
Abstract
Abstract
Nodal/activin signaling is indispensable for embryonic development. We examined what activin does to the embryoid bodies (EBs) produced from mouse embryonic stem cells (mESCs) expressing an epiblast marker. The EBs were produced by culturing mESCs by the hanging drop method for 24 hours. The resulting EBs were transferred onto gelatin-coated dishes and allowed to further differentiate. The 24-hour EBs showed a stronger expression of fibroblast growth factor (FGF)5 and Brachyury (specific to the epiblast) in comparison with mESCs. Treating the transferred EBs with activin A maintained transcript levels of FGF5 and Oct4, while inhibiting definitive endoderm differentiation. The activin A treatment reversed the endoderm differentiation induced by retinoic acid (RA), while the inhibition of nodal/activin signaling promoted RA-induced endoderm differentiation. Inhibition of nodal/activin signaling in EBs, including epiblast-like cells, promotes differentiation into the endoderm, facilitating the transition from the pluripotent state to specification of the endoderm.
Introduction
D
Pluripotent stem cells are defined by their ability to differentiate into the three germ layers and can expand indefinitely in vitro. Various types of pluripotent stem cells of different origin with distinct characteristics have been classified into two categories: the naïve and primed type. The former includes mouse embryonic stem cells (mESCs) and mouse induced pluripotent stem cells (miPSCs), whereas the latter includes mouse epiblast stem cells (mEpiSCs) and human embryonic stem cells (hESCs) (Nichols and Smith, 2009).
mESCs were first isolated in 1981 and derived from the ICM of an embryo at the preimplantation blastocyst stage (Evans and Kaufman, 1981; Martin, 1981). The standard culture of mESCs requires a combination of a cytokine (leukemia inhibitory factor [LIF] to activate STAT3) and either serum or bone morphogenetic protein (to induce inhibitor of differentiation proteins) (Nichols and Smith, 2011; Ying et al., 2008). Nevertheless, the extrinsic stimuli are dispensable for the formation, propagation, and pluripotency of mESCs. Self-renewal is enabled by the elimination of differentiation-inducing signaling by mitogen-activated protein kinase. Additional inhibition of glycogen synthase kinase 3 consolidates the biosynthetic capacity and suppresses residual differentiation.
On the other hand, mEpiSCs are self-renewing multipotent cells that are derived from the epiblast of postimplantation embryos of E5.5–E6.5 (Brons et al., 2007; Tesar et al., 2007). They are maintained in vitro through cultivation in the presence of activin A and fibroblast growth factor (FGF)2. On the other hand, mEpiSCs cannot be propagated efficiently from a single cell and do not readily contribute to formation of chimeras. mEpiSCs share many characteristics with hESCs. mEpiSCs are transcriptionally and epigenetically distinct from mESCs. mEpiSCs are characterized by expression of FGF5 and brachyury, whereas the hallmark of mESCs is expression of Rex1 and Krüppel-like factor (Klf)4 and the absence of X chromosome inactivation (Brons et al., 2007; Tesar et al., 2007). mESCs are believed to be capable of differentiation into mEpiSCs because the origin of mESCs is a little earlier than that of mEpiSCs. The mESCs can readily turn into mEpiSCs in response to activin A and FGF2, whereas mEpiSCs cannot turn into mESCs. Therefore, mEpiSCs can be reprogrammed to acquire the defining characteristics of embryonic stem (ES) cells, including germline competence, after transfection with a single pluripotent factor, such as Klf4 (Guo et al., 2009; Hall et al., 2009).
Differentiation of mESCs can be induced in a variety of ways, but the most common method involves formation of cell aggregates referred to as embryoid bodies (EBs). In developmental experiments, formation of EBs is very important because it allows mESCs to differentiate into specific cell types. Molecular and cellular morphogenic signals and events within EBs recapitulate many characteristics of the developing embryo, including differentiation into cells of all three germ layers—the endoderm, mesoderm, and ectoderm—similar to gastrulation in vivo (Carpenedo et al., 2007; Itskovitz-Eldor et al., 2000; Keller, 1995). The hanging drop (HD) method is used to produce EBs of 200–500 cells dispersed in ∼20-μL drops suspended from the lid of a Petri dish. This method allows for production of a more homogeneous population of EBs than does suspension culture, which is a method that is used to induce cells to aggregate spontaneously into spheroids through simple seeding of the cells in a medium at a density of 104–106/mL on a nonadherent surface (Keller, 1995).
Activin/nodal signaling maintains the undifferentiated state in mEpiSCs, whereas differentiation into the mesendoderm is induced when this signaling pathway is activated in mESCs, whose origin is considered to be earlier than that of mEpiSCs (Brons et al., 2007; James et al., 2005; Tesar et al., 2007; Vallier et al., 2005). Nevertheless, a few studies have shown that activin treatment fails to induce effective differentiation of mESCs into the endoderm (Nakanishi et al., 2009; Sherwood et al., 2007). Thus, in a narrow developmental time window, there is a remarkable change in how nodal signals are interpreted intracellularly.
Expression of FGF5 (characteristic of mEpiSCs) is detectable in EBs as early as the first day in suspension culture of mESCs (Leahy et al., 1999). Compared with the ICM, the primitive ectoderm shows upregulation of FGF5 and downregulation of Rex1 (Haub and Goldfarb, 1991; Hébert et al., 1991; Rogers et al., 1991). Furthermore, gastrulation brain homeobox 2 (Gbx2) and orthodenticle homeobox 2 (otx2) are reported to be induced in the ICM or mESCs and the epiblast or mEpiSCs, respectively (Brons et al., 2007; ten Berge et al., 2011; Tesar et al., 2007). The formation of EBs in vitro can be used to study differentiation and gene expression during early development (Dvash et al., 2004). Therefore, we were interested in what activin A does to the cells expressing the genetic characteristics of the epiblast; this is one of the earliest commitment steps in embryogenesis. We produced EBs in which early embryonic cell lineages develop and then analyzed how activin affects the EBs expressing an epiblast marker such as FGF5. Activin inhibited the time-dependent downregulation of the mRNA expression pattern that is characteristic of the epiblast and the pluripotent state. Inhibition of nodal/activin signaling in EBs, including epiblast-like cells, promoted differentiation into the endoderm, facilitating the transition from the pluripotent state to specification of the endoderm.
Materials and Methods
Cell lines and reagents
mESCs, that is, ES D3 cells (ATCC, Manassas, VA), and miPSCs, that is, iPS-MEF-Ng-20D-17-expressing green fluorescent protein under the NANOG promoter (NANOG iPS cells; RIKEN BioResource Center, Tukuba, Japan), were cultured in tissue culture dishes precoated with 0.1% gelatin, with mitomycin C (Kyowa Hakko Kirin, Inc., Tokyo, Japan)-treated mouse embryonic fibroblasts (MEFs) in the ES medium. The ES medium comprised Dulbecco's modified Eagle's medium (DMEM; Invitrogen, Carlsbad, CA) supplemented with 15% fetal bovine serum (FBS; HyClone, Logan, UT), 1 mM glutamine, 1% nonessential amino acids, mercaptoethanol (5 μL/1000 mL), and 1% penicillin/streptomycin. StemSure LIF (Wako Pure Chemical Industries, Ltd., Osaka, Japan), containing LIF, was added at the concentration of 1000 U/mL for maintenance of the undifferentiated state of the mESCs and miPSCs.
