
Abstract
Acute kidney injury (AKI) is mainly characterized by rapid decline of renal function. Currently, the strategy of stem cells might be a therapy to treat AKI. The objective of this study was to obtain human urine-derived cells (HUCs) from patients with AKI, followed by establishing induced pluripotent stem (iPS) cell line. We isolated urine cells from patients with AKI and found that the cells could survive long term with epithelioid morphology and maintain a normal karyotype. The cell line had expression of renal-specific markers and renal development-related genes. After induction, the urine cells cotransfecting with TET-ON vectors were converted into iPS cells. The HUC-derived iPS (HUC-iPS) was positive for alkaline phosphatase staining, and had expression of pluripotency markers, consistent with human embryonic fibroblast-derived iPS cell. Notably, HUC-iPS could be induced to undergo directional kidney precursor cells (KPCs) differentiation under defined conditions, and transplantation of KPCs resulted in reducing kidney damage from ischemia–reperfusion injury in mice. Therefore, we successfully established HUC-iPS cell from patients with AKI and provided a novel stem cell resource for cell therapy in AKI.
Introduction
Acute kidney injury (AKI) is a common clinical syndrome. After acute renal failure (ARF) develops, it has a high fatality rate and is a common life-threatening disease in clinical practice. In the past few decades, despite the great development of renal replacement techniques, the fatality rate of ARF without complications has been ∼5%–10%, and for ARF with extrarenal organ failure, it has been 50%–70% (Himmelfarb et al., 2008). Thanks to the repair ability of the kidney, renal function may be restored after the termination of damage factors (Benigni et al., 2010; El Sabbahy and Vaidya, 2011). In clinical study, damage factors have been terminated, and further treatment of patients with renal injury is focused on how to promote kidney structure and function.
Recently, the application of stem cells has increasingly attracted attention in the medical field. In particular, the establishment of induced pluripotent stem cells (iPSCs) derived from adult cells makes it possible for individualized therapy, and provides a new idea for the rescue of AKI by using iPSCs (Takahashi and Yamanaka, 2006). For the generation of iPSCs requiring many cells for reprogramming, undergoing an invasive procedure for donating samples is one of the issues to be addressed. However, isolation of exfoliated cells present in urine could represent an excellent candidate for noninvasive cell reprogramming (Zhou et al., 2012).
Remarkably, it is estimated that each human kidney processes ∼100 L of filtrate per day (Witzgall, 2008). Yet, only about 2 L are normally excreted through the urethra as urine daily, and ∼2000–7000 cells could be collected in urine (Rahmoune et al., 2005). At the early stage of AKI, the kidney suffers from damage of tubular epithelial cells (TECs), and the patient has oliguria or anuria. During the repair period, epithelioid cells increase in the urinary sediment, accompanied by the recovery of urine volume (Kanbay et al., 2010; Kellum et al., 2015).
Sutherland was the first to report the successful culture of exfoliated urinary cells from the urine of newborns (Sutherland and Bain, 1972), and multiple groups have reproduced this since (et al., 2000Dörrenhaus et al., 2000; Herz, 1980; Linder, 1976). Notably, Zhang and colleagues have also identified a subpopulation of urinary cells that express stem cell markers and have multidifferentiation potential (Bharadwaj et al., 2011; Zhang et al., 2008). Recent studies have revealed that renal progenitor cells with anti-inflammatory and antiapoptotic phenotypes are present in fetal amniotic fluid (mainly fetal urine) (Monteiro Carvalho Mori da Cunha et al., 2015).
Arcolino's team isolated renal progenitor cells from the urine of preterm infants for the first time and could differentiate them into functional podocytes and proximal tubule cells in vitro (Arcolino et al., 2016). These studies suggested that urine-derived renal progenitor cells could exist in fetuses' or newborns' urine. However, there are still a lot of debates about the existence of renal progenitor cells in adult kidneys, so the use of renal progenitor cells as rescue cells for AKI remains to be further studied.
Since the milestone achievement by Takahashi and Yamanaka (2006), iPSCs derived from a variety of human tissue cells have been successfully established, including urine-derived kidney cells (Zhou et al., 2012). Currently, urine cells are mainly derived from neonates, preterm infants, and fetuses whose kidneys have not yet fully developed, whereas the efficiency of obtaining renal cells from the urine of children and adults was rare (Arcolino et al., 2016; Monteiro Carvalho Mori da Cunha et al., 2015; Sutherland and Bain, 1972).
It has been reported that the isolation efficiency of urine cells from patients with hereditary kidney disease is higher, but still low (Lazzeri et al., 2015). In the process of kidney repair, the urine volume of patients gradually recovered, and various cell components in the urinary sediment also increased continuously, implying a potential best time to isolate renal cells from the urine (Kanbay et al., 2010). However, whether the urine-derived cells from patients with AKI could be isolated and converted into iPSCs has not been reported.
