
Abstract
Abstract
Recently, the search for stem cells has become focused on fetal appendages such as the amniotic membrane and umbilical cord. Previously, we have shown the existence of stem cells in the amniotic membrane that can differentiate into various cells. In this study, we attempt to characterize and maintain the stemness characteristics of mesenchymal stem cells (MSCs) for Wharton's jelly, an inherent tissue of the umbilical cord. Wharton's jelly cells (WJCs) were isolated, adhered to culture plates, and characterized for stem cell and surface markers expression. They expressed the embryonic stem cell markers Nanog, Oct ¾, and Sox2. On flow cytometric analysis, WJCs predominantly expressed the MSC markers CD73, CD90, and CD105 and did not express the hematopoietic lineage markers CD14, CD34, CD45, and HLA-DR. In floating culture, WJCs could maintain stemness, and they could differentiate to osteogenic, chondrogenic, and adipogenic lineages. In conclusion, WJCs satisfy the criteria of MSCs. Given that extraction of the umbilical cord is not invasive, and the umbilical cord can be obtained without ethical and technical issues, we suggest that WJCs, after maintaining stemness, have a potential contribution to medical treatment for patients, even newborns, with congenital skeletal and cartilage disorders.
Introduction
Because Wharton's jelly cells (WJCs) have high proliferative potentiality, many researchers have recently suggested the potential existence of stem cells in the umbilical cord. Carlin et al. (2006) reported the expression of transcription factors Oct-4, Sox-2, and Nanog by porcine umbilical cord matrix cells. Weiss et al. (2006) also showed the expression of transcriptional factors in human umbilical cord matrix cells, but the expression of these transcriptional factors was at low levels.
The umbilical cord contains various cells that have potential for therapy, but it is routinely discarded after birth. In contrast, cord blood cells have been established as a source of stem cell transplantation (Habich et al., 2006; Terai et al., 2005). This source has the advantage that cells are isolated from the fetal structure in the perinatal period as umbilical cord blood and thus may be better tolerated following transplantation, with less incidence of graft-versus-host disease (Ding et al., 2011).
In amniotic membrane, the presence of mesenchymal stem cells (MSCs) is indicated (Zhao et al., 2005). The minimal criteria for multipotent mesenchymal stromal cells have been defined (Dominici et al., 2006; Parolini et al., 2008). First, MSCs must be plastic-adherent when maintained in standard culture conditions and form fibroblast colony-forming units. Second, MSCs must express CD73, CD90, and CD105 and lack expression of hematopoietic markers, such as CD14, CD34, CD45, and HLA-DR. Third, MSCs must differentiate to osteoblasts, adipocytes, and chondroblasts in vitro (Parolini et al., 2008). MSCs have great therapeutic potential due to their capacity of self-renewal and multilineage differentiation.
Recently, many authors have tried to isolate Wharton's jelly MSCs in human and animals and characterize these cells according to viability, survival, immunogenicity, and differentiation to different lineages (Garzon et al., 2012; Iacono et al., 2012; Kim et al., 2011; Lupu et al., 2011; Penolazzi et al., 2012; Ruan et al., 2012; Salehinejad et al., 2012; Zhou et al., 2011).
In this study, we attempted to characterize and maintain the stemness characteristics of human WJCs.
Materials and Methods
Cell cultures
Human umbilical cords were obtained from term cesarian deliveries, and informed consent was obtained. The study and the use of the human umbilical cords were approved by the Ethics Committee of the University of Toyama. Umbilical cords were collected immediately after birth and placed in sterile phosphate-buffered saline (PBS). Then, the umbilical cords were transported to the laboratory and preserved in PBS at 4°C. All samples were processed within 24 h. After rinsing in 0.9% physiological saline, umbilical cords were cut into 2- to 3-cm lengths. Umbilical arteries, vein, and outer membrane were mechanically removed and discarded. The remaining jelly was minced into small fragments, and then placed in Dulbecco's modified Eagle medium (DMEM; Sigma-Aldrich Co., St. Louis, MO, USA) containing 0.2 % collagenase at 37°C. After 3 h, PBS containing 0.025% DNase I and 0.03% hyaluronidase (Sigma-Aldrich Co., St. Louis, USA) was added, and the mixture was incubated for 30 min. The digested mixture was washed three times with PBS and placed in DMEM with 2.5% trypsin (Sigma-Aldrich) for 30 min at 37°C. After washing three times with PBS, the cells were resuspended in DMEM supplemented with 2 mM
Reverse transcription polymerase chain reaction
