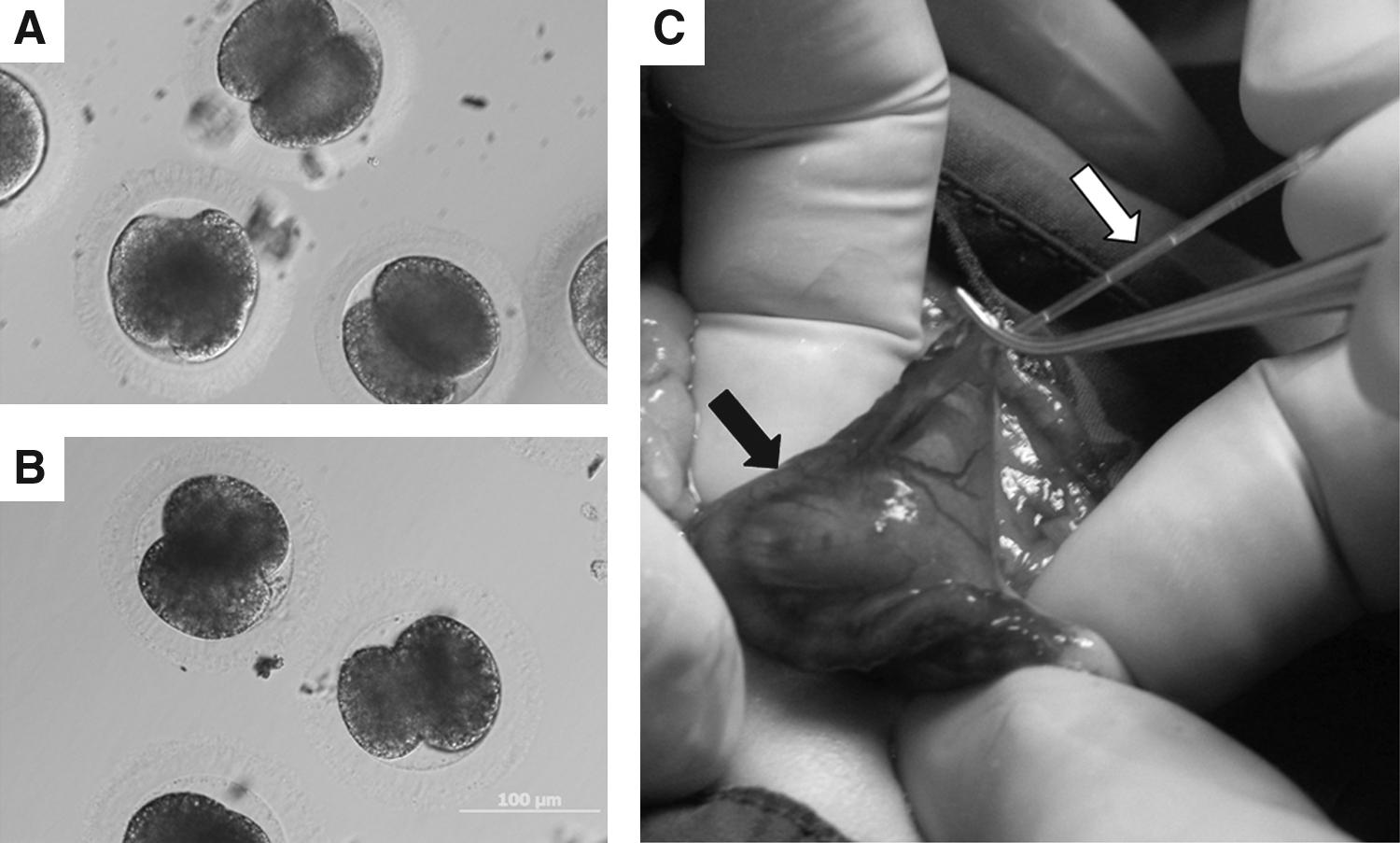
Cloned domestic cat (
Microsatellites analysis
Genomic DNA was extracted from donor cells and muscle samples from cloned kittens and surrogate mothers using the DNeasy Tissue kit (Qiagen, Ontario, Canada) according to the manufacturer's instructions. Five feline microsatellite markers (FCA170, FCA201, FCA224, FCA229, and FCA441, Menotti-Raymond et al., 1999) were amplified and PCR products were resolved on a 20% PAGE to confirm the genetic identity of the cloned kittens and donor fibroblast cells.