Formation of EBs and differentiation of the mouse cells
Before differentiation, the mESCs and miPSCs were maintained on gelatin-coated dishes without MEFs in the ES medium, including StemSure LIF, for three passages to deplete MEFs. Then, the cells were dissociated with 0.05% trypsin–EDTA and suspended in the ES medium in the absence of StemSure LIF. To produce EBs, we cultured the dissociated cells by the HD method. The drops containing 500 cells per 20 μL of the ES medium without StemSure LIF were placed on the inside of the lid; the latter was inverted gently and placed on a 100-mm Petri dish filled with phosphate-buffered saline (PBS). Then, the whole construct was maintained at 37°C in a humidified atmosphere containing 5% of CO2. After 24 hours, the resulting EBs were transferred onto 35-mm gelatin-coated dishes (200 EBs/dish) and allowed to attach to the dishes to obtain outgrowth culture in the differentiation medium comprising DMEM supplemented with 0.1% bovine serum albumin (Nacalai Tesque, Inc., Kyoto, Japan), insulin-transferrin-selenium (Invitrogen), 1 mM glutamine, 1% nonessential amino acids, mercaptoethanol (5 μL/1000 mL), and 1% penicillin/streptomycin. The differentiation was induced by retinoic acid (RA; Sigma–Aldrich, St. Louis, MO), activin A (R&D Systems, Minneapolis, MN), and SB431542 (Wako Pure Chemical Industries, Ltd.), which is a specific inhibitor of TGF-β superfamily type I activin receptor-like kinase (ALK) receptors, ALK4, ALK5, and ALK7, and consequently reduces phosphorylation of SMAD2 or SMAD3 under serum-free conditions.
Alkaline phosphatase staining
This staining was performed by means of the Leukocyte Alkaline Phosphatase (ALP) Kit (Sigma–Aldrich) on the EBs that differentiated on gelatin-coated dishes.
RNA extraction, reverse transcription, and real-time quantitative reverse transcription–polymerase chain reaction
Total RNA was isolated with ISOGEN II (Nippon Gene Co., Ltd., Tokyo, Japan). Reverse transcription (RT) of messenger RNA was performed by means of the VersoTM cDNA Kit (Thermo Fisher Scientific, Inc., Waltham, MA). For each sample, cDNA was prepared from 0.5 μg of total RNA.
Real-time quantitative reverse transcription–polymerase chain reaction (RT-PCR) analysis was carried out on a StepOnePlus Real-Time PCR system (Applied Biosystems, Foster, CA) with the SYBR Premix Ex Taq TM II (Takara Bio, Inc., Shiga, Japan). The sequences of the primers are listed in Table 1. The cDNA was amplified under the following cycling conditions: 95°C for 10 minutes, followed by 40 cycles at 95°C for 30 seconds and 60°C for 1 minute.
Immunofluorescence staining
The cells were fixed with 4% paraformaldehyde for 30 minutes at room temperature. After washing thrice with PBS, the cells were immersed in 0.5% Triton X-100/PBS for 3 minutes at room temperature, followed by a 30-minute incubation with Blocking one (Nacalai Tesque, Inc.) at room temperature. The cells were incubated in the solution, including primary antibodies to α-fetoprotein (AFP) (Cat. No. sc8108, 1:50 dilution; Santa Cruz Biotechnology, Inc., Dallas, TX), transthyretin (TTR) (Cat. No. ab78548, 1:200; Abcam, Cambridge, United Kingdom), Albumin (Cat. No. A90-134A, 1:200; Bethyl Laboratories, Inc., Montgomery, TX), Foxa2 (Cat. No. D56D6, 1:200; Cell Signaling Technology, Inc., Tokyo, Japan), CXCR4 (Cat. No. ab2074, 1:50; Abcam), Pdx1 (Cat. No. MAB2419, 1:50; R&D Systems, Inc.), and Proinsulin (cat. No. MAB13361, 1:50; R&D Systems, Inc.), for 60 minutes at room temperature, followed by washing thrice with PBS, after which they were incubated in the solution, including secondary antibodies (anti-rabbit IgG antibody, Cat. No. ab6717, Abcam; anti-goat IgG antibody, Cat. No. ab6881, Abcam; anti-mouse IgG antibody, Alexa Fluor 488, Thermo Fisher Scientific, Inc.), for 60 minutes at room temperature and washed thrice with PBS. Nuclei were counterstained with Hoechst-33342.
Flow cytometry
For detection of FGF5, Foxa2, CXCR4, and C-peptide, the cells were washed twice in PBS and then incubated with 0.5% trypsin-EDTA for 10 minutes at 37°C. The cells were dissociated by gentle pipetting and resuspended at ∼105/μL in PBS containing 2% FBS. After that the cells were incubated for 10 minutes at room temperature with 3.6% formaldehyde, followed by a 15-minute incubation with methanol at 4°C. The resuspended cells were washed twice with PBS containing 2% FBS (by centrifugation), and then they were incubated for 30 minutes at room temperature with primary antibodies to FGF5 (Cat. No. sc-7914, 1:100 dilution; Santa Cruz Biotechnology, Inc.), Brachyury (Cat. No. ab20680, 1:50; Abcam), or C-peptide (CST #45935, 1:100; Cell Signaling Technology, Inc., Danvers, MA). Then, after two washes with PBS containing 2% FBS, the cells were incubated for 30 minutes at room temperature with secondary antibodies (anti-rabbit IgG antibody, Cat. No. ab6717; Abcam). The labeled cells were analyzed on an FACSCalibur instrument (BD Biosciences, San Jose, CA). At least 10,000 events were recorded during each run of flow cytometry.
Statistical analysis
All data are expressed as mean ± standard error. Differences between groups were evaluated by one-way analysis of variance or the Kruskal–Wallis test. A difference with the p-value <0.05 was considered significant.