Total RNA was extracted from WJCs using ISOGEN kits (Nippon Gene, Tokyo, Japan) according to the manufacturer's instructions. Reverse transcription and PCR were carried out sequentially using the protocol of the QIAGEN OneStep RT-PCR Kit (QIAGEN, Hilden, Germany). Reverse transcription polymerase chain reaction (RT-PCR) was performed on cDNA using the following primers (Invitrogen) and conditions. Oct3/4 (685 bp): forward, 5′-CCGCTGTATGAGTTCTGTGG-3′, reverse, 5′-CATAGTCGCTGCTTGATTGC-3′, 94°C for 1 min, 57°C for 1 min, 72°C for 1 min, for 40 cycles. Nanog (216 bp): forward, 5′-CAGAAGGCCTCAGCACCTAC-3′, reverse, 5′-CTGTTCCAGGCCTGATTGTT-3′, 94°C for 1 min, 56°C for 1 min, 72°C for 1 min, for 40 cycles. Sox2 (410 bp): forward, 5′-AGTCTCCAAGCGACGAAAAA-3′, reverse, 5′-GGAAAGTTGGGATCGAACAA-3′, 94°C for 1 min, 55°C for 1 min, 72°C for 1 min, for 40 cycles. C-myc (273 bp): forward, 5′-GATTCTCTGTCTCCTCGACGGAG-3′, reverse 5′-GCGCTGCGTAGTTCTGCTGAT GTG-3′, 94°C for 1 min, 55°C for 1 min, 72°C for 1 min, for 40 cycles. Klf4 (332 bp): forward, 5′GTTTTGAGGAGGTGCTGAG-3′, reverse 5′-CAGTCACAGTGGTAAGGTTT-3′, 94°C for 1 min, 55°C for 1 min, 72°C for 1 min, for 40 cycles. GAPDH (411 bp): forward, 5′-CAAGAAGGTGGTGAAGCAGG-3′, reverse 5′-ATGGTACATGACAAGGT GCG-3′, 94°C for 1 min, 55–57°C for 1 min, 72°C for 1 min, for 40 cycles. PCR products were separated in 2% agarose gels followed by ethidium bromide staining and visualization on a UV illuminator.
Immunohistochemistry
Immunocytochemistry was performed to examine the expression of Nanog, Oct3/4, Sox2, Klf4, and c-myc in fresh, sphere, and adherent WJCs. To detect the expression of embryonic stem cell (ESC) markers, cultured WJCs, detached from dishes with trypsin, or fresh specimens were made with a Cytospin. Cells were fixed in cold acetone for 2 min and air-dried. After washing with PBS three times for 5 min each time, the cells were fixed in PBS 4% paraformaldehyde with 0.5% CaCl2 for 20 min. Further washing was done followed by permeation with 0.1% NP-40 in 0.01 M PBS. Then the cells were blocked using 10% nonimmune mouse, goat, or rabbit serum (Nichirei, Tokyo, Japan) for 1 h at room temperature and incubated with the primary antibodies, anti-Oct3/4, anti-Nanog, anti-Klf4, anti-c-myc (Santa Cruz Biotechnology, Inc. California, USA), anti-Sox2 (R&D System, Inc. Minneapolis, USA), and vimentin (Dako, Glostrup, Denmark) at a dilution 1:200 in PBS overnight at 4°C. After washing, the cells were incubated with the secondary antibodies: biotin-conjugated anti-mouse, -goat, or -rabbit immunoglobulin G (IgG; Nichirei) for 1 h at room temperature. After washing with PBS, they were incubated with fluorescein isothiocyanate (FITC)-conjugated streptavidin (1:100, Dako Cytomation, Denmark) in PBS for 30 min at room temperature. Counterstaining for the nuclei was performed with a Hoechst 33258 (Dojndo lab, Japan) and examined by fluorescence microscopy (Leika DM RBE-Welzlar, Germany).
Flow cytometry
WJCs were analyzed by flow cytometry in five different samples at passage 3. They were trypsininzed and washed in 0.2% EDTA/PBS. The cells were aliquoted at a concentration of 1.0×106 cells/mL and stained for 20 min at 4°C. Cell-surface antigen phenotyping was performed by using mouse monoclonal antibodies according to the manufacturer's dilution recommendations. The following monoclonal antibodies were used: FITC-conjugated CD14 (FITC-CD14), FITC-CD34, phycoerythrin (PE)-CD 90 (PE-CD 90), FITC-CD 105, FITC-HLA-DR (Beckman Coulter, France), PE-CD 73, and PE-CD45 (BD Bioscience Pharmingen, Vienna, Austria). A negative control was prepared by incubation with mouse IgG1-isotyped antibodies conjugated to FITC or PE (Beckman Coulter, France). Labeled cells were washed twice and resuspended in 0.2% EDTA/PBS. Analysis was performed with a flow cytometer (EPICS Altra, Beckman Coulter). A total of 104 events were acquired in a histogram figure with Cell Quest software (BD Bioscience), and the data were analyzed with WinCyte acquisition software.
Differentiation of cells
Inductions of adipogenic, osteogenic, and chondrogenic differentiation were carried out using the protocol of the Lonza differentiation Kit (Walkersville, MD, USA). Multilineage differentiation to adipocyte, chondrocyte, and osteocyte lineages was performed in five different samples.
For analysis of adipogenic differentiation, WJCs (4.2×104 cells/cm2) were seeded on each well of two-well chamber slides and cultured in the proliferative medium for 5–10 days to reach 100% confluence. Adipogenic differentiative medium was then added and changed twice a week. Incubation of the cells in induction and then maintenance media (step 1 and 2) was repeated four to five times for 4 weeks. As a negative control, the same numbers of cells were cultured in the proliferative medium. Finally cells were fixed with 10% formaldehyde for 30 min and stained with fresh Oil Red O for 10 min at room temperature. Lipid droplets were visualized and photographed.