Absolute quantitative telomere length
Triplicate qPCR reactions for telomeric and granulocyte colony-stimulating factor (G-CSF, a single copy gene in feline species) (Yamamoto et al., 2001) assays were carried out in a 20-μL final volume with a final concentration of 1×DyNamo™ HS SYBR Green® qPCR kit (NEB, MA, USA), 900 nM and 400 nM of each telomeric and G-CSF primer, respectively, and approximately 50 ng of genomic DNA. qPCR assays were performed in the Bio-Rad Chromo 4 Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA). Standard curves were generated for both telomere length and G-CSF genes using dilution of known quantities of pGEM-T-easy vectors carrying a synthesized 84-bp oligonucleotide containing 14 tandem repeats of TTAGGG and G-CSF PCR product, respectively, following the methods by O'Callaghan et al. (2008). Forward and reverse telomeric primers were telg 5′-ACACTAAGGTTTGGGTTTGGGTTTGGGTTTGG GT TAGTGT-3′, and telc 5′-TGTTAGGTATCCCTATCCCT ATCCCTATCCCTATCCCTAACA-3′, respectively (Cawthon, 2009). Sequences of the G-CSF primers designed for this study were G-CSF forward 5′- GTTTCATTCTCCTG CTTGTAG-3′ and G-CSF reverse 5′- GTTCACTGGCGTTC TTCC-3′. Amplification settings were as follows: initial heating to 95°C for 15 min; 2 cycles of 94°C for 20 sec; 49°C for 20 sec; 20 cycles of 94°C for 20 sec; 60°C for 20 sec; and finally 72°C for 20 sec for the telomere reactions or 30 cycles of 94°C for 20 sec, or 50°C for 20 sec and 72°C for 20 sec for the G-CSF reactions. A melting curve analysis was performed for each sample to verify that only a single specific product was generated for each reaction. The melting cycle settings were as follows: 95°C for 20 sec, 65°C for 20 sec, and a step cycle starting at 65°C to 95°C in 0.2°C increments.
To convert the resulting cycle threshold (CT) values into telomere lengths, the number of telemetric repeats and G-CSF copies per reaction were calculated by the method of O'Callaghan et al. (2008). After the thermal cycling reactions and raw data collections were completed, Opticon Monitor 3 software (Bio-Rad) was used to produce a value for each reaction equivalent to kb/reaction based on the telomere standard curve values. The telomere kb per reaction value was divided by the copy number of G-CSF (calculated from G-CSF CT values and standard curve) to give a total telomeric length in kb per diploid genome.
Statistical analyses
SCNT, iSCNT, and PA were repeated at least four times in each treatment group. Quantification of telomere length was carried out in triplicate. Statistical analysis of the data was conducted using completely randomized design (CRD) with the Statistical Analysis System (SAS Inst. Inc., Cary, NC, USA). Analysis of variance (ANOVA) was used to determine significant differences between samples. Mean comparisons by Duncan's Multiple Range Test (DMRT) was used to determine differences between means at p<0.05.
Results
Feline cloned and PA embryo development
To evaluate the capability of using domestic cat oocytes as recipient cytoplasts in iSCNT, ear skin fibroblasts of marbled cats were fused with domestic cat oocytes. In vitro developmental rates of the iSCNT embryos at the cleavage and blastocyst formation stages were assessed and compared with domestic cat SCNT and PA embryos (Table 1). No differences were observed in the fusion rate when domestic or marbled cat fibroblasts were used as donor nuclei. Cleavage rates were similar between the PA and cloned embryos. However, the development to eight-cell and morula stages was lower in the marbled cat iSCNT group compared to both PA and domestic cat SCNT embryos. The blastocyst formation rate in cloned domestic cats (28.3%) was comparable to those of PA (26.3%). But no blastocyst formation was observed in marbled cat iSCNT.