Results
Induction of the early embryonic lineage, including the epiblast by the 24-hour HD method
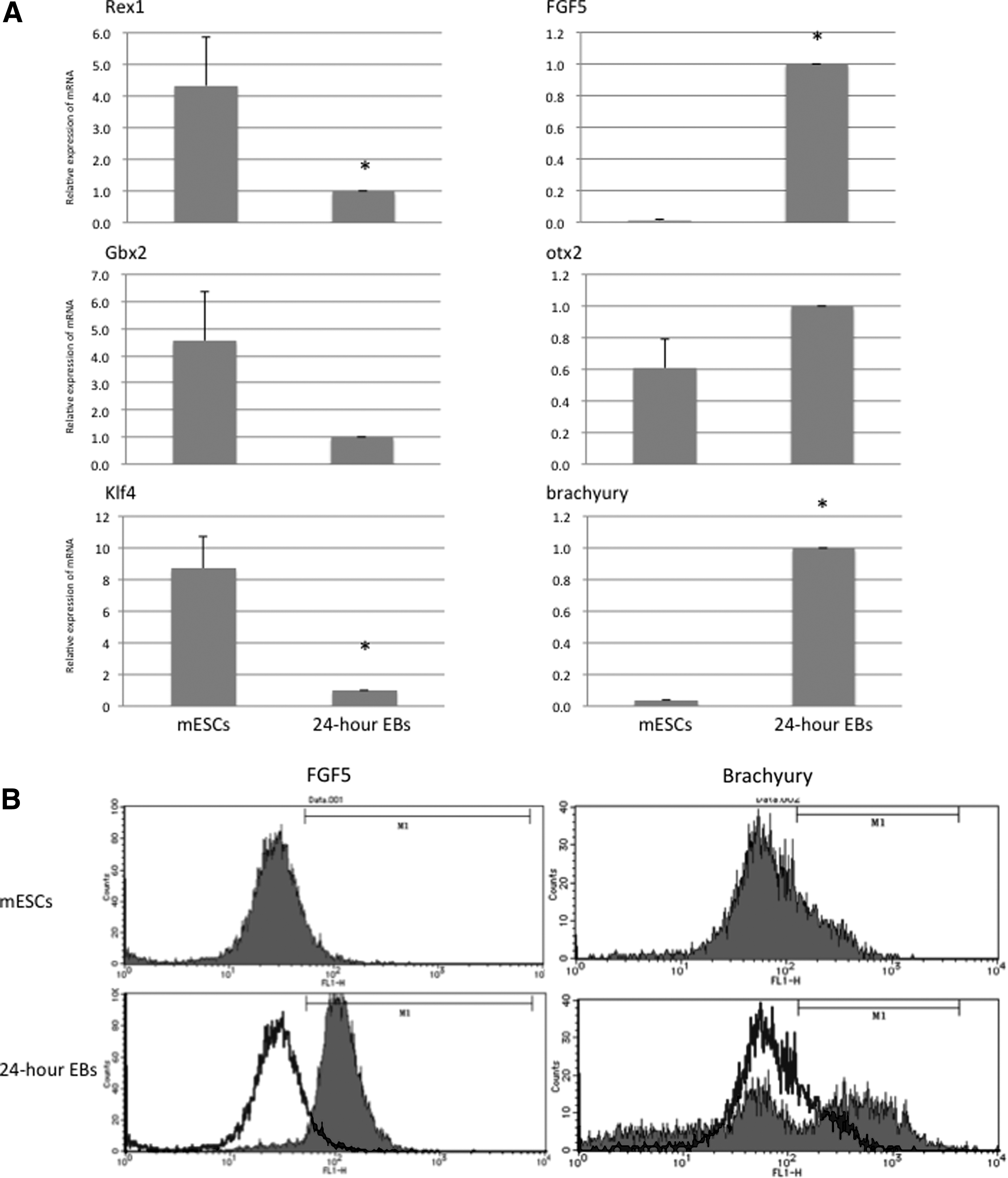
The EBs produced from ES D3 cells by the HD method after 24 hours were prepared for real-time quantitative RT-PCR and flow cytometric analysis. The transcript levels of genes Rex1 and Klf4 in 24-hour EBs showed a significant decrease in comparison with mESCs, although the decrease in Gbx2 mRNA expression was not statistically significant. On the other hand, 24-hour EBs showed a significantly higher expression of FGF5 and brachyury mRNA than did mESCs, although the increase in otx2 mRNA expression was not statistically significant (Fig. 1A). Flow cytometric analysis confirmed the higher expression of FGF5 and Brachyury in 24-hour EBs than in mESCs (Fig. 1B).
The effect of activin A treatment on the transferred 24-hour EBs
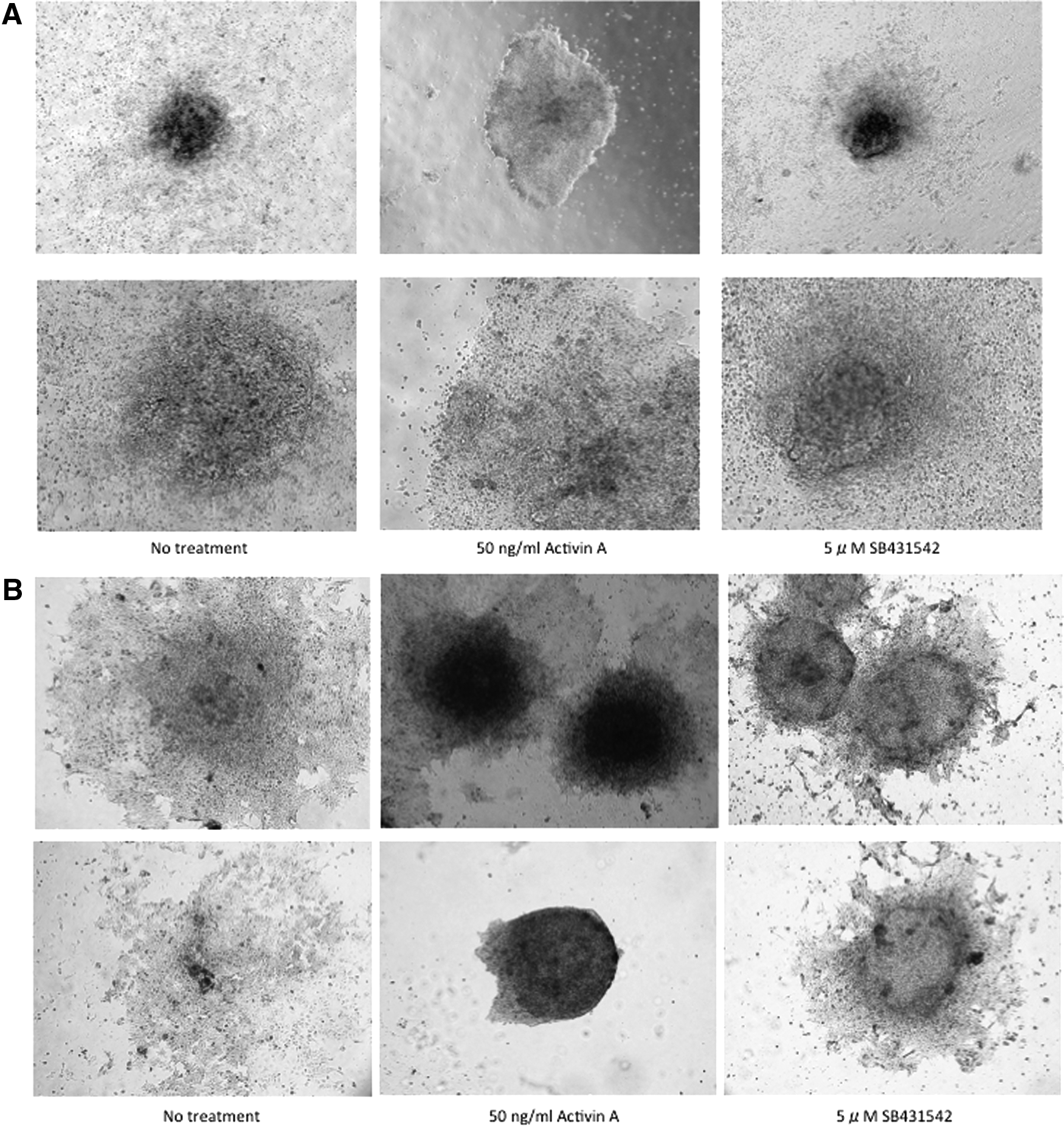
The 24-hour EBs that were produced from ES D3 cells were transferred onto 35-mm gelatin-coated dishes and allowed to further differentiate under serum-free conditions. After 6 days, the EBs in the no treatment group showed vigorous outgrowth similar to those treated with SB431542 (5 μM); activin A treatment (50 ng/mL) kept the transferred EBs rounder than no treatment and SB431542 treatment. The EBs in both no treatment and SB431542 treatment groups showed more modest ALP staining than in the activin A treatment group (Fig. 2).