For induction of osteogenic differentiation, WJCs (4.2×104 cells/cm2) were cultured on each well of two-well chamber slides. After 24 h, the medium was changed with the osteogenic differentiation medium. The medium was replaced twice a week. After 4 weeks in differentiation medium, cells were stained with Alizarin Red S solution. Briefly, cell layers were fixed with 10% formalin for 30 min and incubated with 2% Alizarin Red S solution for 15 min. The layer was washed with distilled water. As a negative control, the same numbers of cells were cultured in the proliferative medium.
A pellet culture system was used for chondrogenic differentiation. A total of 2.5×105 cells were centrifuged in a 15-mL polypropylene tube at 500×g for 5 min, and the pellet was resuspended in 1–2 mL of chondrogenic noninduced medium. Without disturbing the pellet, cells were resuspended in 2 mL of chondrogenic-induced medium. Medium changes were made every 2–3 days. After 4 weeks, cell pellets were fixed with 4% paraformaldehyde at room temperature. As a negative control, the same numbers of cells were cultured in the proliferative medium. The production of mucopolysaccharide, an indicator of chondrogenic differentiation, was measured by staining thin sections of the pellets with Toluidine Blue.
A total of 2.5×105 WJCs were washed at 150×g for 5 min at room temperature, and the supernatant was discarded. After resuspension in 1 mL of incomplete chondrogenic medium per 7.5×105 cells, the cells were centrifuged again at 150×g for 5 min, and the medium was discarded. WJCs were resuspended in complete chondrogenic medium to a concentration of 5.0×105 cells/mL, and aliquots of 0.5 mL (2.5×105 cells) of the cell suspension were made in 15-mL polypropylene culture tubes. These were centrifuged at 150×g for 5 min