Percentages calculated from the total number of oocytes used.
Percentages calculated from the number of fused couplets that cleaved and developed to each stage. In PA, numbers cleaved and developed to each stage were calculated from total number of oocytes used. Within a column, values with different superscript letters differ significantly (p<0.05).
The cell number of morula stage embryos was confirmed by Hoechst 33342 staining (data not shown).
Embryo transfer and pregnancy results
A total of 247 domestic and 461 marbled cat reconstructed embryos, between one- and eight-cell stages (Fig. 1), were transferred into 24 surrogate mothers (see Supplemental Data Table S1; see online supplementary data at www.liebertonline.com/cell). The pregnancy results are shown in Table 2 and Supplementary Table S1. The average number of embryos transferred to each mother was 20.6 and 38.4 for cloned domestic and marbled cat embryos, respectively. At 45 days after embryo transfer, two pregnancies from surrogate mothers with cloned domestic cat embryos were detected, but no pregnancies from surrogate mothers with cloned marbled cat embryos were observed. The first pregnant mother (A2) underwent a caesarean section at 64 days after embryo transfer. The single kitten (A2/1) died prenatally with incomplete closure of its abdominal wall. The musculature and abdominal organs protruded externally (Fig. 2B). The second pregnant mother (A10) delivered six kittens naturally at 56 days after embryo transfer. These kittens were of various weights and stages of development, as outline in Table 3. Only two kittens were born live (Fig. 2E). One kitten (A10/2) was stillborn and one was mummified (A10/4) (Fig. 2D). Two were born with incomplete fetal development (A10/1 and A10/6) (Fig. 2C). Of the two live born kittens, one died postnatally within 24 h (A10/3) and another died 5 days after birth due to dehydration from diarrhea (A10/5). Subsequent DNA analysis of five feline microsatellite markers confirmed that these kittens were genetically identical to the donor nuclei (Fig. 3).
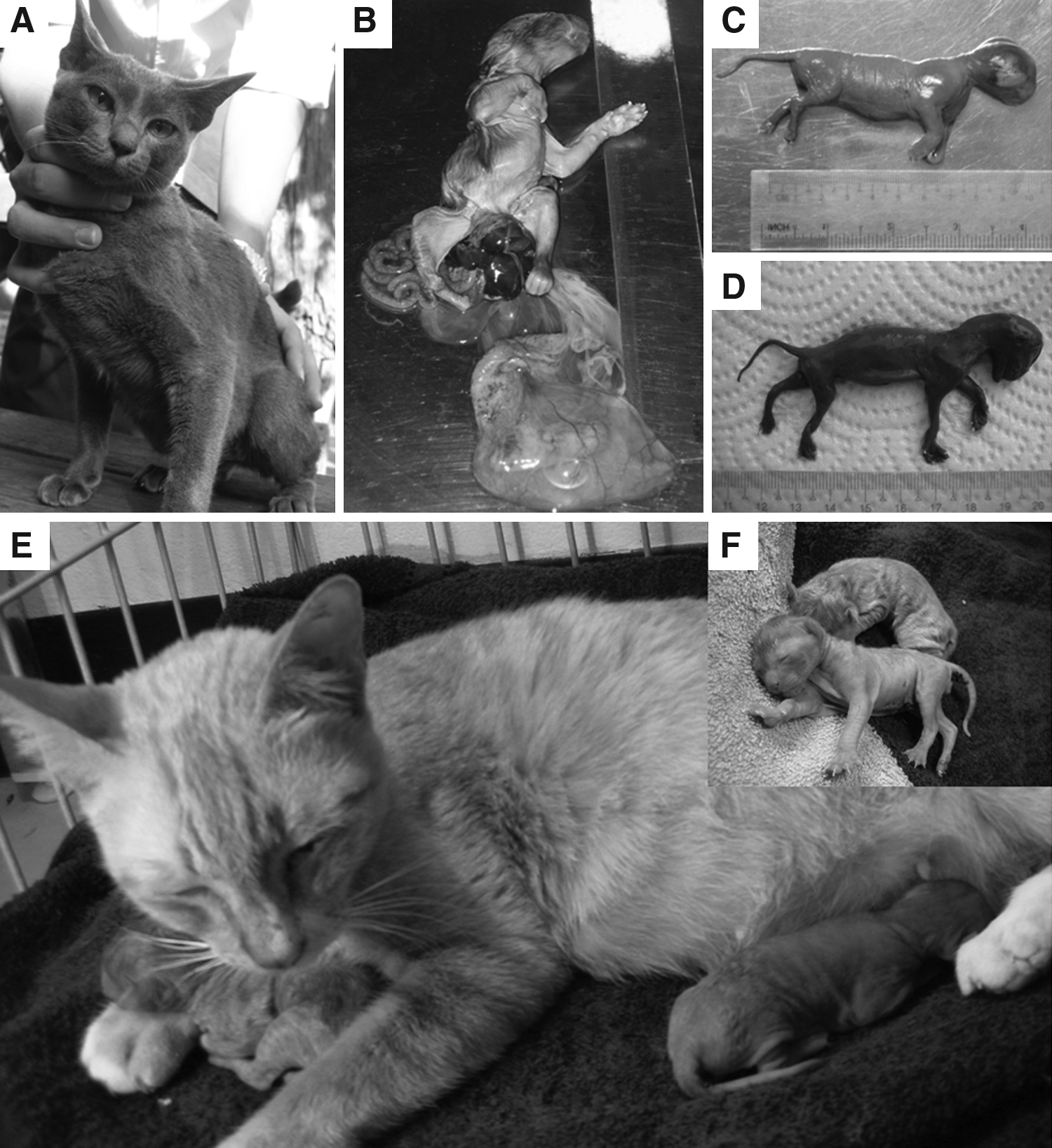
Somatic cell donor (
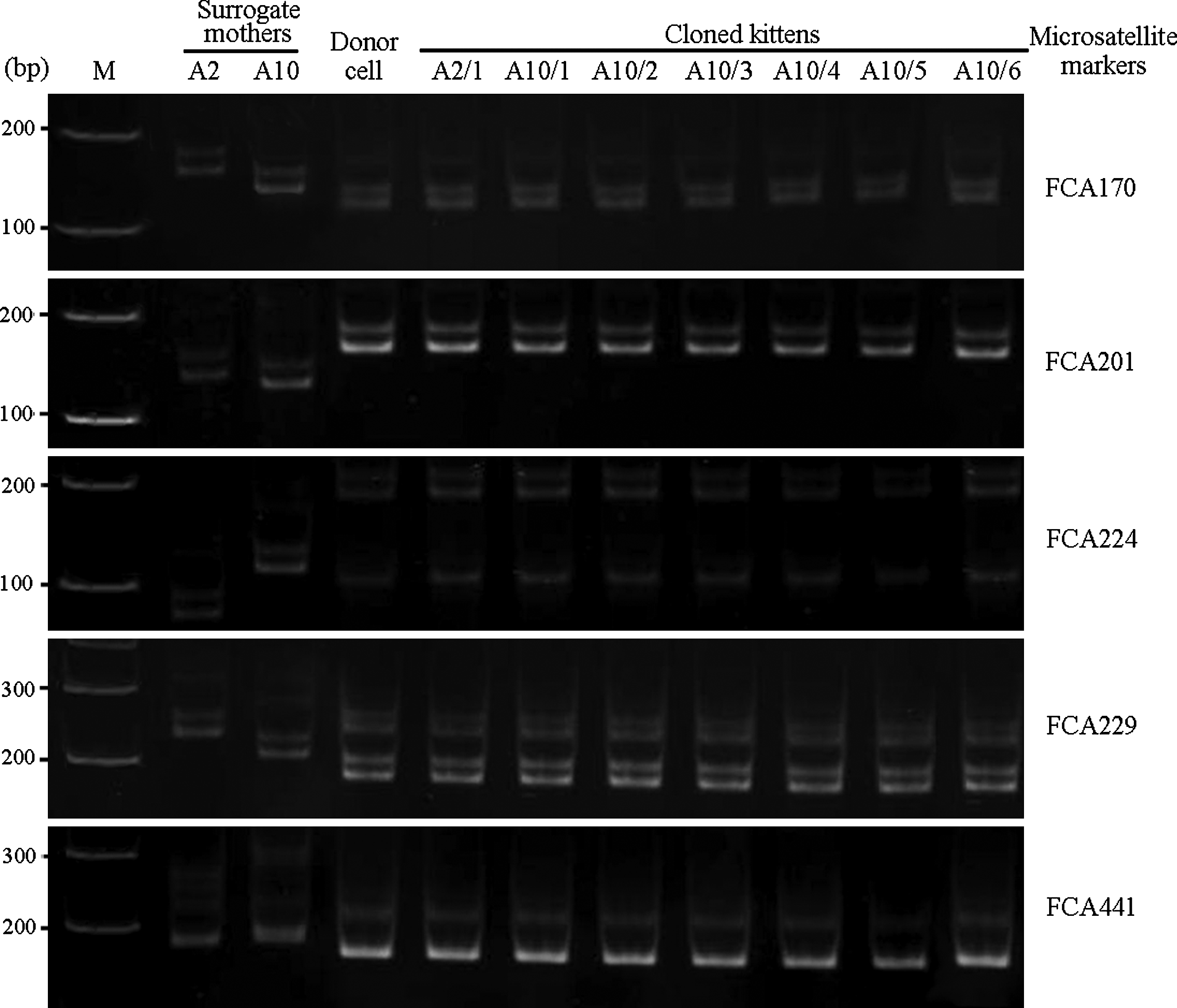
Polyacrylamide gel electrophoresis of feline microsatellite markers of donor nuclei, cloned kittens, and their surrogate mothers, M=DNA markers.
The kittens were numbered by the mother and in the order of delivery.
Absolute quantitative telomere length analysis
Muscle samples were collected from cloned kittens (Table 3) to measure the telomere length of chromosomes of muscle cells compared to the muscle cell DNA of both surrogate mothers and the donor cells at passage 3 by qPCR. The detection of absolute telomere length of each sample was included in the formula to calculate telomere length in kb per reaction from a linear standard curve. The number of diploid genomes was calculated using a cycle threshold and standard curve for G-CSF for a single copy gene product (Fig. 4A). The telomere length in kb per reaction of each sample was divided by diploid genome copy number to give the absolute telomere length (kb) per diploid genome (Fig. 4B). qPCR of cloned kittens' telomeric DNA revealed that each seemed to independently rebuild telomeres to different lengths. The 24-month-old male donor cat fibroblasts had a telomere length of approximately 37 kb per diploid genome. Muscle cells from the two surrogate mothers (age 12–18 months) had telomere lengths of approximately 27–28 kb per diploid genome. We observed that of seven cloned kittens, two (A2/1 and A10/3) had significantly shorter telomere lengths (25 and 24 kb per diploid genome, respectively), and two (A10/1 and A10/6) showed telomere lengths comparable to donor cells (38 and 40 kb per diploid genome, respectively). Three of these cloned kittens (A10/2, A10/4, and A10/5) had significantly longer telomere lengths (59, 46s, and 58 kb per diploid genome, respectively) than that of their donor cells (37 kb, p<0.05) (Fig. 4B).