Expression of epiblast markers, such as FGF5 and nodal, showed a time-dependent decrease when the transferred EBs were allowed to outgrow under serum-free conditions for 8 more days. The transferred EBs were incubated with activin A (5 or 50 ng/mL) or SB431542 (5 μM) for the first 4 days only. The transcript levels of FGF5 and nodal in the EBs were stable during the first 4-day activin A treatment (5 or 50 ng/mL) and declined after that. The activin A treatment brought about statistically significant differences with the other groups. There were no significant differences in the mRNA expression of epiblast markers between 4-day SB431542 treatment and no treatment groups. In particular, because Nodal mRNA expression showed no difference between no treatment and SB431542 treatment, the lines showing mRNA expression time course in both groups could not be distinguished in Figure 3. Oct4 and NANOG are transcription factors necessary for maintenance of the pluripotency and self-renewal of mESCs. Their mRNA expression showed a steady decline during the 8-day culture under serum-free conditions after the 24-hour EBs were transferred onto the gelatin-coated dishes. The first 4-day activin A treatment (5 or 50 ng/mL) prevented the time-dependent decrease in their mRNA expression, although the downregulation of NANOG mRNA was not significant. On the other hand, SB431542 treatment (5 μM) produced no significant differences with the no treatment group (Fig. 3).
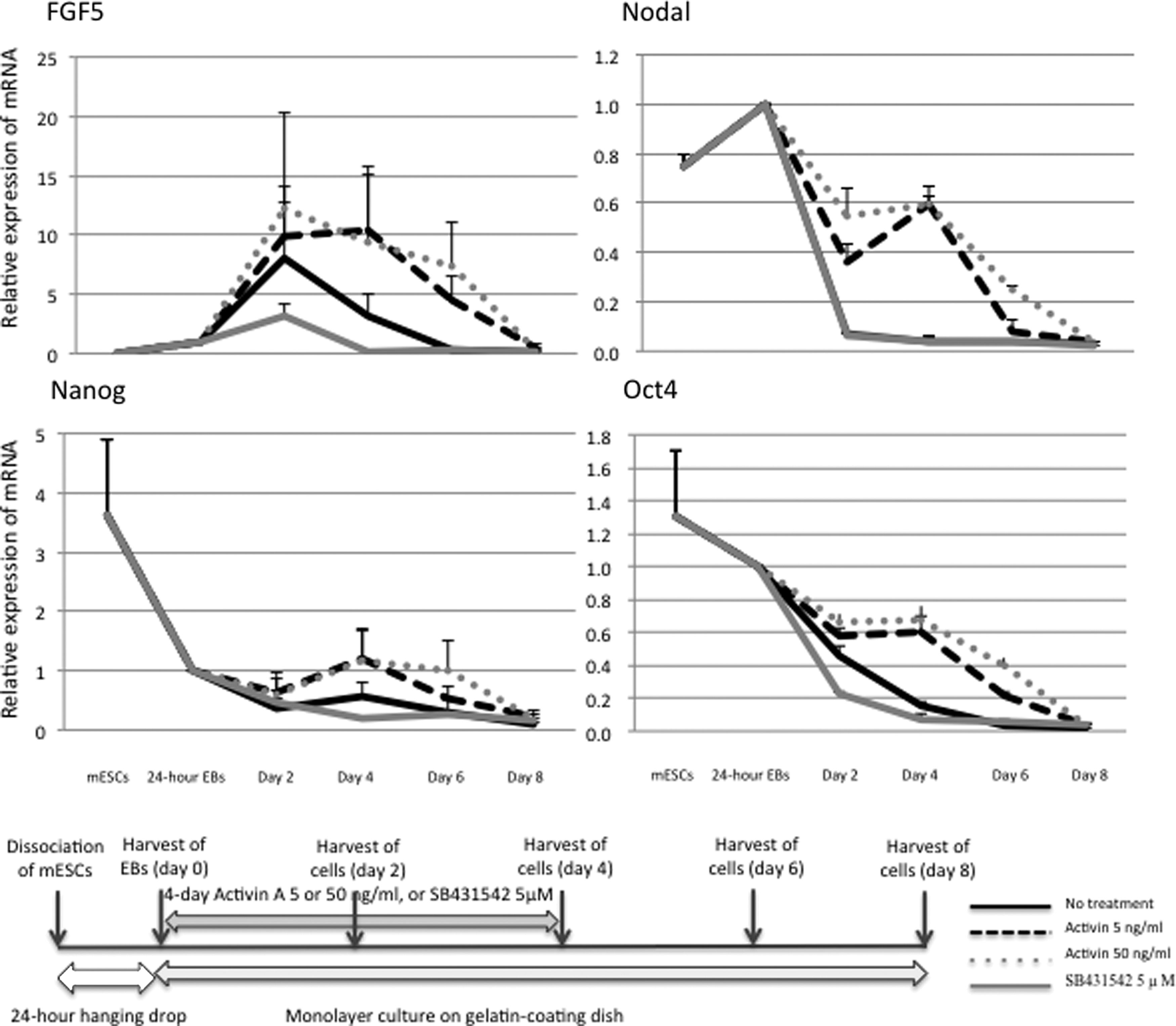
Real-time quantitative RT-PCR analyses for FGF5, nodal, NANOG, and Oct4 mRNA in the EBs that were allowed to differentiate for a short period in the presence of activin A or a nodal inhibitor, and schematic representation illustrating the experimental protocol. Real-time quantitative RT-PCR analysis was performed on each sample prepared from mESCs, the 24-hour EBs, and the EBs that were allowed to further differentiate for 2, 4, 6, or 8 days in the presence of 5 or 50 ng/mL activin A or 5 μM SB431542 for the first 4 days. The data represent mean ± SE of three independent experiments. Activin treatment at 5 and 50 ng/mL made a significant difference (p < 0.05) in FGF5, Nodal, and Oct4 mRNA expression in comparison with no treatment control or SB431542 treatment.