at room temperature. Loosening the caps of the tubes one-half turn allowed gas exchange, and the samples were then incubated at 37°C in a humidified atmosphere of 5% CO2. The cell pellets were fed every 2–3 days by completely replacing the pellets when aspirating the medium and attaching a sterile 1- to 200-μL pipette tip of freshly prepared complete chondrogenic medium to each tube. The total number of pellet cultures was calculated.
Floating culture protocol
WJCs after harvesting were suspended and disseminated on ultra-low-attachment dishes in DMEM low glucose supplemented with 2 mM
Results
Isolated WJCs were expanded in culture and adhered to growth plates. They displayed a heterogeneous morphology with many spindle-shaped cells, and proliferated (Fig. 1A). Also, WJCs formed colonies after passage 2 (Fig. 1B). All WJCs expressed vimentin (green) at passage 2 (Fig. 1C). Thus, WJCs have the characteristics of mesenchymal cells.
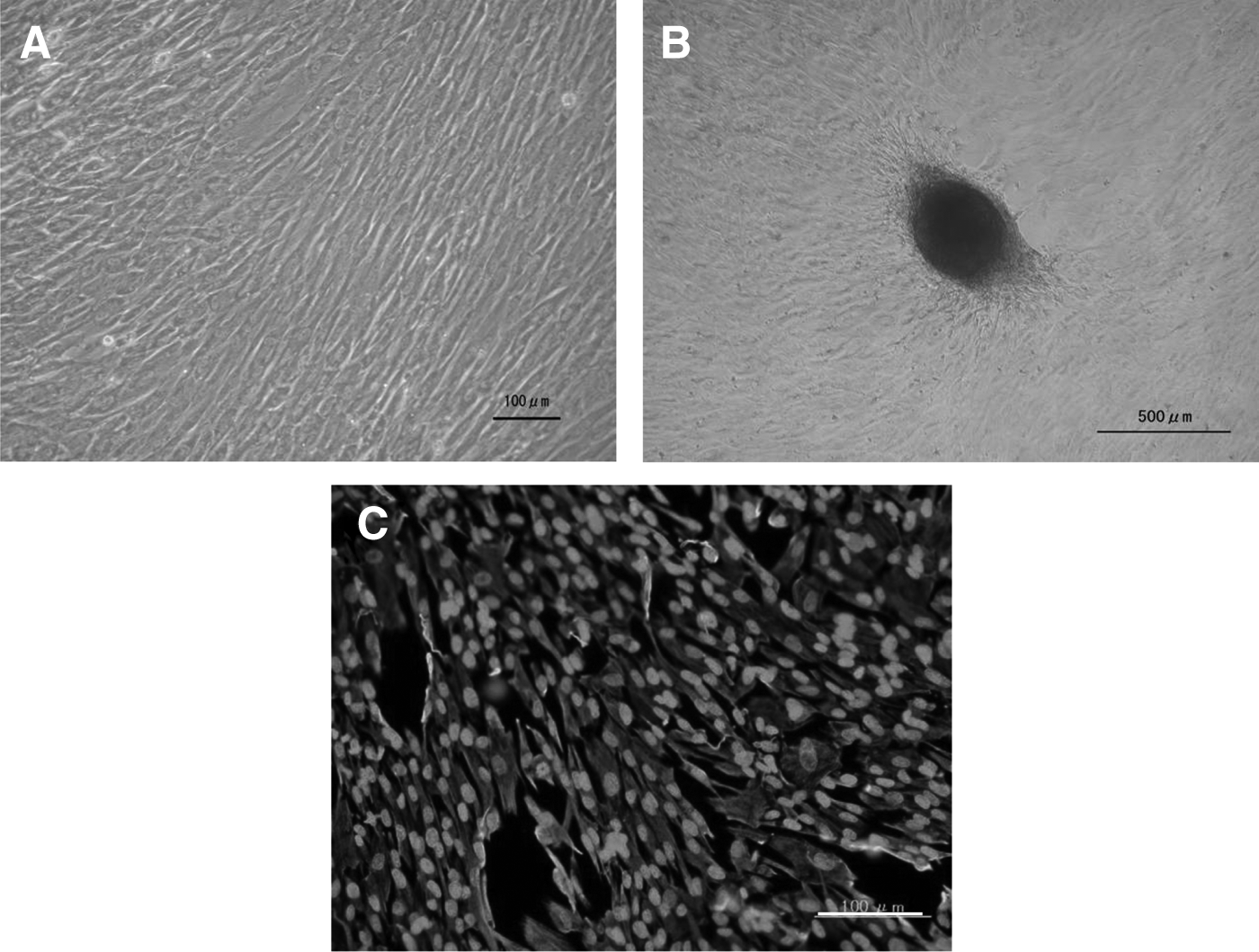
WJCs adhered to plastic plates and formed fibroblast colony-forming units. (
The expression of transcription factors such as Nanog, Oct3/4, Sox2, Klf4, and c-myc was analyzed in fresh WJCs with RT-PCR (Fig. 2A). Fresh WJCs expressed Nanog (Fig. 2B), Oct3/4 (Fig. 2C), and Sox2 (Fig. 2D) by immunofluorescence analysis. The expression of the transcription factors in WJCs at the level of RNA and protein was detected.
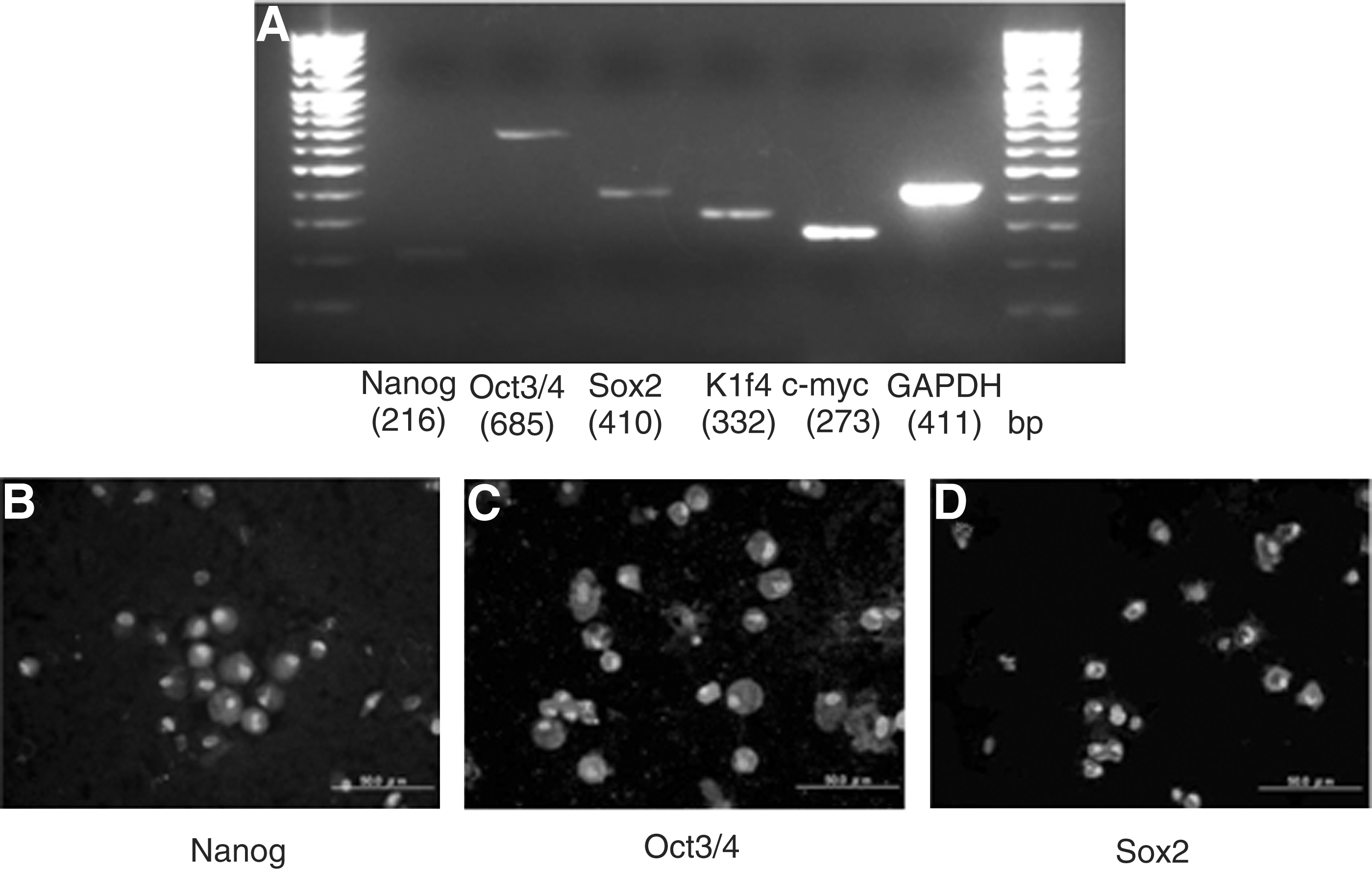
Stem cell markers are present in fresh WJCs. (
Furthermore, WJCs were analyzed by flow cytometry. They predominantly expressed the MSC markers CD73, CD90, and CD105, whereas the hematopoietic lineage markers, such as CD14, CD34, CD45, and HLA-DR, were not expressed. Thus, WJCs expressed MSC markers and lacked expression of hematopoietic stem cell markers (Fig. 3).