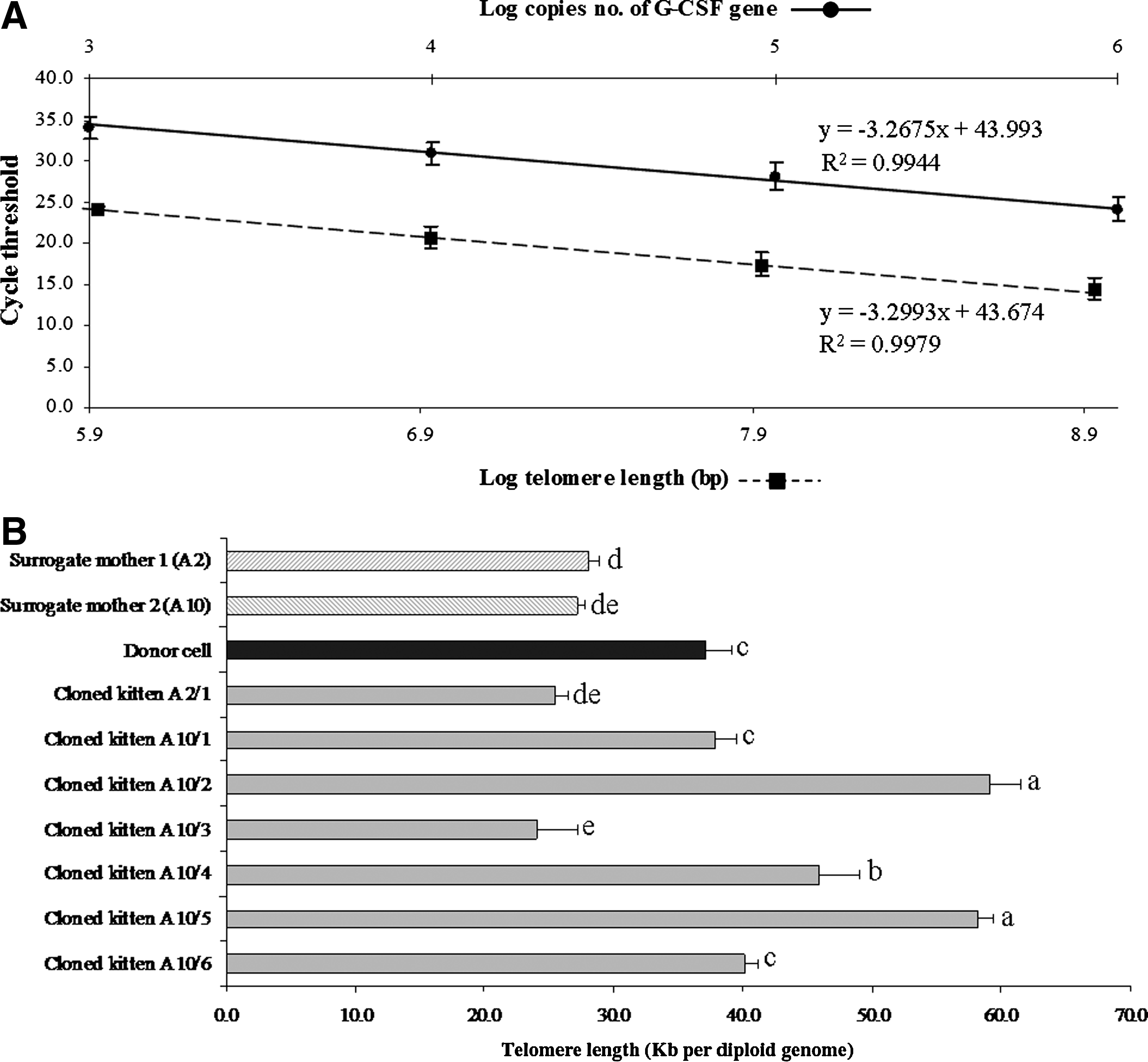
Analysis of telomere length by qPCR. (
Discussion
Since the birth of Ditteaux, the African wildcat cloned by iSCNT and reported by Gomez et al. in 2004, interest has arisen in cloning several different species (Gomez et al., 2006). In some iSCNT cases, a process by which cloned embryos are generated from donor cell and recipient cytoplasts of different genera, pregnancy has been successful, but no live kittens have been reported (Thongphakdee et al., 2010; Yin et al., 2006). In this study, marbled cat iSCNTs were used to evaluate the in vitro and in vivo developmental competence of embryos reconstructed with domestic cat cytoplast. The in vitro development of the cloned embryos revealed that somatic fibroblasts of marbled cats could be fused and cleaved when reconstructed with domestic cat cytoplasts, but arrested development occurred at the morula stage. Similar developmental results at the morula stage for both domestic cat SCNT and marbled cat iSCNT have also been reported (Thongphakdee et al., 2006; Wen et al., 2003). Although cloned marbled cat embryos did not develop to the blastocyst stage in vitro, we believe that an in vivo system may hold more potential to support embryo development. The early cleaved cloned marbled cat embryos were transferred into oviducts of surrogate mothers at an average of 38 embryos per mother. Cloned domestic cat embryos were separately transferred as a control group with an average of 20 embryos per mother. No pregnancies resulted when surrogate mothers received cloned marbled cat embryos. However, surrogate recipients of cloned domestic cat embryos resulted in a 17% pregnancy rate. Pregnancy rates from cloned feline embryos vary from 11% to 65% in the literature (Gomez et al., 2009, 2011; Shin et al., 2002; Yin et al., 2006, 2008). These comparisons may be inaccurate, however, because pregnancy rate depends the donor cell type, discrepancies between laboratory procedures of SCNT, and embryo transfer methods. Pregnancy failure in the marbled cat iSCNT group may be attributed to incomplete reprogramming of the donor cell (Imsoonthornruksa et al., 2010) over technique. Multiple factors affecting iSCNT pregnancy rate have been reported, including SCNT procedure, mitochondrial heteroplasmy, insufficient nucleocytoplasmic interaction, abnormality of nuclear reprogramming, number of transferred embryos, in vivo developmental block, and the genetic background of surrogate mothers and cloned embryos (Chen et al., 2002; Gomez et al., 2004, 2008; Thongphakdee, et al., 2008; Wen et al., 2003; Yin et al., 2006). Although marbled