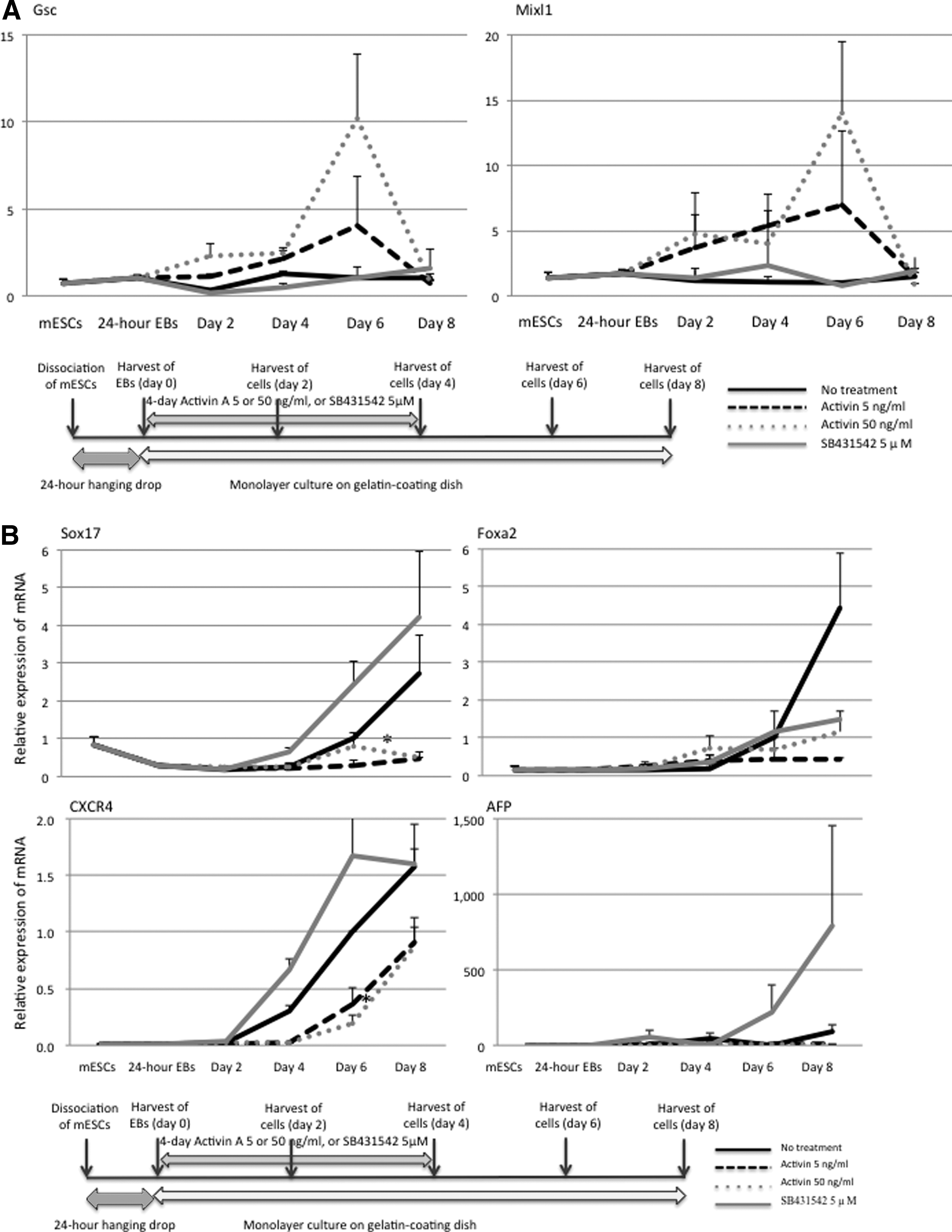
The temporary induction of mesendodermal transcription factors—goosecoid (Gsc) and Mix1 homeobox-like 1 (Mixl1)—was caused depending on the dose of the first 4-day activin A treatment. The mRNA expression of Foxa2, Sox17, AFP, and CXCR4 (specific to the endoderm) was also examined. Their expression showed a spontaneous increase by the eighth day after the transfer of the 24-hour EBs. Activin A treatment (5 or 50 ng/mL) significantly prevented their expression in comparison with the no treatment group. SB431542 treatment (5 μM) significantly upregulated the expression of Sox17, CXCR4, and AFP mRNA in comparison with activin A treatment (5 or 50 ng/mL). The immunofluorescence staining to detect Fxa2, AFP, and CXCR4 also showed the increased protein expression by SB431542 treatment (Fig. 4). The experiments that we performed on miPSCs showed the same results as did the experiments on mESCs (data not shown).
Induction of differentiation of EBs into the endodermal lineage by RA
We used RA to promote differentiation of the transferred EBs into the endodermal lineage. One report showed that RA induces formation of Pdx1-positive cells from EBs that developed from mESCs (Micallef et al., 2005). The time course of 48-hour RA treatment (10−6 M) was examined first. The 24-hour EBs that we produced from mESCs were transferred onto the gelatin-coated dishes (day 0). Then, they were allowed to attach under serum-free conditions for 12 or 18 days. The 48-hour RA treatment was carried out on day 0, 2, 4, 6, or 8; after that Pdx1 mRNA expression was analyzed on days 12 and 18. When the RA treatment was performed on day 2, the highest mRNA expression of Pdx1 was induced both on days 12 and 18 (Fig. 5A). The mRNA expression of the other endodermal lineage markers, including albumin and glucagon, was also induced most strongly when the treatment started on day 2.
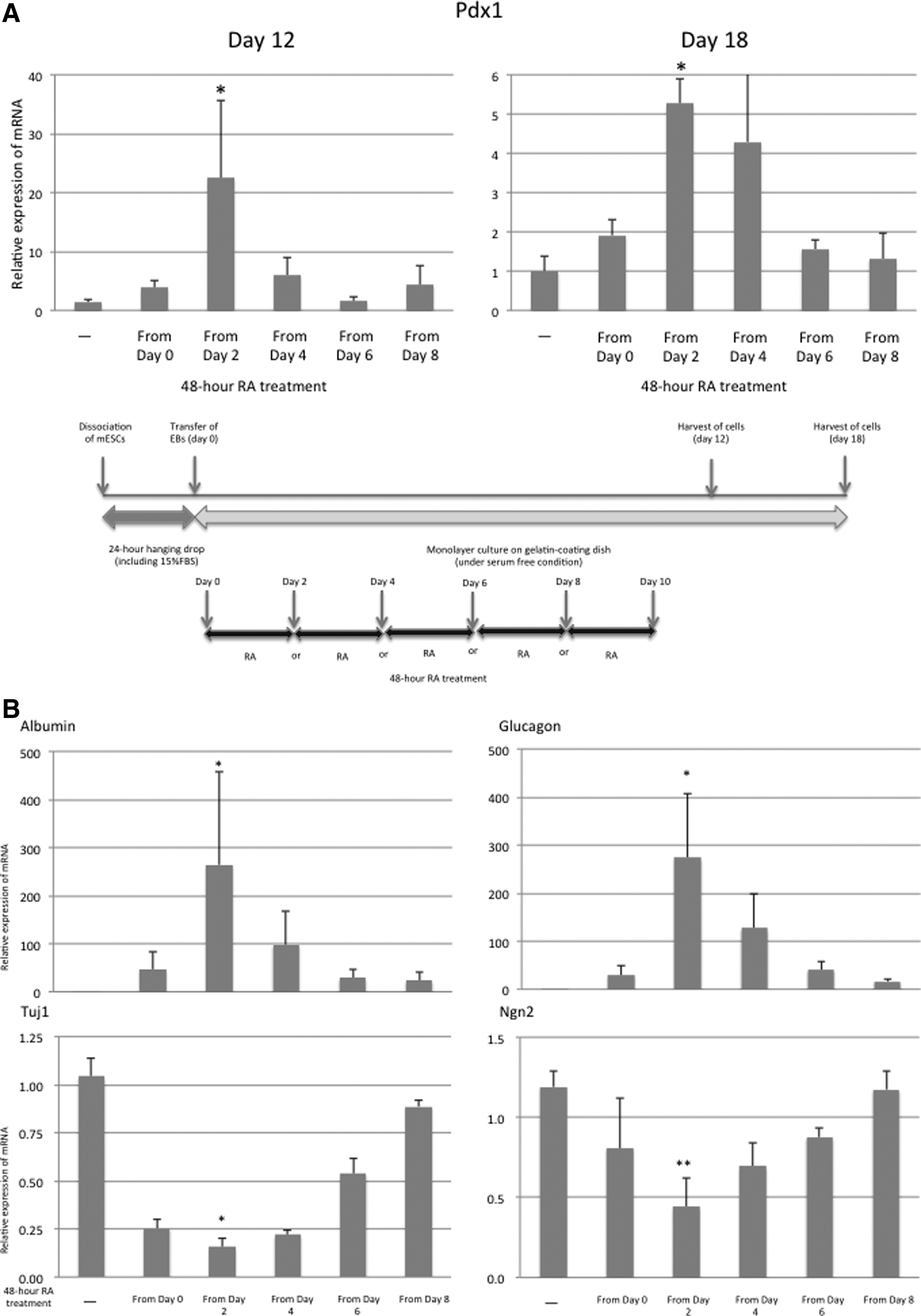
The RA treatment on day 2 induced the weakest differentiation into the neural lineage (Fig. 5B). Consequently, the 48-hour RA treatment starting on day 2 was used in our subsequent experiments.