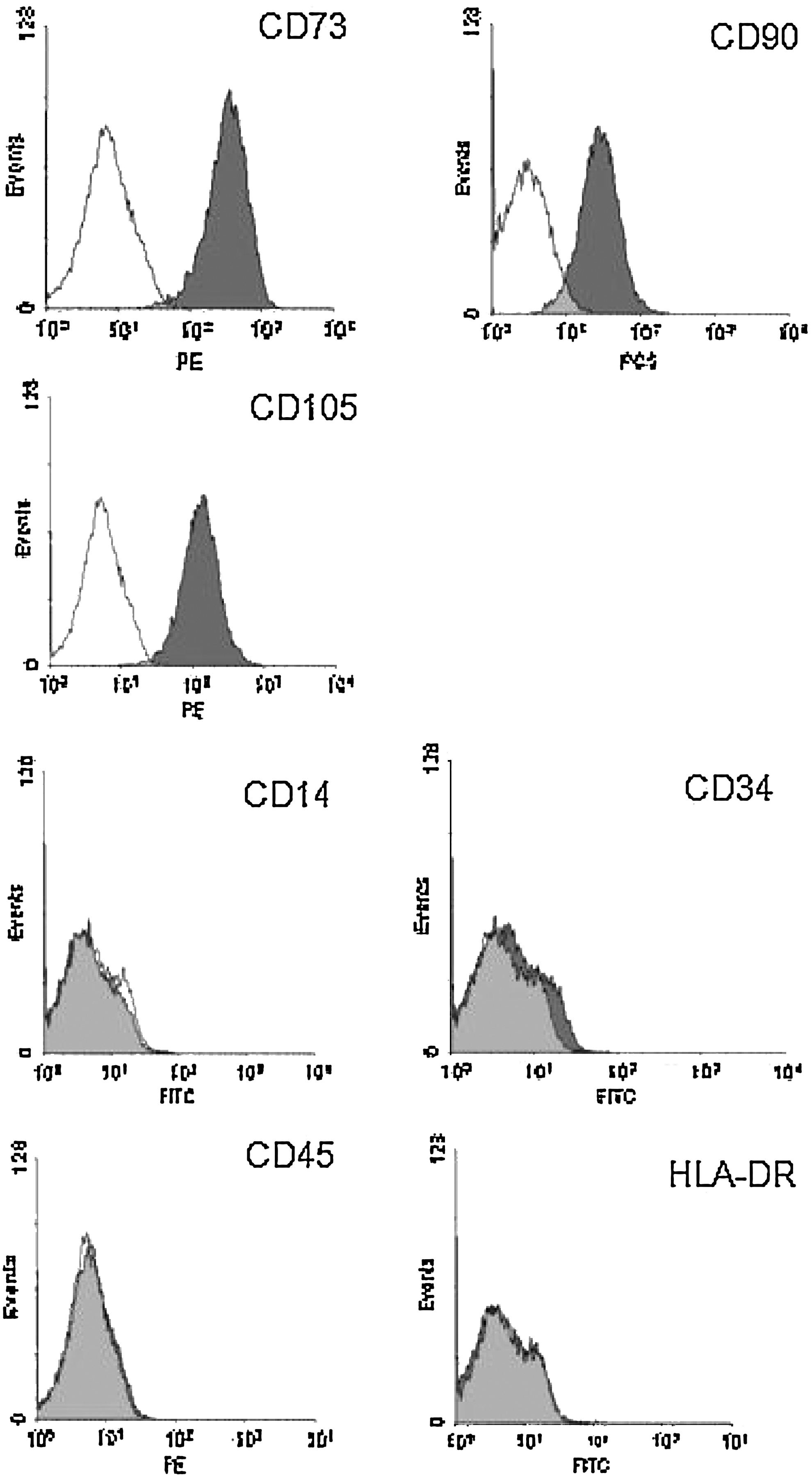
Markers expression of WJCs by flow-cytometric analysis. WJCs predominantly expressed MSC markers CD73, CD90, and CD105 (upper two rows), but did not express the hematopoietic lineage markers CD14, CD34, CD45, and HLA-DR (lower two rows).