cat iSCNT pregnancy was not achieved in this study, the births of cloned domestic kittens were successful. Thus, under our SCNT procedure, transfer of 15 to 20 cloned domestic cat embryos per mother seems sufficient to confer a successful pregnancy (see Supplemental Table 1S). In 2004, Gomez and colleagues showed that the transfer of at least 30 iSCNT African wildcat embryos per recipient was required to establish pregnancy. For a domestic cat, 65 skin and 140 fetal fibroblast SCNT embryos (Yin et al., 2005) and 25–28 fibroblast SCNT embryos (Yin et al., 2008) were needed to ensure pregnancy. A theory explaining a needed lower number may involve our system incorporating a high percentage of cloned domestic cat embryos that had already developed to blastocyst stage (28%). This is actually higher than the 3–6.5% in previous reports (Yin et al., 2005). However, the cat iSCNT procedure in Gomez et al., 2008 reported that successful in vitro development to the blastocyst stage occurred in 6–43% of cases and was dependent on cell treatment. Requirements for the successful transfer of embryos correlate with their development potential to the blastocyst stage and to the uterus stage of the surrogate mother. Success of SCNT in felines regarding development varied from 0.5–20% of total embryos transferred to domestic cat recipients (Gomez et al., 2011; Shin et al., 2002; Yin et al., 2006, 2008). This low survival rate is most likely dependent on synchrony between embryos and the uterine endometrium, which is essential for pregnancy and healthy offspring (Pope et al., 2006). In this study, two pregnancies of surrogate mothers that received cloned domestic cat embryos were observed. One mother delivered by caesarean section 64 days after embryo transfer, but this cloned kitten died prenatally with incomplete closure of the abdominal cavity. This observation was similar to those of the African wildcat and sand cat cloned kittens (Gomez et al., 2004, 2008). The second mother delivered naturally 56 days after embryo transfer, but these offspring suffered from incomplete fetal development: one was stillborn, one mummified, and two were live births. These live births, however, were premature and may have been caused by spontaneous abortion to eliminate the dead fetuses. Both died shortly after birth from conditions typically associated with premature delivery. Therefore, a low pregnancy rate following embryo transfer, high rate of abortion during early gestation, and fetal abortion and resorption (Gomez et al., 2004, 2008; Yin et al., 2006, 2008) are all expected difficulties in cloning feline embryos.