The impact of activin A treatment on differentiation of EBs into the endodermal lineage under the influence of RA
The transferred 24-hour EBs that we produced from mESCs were allowed to differentiate, with 4-day activin A treatment starting on day 0 and/or 48-hour RA treatment starting on day 2 under serum-free conditions for 12 or 18 days. On day 12, mRNA expression of the endoderm lineage markers, including Foxa2, CXCR4, and AFP, was examined. Activin A treatment (50 ng/mL) alone had no effect on induction of their expression. The RA treatment successfully induced the differentiation into the endoderm. Activin A treatment (50 ng/mL) significantly attenuated the induction of Foxa2 and CXCR4 by the RA treatment. Just as the 12-day differentiation procedure on the 24-hour EBs, the first 4-day activin A treatment (50 ng/mL) failed to induce differentiation into the endoderm, for example, it failed to induce expression of Pdx1, glucagon, and insulin 1, even though the EBs were allowed to differentiate for 6 more days. The RA treatment significantly enhanced their mRNA expression in the EBs after the 18-day differentiation procedure. When activin A treatment (50 ng/mL) was combined with RA treatment, the RA-induced endoderm-related mRNA expression was offset (Fig. 6A).
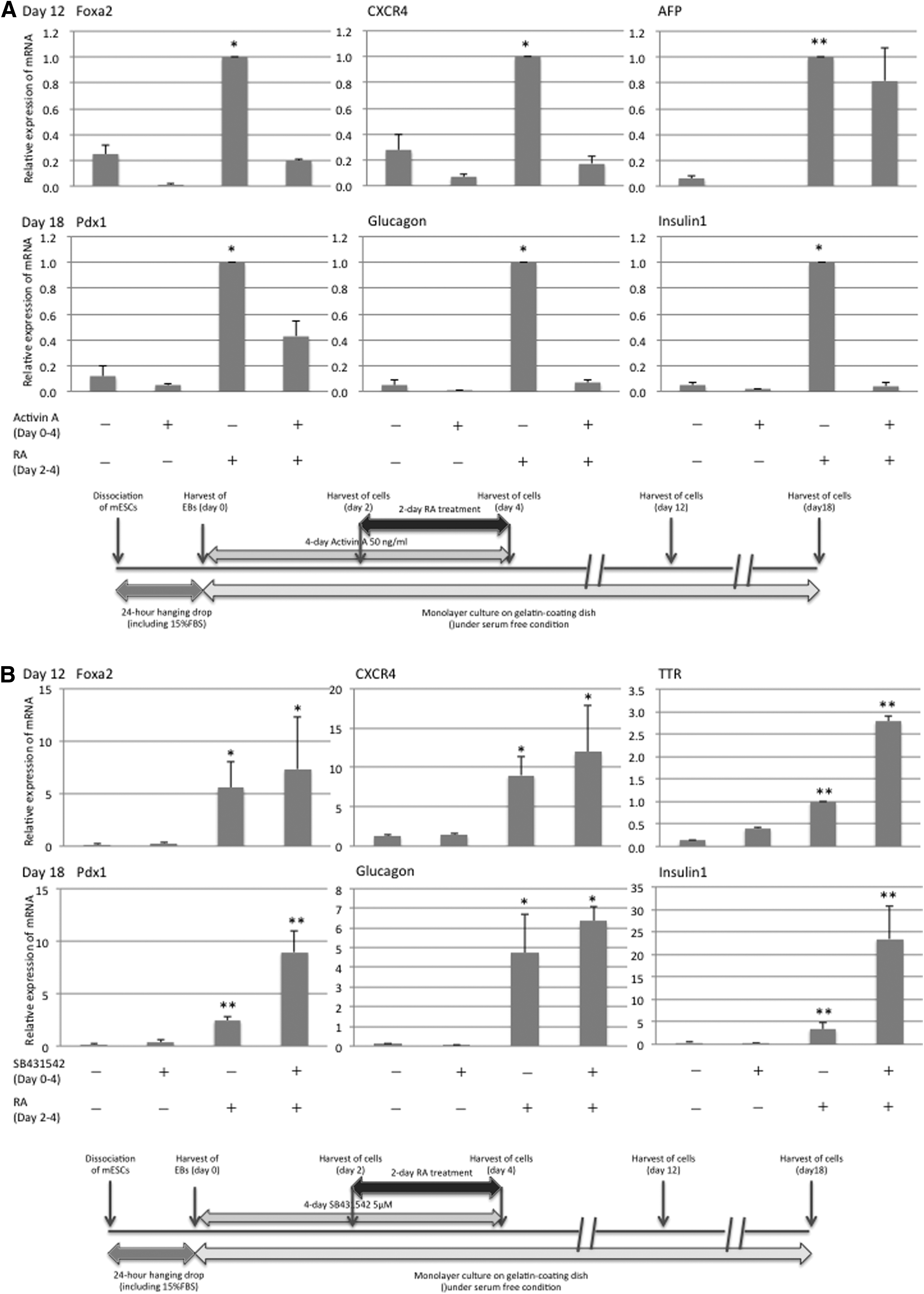
Next, the transferred 24-hour EBs were subjected to SB431542 treatment (5 μM) to inhibit activin/nodal signaling for the first 4 days, with the 48-hour RA treatment starting on day 2 under serum-free conditions for 12 or 18 days. The SB431542 treatment alone had no effect on the expression of the endoderm-related transcripts, including Foxa2, CXCR4, and TTR mRNA, on day 12. The SB431542 treatment did not inhibit the induction of the endoderm lineage transcripts by RA. On the contrary, it promoted the RA-induced TTR mRNA expression more strongly than did the RA treatment alone. Moreover, on day 18, inhibition of activin/nodal signaling with SB431542 treatment for the first 4 days resulted in higher expression of the endodermal lineage transcripts, including Pdx1, glucagon, and insulin 1 mRNA, in the EBs treated with RA under serum-free conditions, although the increase in glucagon mRNA expression was not statistically significant (Fig. 6B). The experiments that we performed on miPSCs yielded the same results as the experiments on mESCs (data not shown).