To investigate the differentiation potency, WJCs at passage 2 were cultured with osteogenic, adipogenic, and chondrogenic induction medium. The osteogenic differentiation was indicated by calcium deposition, as demonstrated by Alizarin Red staining (red) in induced cells. Accumulation of lipid droplets was observed by staining with Oil Red O in treated cells. Chondrogenic differentiation was indicated by Toluidine Blue staining in treated cells. WJCs could differentiate to multiple connective tissue lineages (Fig. 4).
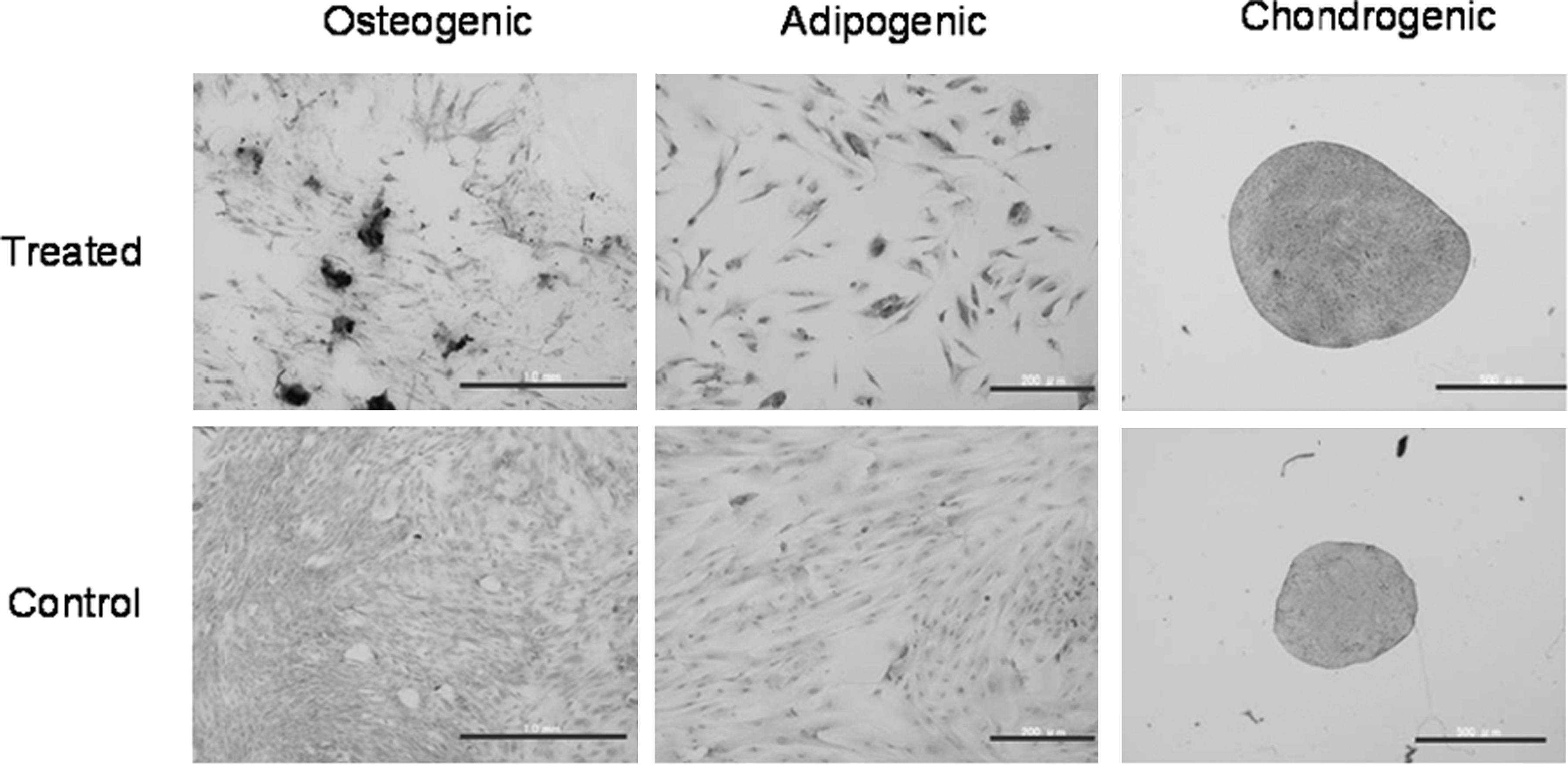
WJCs differentiated to multiple connective tissue lineages. Osteogenic differentiation was indicated by calcium deposition as demonstrated by Alizarin Red staining in treated cells (left panel). Scale bar, 1.0 mm. Accumulation of lipid droplets was demonstrated by staining with Oil Red O in treated cells (middle panel). Scale bar, 200 μm. Chondrogenic differentiation was indicated by Toluidine Blue staining in treated cells (right panel). Scale bar, 500 μm. Upper-row photos indicate the differentiated cells treated with the specific differentiation media, whereas lower-row photos indicate the negative control cells treated with the normal proliferative medium.