Little is known about telomere length in feline species. Telomere lengths in cloned kittens have not been previously reported. We used qPCR to measure telomere length in muscle tissue cells of seven cloned kittens. This method is a proven, useful tool to rapidly and precisely quantify telomere length (Cawthon, 2009; O'Callaghan et al., 2008), and surpasses the Southern blot method for measuring terminal restriction fragment (TRF) lengths. O'Callaghan et al. (2008) and Cawthon (2009) reported that telomere lengths measured by qPCR were highly correlated with TRF length, but were more accurate and reproducible in independent runs. Several changes occur to somatic cell nuclei after incorporation into unfertilized cytoplasts. A major contributor to the success of nuclear reprogramming in cloned animals is telomerase activity (Marión and Blasco, 2010). Our study marks the first use of qPCR to determine the absolute telomere length in cloned kittens. Quantitative real-time amplification of the telomere sequences was performed as described by O'Callaghan et al. (2008) and Cawthon (2009) with slight modifications. Our assay measured an average telomere length ratio by quantification of the telomeric DNA sequence with specially designed primers. This ratio was divided by the quantity of a single copy gene. The results of this study revealed that the telomere length was not a key element in determining the live birth percentage of cloned kittens. The telomere lengths varied from Brummendorf et al. (2002), which described telomere lengths of domestic cat blood cells (lymphocytes and granulocytes) of 2-week-old newborn kittens and 24-month-old cats as 70 kb and 50–60 kb, respectively. Our findings revealed that telomere length variation among newborn kittens can be attributed to nuclear reprogramming errors. Alterations in telomere length in cloned animals could potentially be explained by the abnormal resetting of telomere length via aberrant expression of telomerase during embryo development (Lanza et al., 2000; Shiel et al., 1999). The relationship between telomere length and life span in cloned animals remains to be elucidated, and the contribution of telomere restoration is still under debate. Our results demonstrate that cloned embryos inherit genomic modifications by the interaction of donor cell nuclei and cytoplasmic factors in the SCNT process, but telomere length reprogramming processes during in vitro and in vivo development of cloned embryos varies and cannot be predicted. The discrepancy of telomere length after SCNT in different reports may be due to factors such as species, age, gender, donor cell type, sampling, and measuring protocol, efficiency of telomerase reactivation, and timing during nuclear reprogramming (Bett et al., 2001; Lanza et al., 2000; Miyashita, et al., 2002; Shiels and Jackine 2003; Tian et al., 2000).
Our results indicate that in vitro and in vivo development of the iSCNT marbled cat in domestic cat oocytes was incomplete. Further understanding of the mechanisms that control the nuclear reprogramming of the donor cell is critical for increasing the number of endangered species that may benefit from this technique. In this study, live birth cloned domestic cats proved successful, but cloned kittens died shortly after birth. This result demonstrated abnormal development of the cloned fetuses, anatomic abnormality of the cloned offspring, and postpartum mortality and low production efficiency of healthy cloned kittens. Importantly, these problems have been encountered in other studies (Gomez et al., 2004, 2006, 2008; Yin et al., 2006). Further, we described the application of qPCR as a tool to evaluate telomere lengths of cloned kittens. This observation demonstrated that the shortened telomere length of the aged donor cell is restored to various lengths—shorter, longer, and the same as those of donor cells after SCNT, in vitro, and in vivo development, and regardless of the cloned kitten viability. These results suggest that short telomeres are not the sole contributors to frequent neonatal death of cloned animals. The abnormalities that occur in cloned animals are intriguing and warrant long-term studies to better understand SCNT technology.
Footnotes
Acknowledgments
Nakhon Ratchasima Zoo provided the marbled cat tissue. Our sincere thanks go to Dr. P. Grote for his critical reading of the manuscript. This work was partially supported by Suranaree University of Technology (SUT) Research and Development Fund and the Higher Education Research Promotion and National Research University Project of Thailand, Office of the Higher Education Commission. S. Imsoonthornruksa and K. Srirattana were supported by SUT postgraduate research fellowships.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.