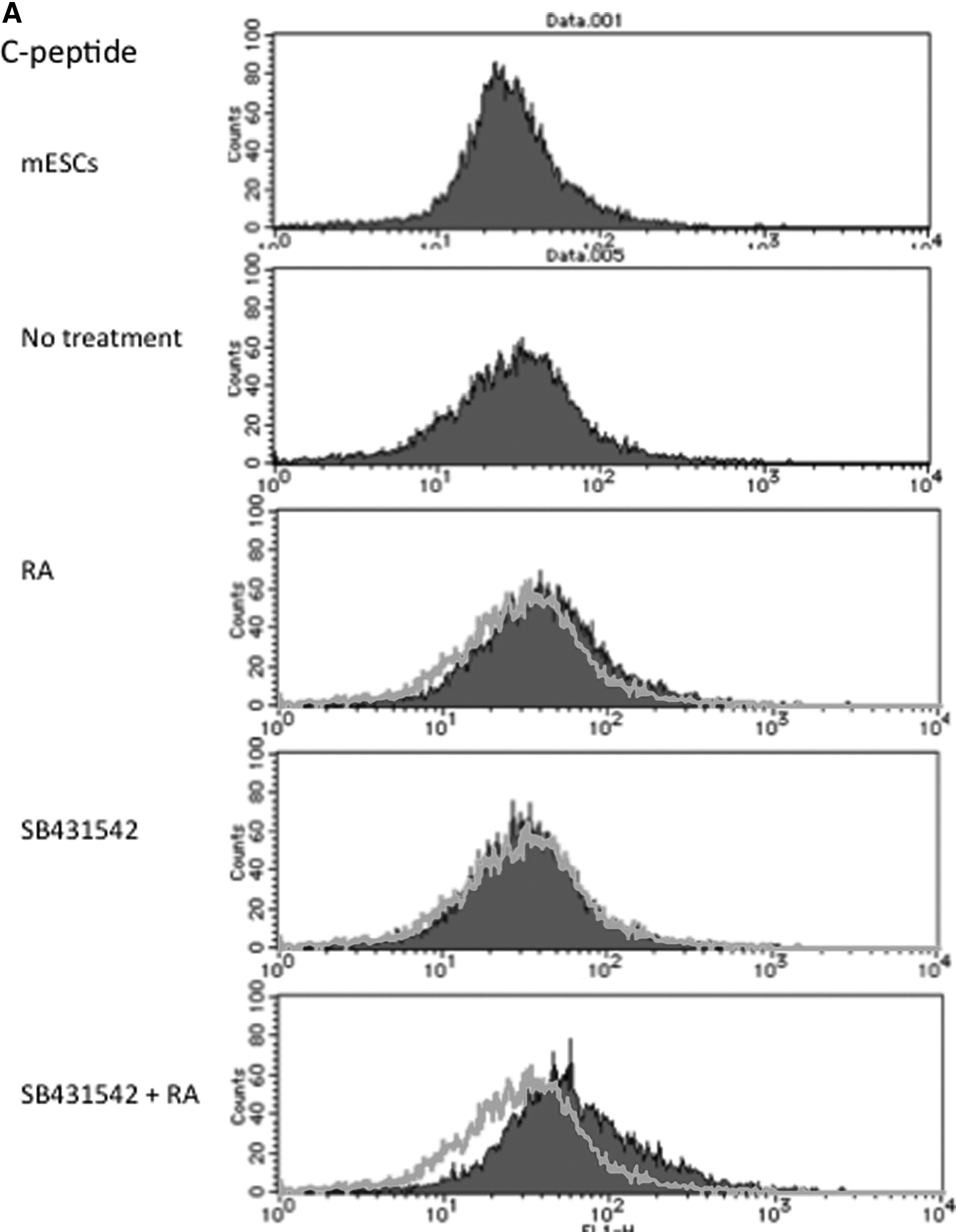
Fluorescence-activated cell sorting (FACS) analysis showed that C-peptide expression was induced after the 18-day differentiation procedure with the 48-hour RA treatment, and the nodal inhibitor enhanced the RA-mediated induction of C-peptide. Immunofluorescence staining to detect Foxa2, AFP, TTR, Albumin, Pdx1, and Proinsulin was performed. The samples were harvested after 12 days for Foxa2, AFP, and TTR and after 18 days to detect Albumin, Pdx1, and Proinsulin. Foxa2, AFP, and TTR protein expression was detected following RA treatment and it was increased when SB431542 was added. SB431542 treatment induced modest expression of these proteins, confirming the results of RT-PCR. After 18 days, RA treatment induced Albumin, Pdx1, and Proinsulin expression and the addition of SB431542 strengthened it (Fig. 7).
Discussion
The cells in the ICM meet either the primitive endoderm or ectoderm cell fate. The early events of postimplantation development, between E4.5 and E6.5, involve ingression and subsequent differentiation of the primitive ectoderm through the primitive streak; these events lead to formation of the three germ layers (Rathjen et al., 1999).
The appearance of developmental markers and their overall distribution in EBs strictly correlate with their expression patterns observed at specific stages of embryogenesis; all embryonic cell lineages are also sufficiently represented in EBs. Leahy et al. (1999) reported that expression of FGF5, which is a prominent marker of the early postimplantation embryo within the primitive ectoderm and is typical of mEpiSCs, is detectable in the EBs as early as the first day in suspension culture. Moreover, the EBs that were produced by the HD method show more rapid and complete development of mESCs than do the EBs that were produced in static suspension culture (Mogi et al., 2009; O'Shea, 1999). As expected, we confirmed the expression of FGF5 in the 24-hour EBs. When the EBs were produced from mESCs by the 24-hour HD method, some of the EBs acquired characteristics of mEpiSCs or the epiblast. Moreover, the transition from mESCs to epiblast-like cells when we used the 24-hour HD method was supported by the decrease in mRNA expression of Rex1 and Klf4 (characteristic of mESCs) and by the increase in brachyury mRNA expression in mEpiSCs. mESCs are compared with the state of the ICM. The ICM can develop into epiblast.
We did not know whether LIF could prevent spontaneous differentiation from mESCs to epiblast-like cells during 24-hour HD. Therefore, we employed not EBs made by 24-hour HD with LIF, but mESCs maintained under a combination of LIF and serum, corresponding to the ICM, as the experimental control to examine epiblast-like cells in the 24-hour EBs. Guo et al. (2009) found that mESCs that are transferred to mEpiSC culture conditions continue to proliferate and become relatively homogeneous and mEpiSC-like, showing the marker profile characteristic of mEpiSCs rather than of mESCs, with maintained expression of Oct4, somewhat downregulated NANOG, and strongly downregulated Rex1 and Klf4 (Hall et al., 2009). In the present study, mRNA expression of FGF5 and brachyury was induced in 24-hour EBs. Then, we showed that mESCs can grow in an epiblast-like manner spontaneously, whereas mESCs formed a cluster under culture conditions, including FBS.
In the early mouse embryo, mutations that perturb the level of nodal/activin signaling inhibit induction of the primitive streak/mesoderm, mammalian organizer, mesendoderm, and endoderm during establishment of the anterior–posterior axis (Perea-Gomez et al., 2002; Tremblay et al., 2000); in fact, it can induce differentiation of mESCs into the endoderm and mesoderm in culture (Schroeder et al., 2012; Tada et al., 2005). In contrast to the evidence that nodal/activin signaling predominantly promotes differentiation events, some studies showed that this pathway paradoxically plays important roles in the maintenance of self-renewal and pluripotency (Ogawa et al., 2007; Schroeder et al., 2012). Indeed, activin A is frequently used directly in culture for continued expansion of hESCs and mEpiSCs (James et al., 2005; Vallier et al., 2005).
Vallier et al. (2009) found that the nodal/activin signaling pathway directly controls the expression of NANOG in hESCs and in mEpiSCs, thereby blocking their differentiation toward the neuroectoderm lineage. Therefore, we were interested in what activin A does to the 24-hour EBs expressing transcripts characteristic of mEpiSCs or the epiblast. Around the transferred EBs, the monolayer cells that stained insignificantly with ALP expanded during serum-free culture. Activin A treatment caused the attached EBs to produce negligible outgrowth. In short, the results showed that the differentiation of attached EBs was inhibited by activin A treatment.
Brons et al. (2007) demonstrated that transcripts of FGF5 and nodal, the two genes specifically expressed in the late epiblast layer after implantation, are detectable in mEpiSCs. Nodal signals are also implicated in the maintenance of ES cell properties according to the finding that nodal-deficient mouse embryos show an epiblast that is substantially reduced in size, with negligible levels of Oct3 and Oct4 expression (Conlon et al., 1994; Robertson et al., 2003). Xiao et al. (2006) also found that activin can support long-term feeder-free culture and maintain pluripotency of hESCs possibly by inducing the expression of Oct4 and NANOG. In our experiments, activin A treatment showed a tendency to maintain the expression of FGF5 and nodal (characteristic of an epiblast-like state), as well as NANOG and Oct4 (indicative of pluripotency), in the transferred 24-hour EBs. In other words, extrinsic activin A seemed to inhibit, not induce, the differentiation in mESC clusters, including epiblast-like cells. Nodal expression is first detected during implantation (E4.5) in the ICM of the blastocyst (Mesnard et al., 2006) and thereafter it persists until the extended primitive streak stage (E7.5) and disappears before the maturation of the gut tube (Collignon et al., 1996; Meno et al., 2001). In our in vitro experimental system, nodal expression in the EBs made by 24-hour HD declined rapidly after transfer, and extrinsic activin treatment to the EBs maintained the nodal expression contrary to the in vivo endoderm differentiation situation. We could show no significant difference in the gene expression between 5 and 50 ng/mL of activin A, although the latter brought about the greater change in the gene expression than the former. Moreover, a lot of previous studies employed the concentration of 25–100 ng/mL to treat mESCs with activin A. Therefore, we used the concentration of 50 ng/mL in accordance with many previous reports, although the lower dose of activin A also produced significant differences in the gene expression in comparison with no treatment.