Thus, we demonstrated that WJCs satisfy the criteria for defining multipotent MSCs, but the stemness of immature cells disappeared during cultivation. Three weeks after being expanded on culture plates, WJCs showed decreased expression of stem cell markers like Oct3/4, Sox2, Klf4, and c-myc that have been induced in induced pluripotent stem cells (iPSCs). We attempted a floating culture to maintain the stemness on ultra-low-attachment dishes. Seeded WJCs formed spheres and were cultured for 21 days (Fig. 5A). After aspirating with a pipette, WJCs spheres were put on slides by the cytospin method. With immunostaining, the spheres of WJCs expressed stem cell marker proteins, such as Oct3/4, Sox2, Klf4, and c-myc, whereas cultured adherent WJCs did not (Fig. 5B). Therefore, WJCs could maintain their stemness in a floating culture.
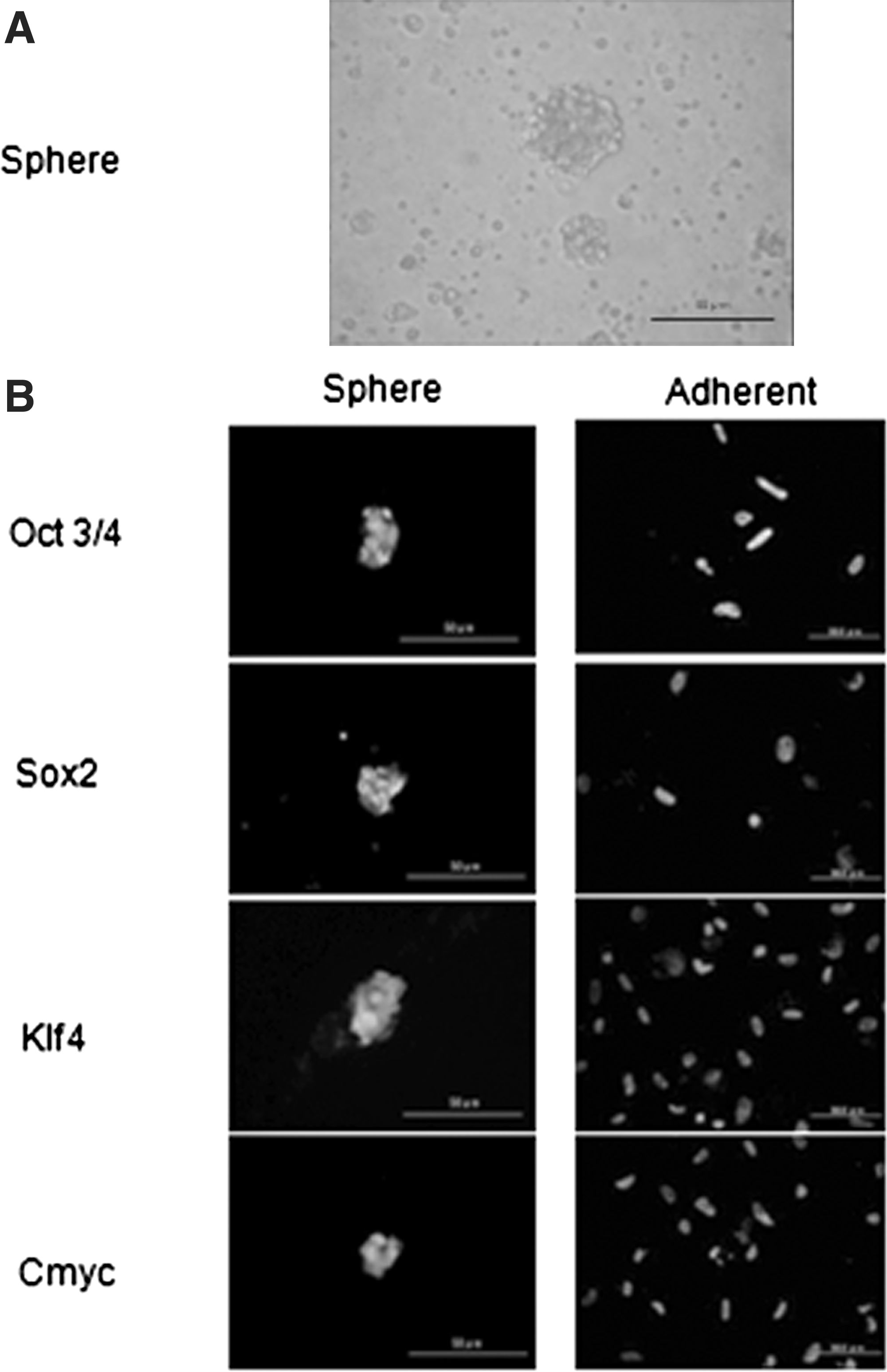
Spheres of WJCs express stem cell marker proteins, but adherent cells do not. (
Discussion
In this study, we proved the existence of MSCs in Wharton's jelly. WJCs were plastic-adherent in standard culture conditions and formed fibroblast colony-forming units. In flow-cytometric analysis, WJCs expressed CD73, CD90, and CD105, but not hematopoietic markers, such as CD14, CD34, CD45, and HLA-DR. Furthermore, WJCs differentiated in vitro into osteoblasts, adipocytes, and chondroblasts.
Carlin et al. reported the expression of the early transcription factors Oct-4, Sox-2, and Nanog by porcine umbilical cord matrix cells (Carlin et al., 2006). Stromal cells of the umbilical cord were also found to express low levels of these transcription factors at the mRNA level. These transcription factors are expressed at high levels in ESCs (Takahashi et al., 2007). These factors regulate the expression of other genes during development and are found at high levels in the pluripotent cells of the inner cell mass. The downregulation of these transcription factors correlates with the loss of pluripotency and self-renewal and the beginning of subsequent differentiation steps.
Previous reports isolated and characterized human and animal Wharton's jelly MSCs (Garzon et al., 2012; Iacono et al., 2012; Ruan et al., 2012). Salehinejad et al. (2012) reported that the collagenase/trypsin method of cell isolation yields a high cell density, and the isolated cells expressed a high rate of pluripotent cell marker Oct-4, whereas the explant method of cell isolation resulted in a high cell proliferation rate and activity (Salehinejad et al., 2012).
Umbilical cord stromal cells have the potential to differentiate into adipocyte, chondrocyte, osteocyte, cardiomyocyte, skeletal myocyte, neuronal precursor, dopaminergic neuron, and endothelial cell lineages (Conconi et al., 2006; Karahuseyinoglu et al., 2007; Wang et al., 2004; Weiss et al., 2006; Wu et al., 2007). We also found that fresh WJCs expressed ESC-specific markers, but over time these markers decreased and disappeared. In flow cytometric analysis, expression of CD90 and CD105 decreased by passaging of the cells (data not shown). Therefore, to maintain the stemness of stem cells, we cultured WJCs with a floating culture method. We proved that it was possible to maintain the stemness of WJCs with immunostaining. It was shown that the floating culture method maintains the stemness of immortalized human amniotic MSCs (Teng et al., Cell Transplantation, 2012, in press).
Stem cells isolated from adults or developing embryos (ESCs) are currently thought to be a source of cells for regenerative medicine. However, despite their therapeutic potential, both adult and ESCs present a number of challenges associated with their clinical application and are thus not in general use yet (Smith et al., 2006). For example, although adult stem cells can be directly isolated from the patient and are therefore immunologically compatible with this patient, they are generally hard to isolate and grow in culture; and moreover, transplantation of a sufficient number of cells to adult tissue needs a large-scale cell supply.
Recently, the multipotent differentiation ability of placenta- or amnion-derived cells has been reported, and these cells have attracted a lot of attention as a cell source for cell transplantation therapy (Chan et al., 2006; Fukuchi et al., 2004; In 't Anker et al., 2004; Soncini et al., 2007; Tamagawa et al., 2004). The amnion-derived cells have considerable advantages: They can differentiate into all three germ layers, they have low immunogenicity and antiinflammatory functions, and they do not require the sacrifice of human embryos for their isolation, thus avoiding the current controversies associated with the use of human ESCs.
Bone marrow stem cells or umbilical cord blood cells are mainly used in pediatric transplantation treatments for leukemia or other immunologic diseases (Jaing et al., 2009). However, by using MSCs from Wharton's jelly, diseases such as congenital bone defects or cartilage dysplasia are potentially therapeutic objectives. Although rare, various types of diseases exist, such as achondroplasia, atelosteogenesis, or osteolyses. At present, only supportive care is available for congenital bone and cartilage diseases, and major interventions cannot be done. Regenerative medicine has potential to offer treatments for such inherent diseases. However, such diseases require time before a diagnosis is reached. During the time required for examination, stemness of WJCs would be lost. To compensate for this shortcoming, it will be beneficial to use this floating culture in the future.
iPS cells can be created from skin (Takahashi et al., 2007), but it is very invasive to harvest skin cells from a child, especially a newborn, and little blood can be collected. Appendages of the fetus, such as the umbilical cord and the amnion, are the best choices to obtain sufficient cells without invasive treatment for newborns. We hope that this method will help to relieve bone dysplasia.
In conclusion, WJCs expressed the ESC markers Nanog, Oct3/4, and Sox2. WJCs predominantly expressed the MSC markers CD73, CD90, and CD105, but not hematopoietic lineage markers, such as CD14, CD34, CD45, and HLA-DR. In the floating culture, WJCs could maintain stemness for a while. After induction of differentiation, WJCs after two passages could differentiate to osteogenic, chondrogenic, and adipogenic lineages. Thus, WJCs satisfy the criteria for MSCs.
Given that extraction of the umbilical cord is not invasive, and the umbilical cord can be argued to be without ethical and technical issues, it is suggested that WJCs after maintaining the stemness characteristics have the potential to contribute to medical treatments for patients with congenital skeletal and cartilage disorders, even newborns.
Footnotes
Author Disclosure Statement
The authors declare no conflicts of interest exist.