During gastrulation in mice, the definitive endoderm lineage originates from a common bipotential progenitor cell type called the mesendoderm. Formation of the mesendoderm population is induced by nodal signaling in the most anterior region of the primitive streak (epiblast layer) (Tam and Loebel, 2007). In fact, studies on mESCs also showed that formation of endoderm cells can be induced by treatment with activin A (Kubo et al., 2004; Tada et al., 2005). Therefore, we tested whether extrinsic activin A can induce differentiation from the epiblast-like state into the mesendoderm or endoderm lineage. It was confirmed that the spontaneous differentiation into the endodermal lineage had been caused in the EBs made by HD for 24 hours during 8-day culture. In particular, SB431542 treatment induced the remarkable increase of AFP mRNA expression compared with no treatment. Although it is certain that the inhibition of nodal signaling promoted differentiation in the EBs, we are not able to explain such a prominent increase of AFP.
Although mesendoderm markers, including Gsc and Mixl1, showed temporary upregulation after the activin A treatment, the subsequent endoderm-related pattern of mRNA expression was not observed. In one report, activin treatment was shown to transcriptionally activate the Gsc gene (Ku et al., 2005). Moreover, Mixl1 acts downstream of the nodal/activin signaling pathways and regulates formation of both the mesoderm and endoderm (Hart et al., 2002; Poulain and Lepage, 2002). Our experiments also revealed activin-induced differentiation into the mesendoderm. Nonetheless, the temporary activation of mesendodermal genes was not followed by expression of endodermal genes, such as Sox17 and Foxa2.
Some researchers have reported that extrinsic activin A has little or no effect on the differentiation of mESCs into the endoderm lineage, whereas there are a lot of studies where activin A treatment was used to induce formation of the definitive endoderm from mESCs (Kubo et al., 2004; Nakanishi et al., 2009; Parashurama et al., 2008; Tada et al., 2005). Teo et al. stated that the function of activin/nodal signaling is defined by the presence of specific partners of SMAD2 or SMAD3, which can either maintain pluripotency or induce differentiation (Teo et al., 2011). Moreover, Tsuneyoshi et al. (2012) found that SNON is a key component of the nuclear machinery that safeguards pluripotency through direct repression of differentiation-related genes downstream of activin/nodal. SNON is an SMAD repressor and it exerts this suppressive action by interacting with SMAD complexes, thus inhibiting their transcriptional activity (Luo, 2004).
In general, activin A induces differentiation of mESCs into the endoderm, but activin A inhibits differentiation of mEpiSCs. In our experiments, uncommitted mESCs must also have been present (in addition to the cells expressing the genes characteristic of mEpiSCs) in the 24-hour EBs. The temporary induction of mesendodermal transcripts and the maintenance of the pluripotency-specific pattern of gene expression may reflect the effects of activin A on the remaining uncommitted mESCs and epiblast-like cells produced in the EBs, respectively. As a result, the induction of mesendodermal genes may be too weak to be followed by the endoderm-specific pattern of mRNA expression.
RA is a ligand of nuclear RA receptors, which control differentiation of progenitor cells important for vertebrate development (Gudas, 2013). It is involved in regionalization of the embryonic endoderm. Specifically, it has been revealed that RA signaling at the end of gastrulation is required for both liver and pancreas specification (Stafford and Prince, 2002). RA is also used to induce differentiation into endodermal lineages and is known to induce formation of Pdx1-positive cells in EBs (Cai et al., 2010; Johannesson et al., 2009; Micallef et al., 2005; Shirasawa et al., 2011). We employed the RA-induced endoderm differentiation protocol reported by previous studies (Micallef et al., 2005; Nakanishi et al., 2007; Shi et al., 2005). They employed RA treatment to the EBs made by suspension culture for 2–4 days to induce endoderm.
We applied the RA-induced endoderm differentiation system to examine whether extrinsic activin treatment to mESC clusters, including epiblast-like cells, promotes or inhibits endoderm differentiation. Accordingly, we first identified the most effective time point for treatment of the transferred EBs with RA. RA treatment starting on day 2 was most effective not only at inducing differentiation into the endoderm but also at producing less abundant neural lineages. This approach seemed to induce differentiation into more specialized endodermal lineages. Maybe further protein analysis of the examined genes should be performed because there was only a modest difference in the gene expression by RA treatment after 2 and 4 days. However, in our experiment, the RA treatment from day 2 showed significant differences in the gene expression than no RA. The aim of the present study was to examine the effect of nodal signaling in the experimental system to promote endoderm differentiation. It did not seem that it was an important matter to our experiment to further examine the timing of the RA treatment to induce endoderm differentiation because the RA treatment from day 4 showed no significant differences in the gene expression than no RA.
Activin A was unable to induce differentiation into the endoderm; moreover, activin A reversed the differentiation into the endoderm under the influence of RA. This finding confirmed maintenance of pluripotency in epiblast-like cells treated with activin A. Moreover, inhibition of nodal/activin signaling in epiblast-like cells promoted the differentiation into the endoderm under the influence of RA. In other words, a temporary inhibition of nodal/activin signaling in epiblast-like cells may support the transition from the pluripotent state to specification of the endoderm. Parashurama et al. (2008) studied the effects of activin A on mESCs cultured on collagen gels. They cultured mESCs on fibronectin-coated collagen gels in the presence of 10% serum for 10 days in the presence of activin A. The latter caused a dose-dependent decrease in the expression of endoderm markers while inducing the expression of epiblastic and mesendodermal markers, such as FGF5 and brachyury. Moreover, follistatin, a known activin inhibitor, increased the endodermal population, as we saw in our study. Those authors stated that an increase in the activin A concentration may shift the kinetics of differentiation into the epiblast and mesendoderm from rapid downregulation to persistent expression (Parashurama et al., 2008).
Our experiments also suggest that the maintenance of expression of epiblast-like cell markers by activin is related to inhibition of differentiation into the endoderm, in agreement with the above study. There have been some reports of failure to induce effective differentiation into the endoderm by extrinsic activin treatment (Nakanishi et al., 2009; Sherwood et al., 2007). When activin fails to induce formation of the endoderm effectively, the properties of epiblast-like cells need to be considered; these cells can readily form as a result of spontaneous differentiation of mESCs.
In summary, we detected expression of an epiblast-like marker in the EBs that were produced from mESCs by the 24-hour HD method. In the transferred EBs, including epiblast-like cells, activin A treatment maintains the characteristics of the epiblast and of the pluripotent state while inducing weak differentiation into the mesendoderm. Activin A treatment inhibits, not induces, differentiation into the endoderm. Moreover, inhibition of nodal/activin signaling in the epiblast-like cells promotes differentiation into the endoderm, suggesting that temporary inhibition of nodal/activin signaling in epiblast-like cells may facilitate the transition from the pluripotent state to specification of the endoderm.
Footnotes
Acknowledgment
Y.S. was supported by a grant from the Takeda Science Foundation.
Author Disclosure Statement
The authors declare that no conflicting financial interests exist.