In this study, we successfully obtained kidney cells from patients with AKI by the special urine cell separation method. Through immunofluorescence analysis, we found that human urine-derived cells (HUCs) had expression of renal proximal tubular epithelial markers, such as Pax8, E-cadherin, and AQP1, as well as renal development-related genes (Pax2, WT1, Six1, and Six2). The cells were cotransfected with TET-ON-induced expression vector plasmids by nuclear transfection technology, and monoclonal transgenic cell lines were obtained under the condition of drug screening.
Subsequently, we established HUC-derived iPS (HUC-iPS) and differentiated HUC-iPS or human embryonic fibroblast-derived iPS (HEF-iPS) into kidney precursor cells (KPCs), respectively. Moreover, kidney injury after ischemia–reperfusion injury (IRI) was relieved by transplantation of KPCs. Our study laid a foundation for the establishment of urine-derived iPSCs from patients with AKI and provided a novel stem cell resource for cell therapy in AKI.
Materials and Methods
Chemicals and urine samples
All chemicals were purchased from Millipore Sigma (Burlington, MA) and Thermo Fisher Scientific unless otherwise indicated. Urine samples from patients with AKI were provided by the Department of Nephrology, Nanjing First Hospital, Nanjing Medical University. Our study protocols were approved by the Medical Ethics Committee and Animal Care and Use Committee at Nanjing Medical University.
Urine cell isolation and culture
The midstream urine of patients in the recovery stage of AKI was collected by using special devices into sterile centrifuge tubes. About 30–50 mL of the urine was stored at 4°C and transported to the laboratory within 2 hours. The urine cells were centrifuged at 4°C, 800 g, 15 minutes, and then cultured in a six-well plate (82% high-glucose Dulbecco's modified Eagle's medium [DMEM], 15% fetal bovine serum [FBS], and 3% Pen Strep) for 24 hours. The cell culture medium was replaced with urine cell culture medium (knockout-DMEM, 1% ITS, 200 ng/mL FGF9, 50 ng/mL BMP7, 0.1 μmol/L RA, 1 μg/mL heparin, 10 ng/m EGF, 10 ng/m activin) A, 300 ng/mL BMP4, 15% FBS, and 1% Pen Strep). After 7–10 days of culture and changing the culture medium every 2 days, urine cells were obtained.
Human proximal TECs (HK2) and human embryo kidney cells (HEK293) were used in this study. The HK2 cells and HEK293 cells were immortal cells (a gift from Dr. Xiubin Liang, Nanjing Medical University, China) that were cultured in DMEM/F-12 (Gibco, Grand Island, NY) supplemented by a 10% FBS (Gibco BRL) and a 1% penicillin–streptomycin solution (Gibco) at 37°C in 5% CO2.
Karyotyping
The chromosomes of established urine cells were analyzed at passages 1 and 2, respectively. Urine cells were incubated in a medium supplemented with 0.02 mg/mL colcemide for 1 hour at 38.5°C in an atmosphere of 5% CO2 in air. After trypsinization and treatment with hypotonic KCl (0.56%) for 30 minutes, the cells were fixed with a 3:1 (v/v) mixture of methanol and acetic acid and spread on clean microscopic slides by gentle dropping. After staining with Giemsa (1:10 dilution) for 20 minutes, the chromosomes were examined under an oil microscope.
Immunofluorescence
For immunofluorescence staining, the HUCs, HK2 cells, HEK293 cells, HUC-iPS, HEF-iPS, and KPCs (12 days differentiation) were cultured on coverslips and fixed with 4% paraformaldehyde for 10 minutes, permeabilized with 1% Triton X-100 for 1 hour, and blocked with 10% goat serum for 1 hour at room temperature. The cells were then incubated with primary antibodies at 4°C overnight and incubated with secondary antibodies for 1 hour at room temperature. The primary antibodies included Pax8 (1:100; Abcam, Cambridge, MA), E-cadherin (1:200; Proteintech, Wuhan, China), AQP1 (1:200; Proteintech), Pax2 (1:200; Abcam), WT1 (1:200; Abcam), Six1 (1:100; Santa Cruz Biotechnology, Dallas, TX), Six2 (1:100; Proteintech), OCT4 (1:100; Santa Cruz Biotechnology), SOX2 (1:200; Calbiochem), and TRA-1-60 (1:200; Millipore).
Secondary antibodies included goat antirabbit IgG Alexa Fluor 546, goat antirabbit IgG Alexa Fluor 488, and goat antimouse IgG Alexa Fluor 546 (Thermo Fisher Scientific). The nuclei were stained with DAPI, and the stained cells were examined using a fluorescence microscope. The image acquisition, analysis, and processing were standardized within each experiment.
Cell transfection and selection
Pairs of 4 μg of the TetO-FUW-OSKM vector and 4 μg of the FUW-M2rtTA vector with 0.5 μg of the neomycin-expression plasmid (pCMV-tdTomato) were cotransfected into 1 × 106 urine cells using a basic fibroblast nucleofection kit (VPI-1002; Amaxa Biosystems-Lonza, Cologne, Germany) and nucleofection program U023, following the manufacturer's protocols. After 24 hours of recovery, the electroporated cells were selected with 100 μg/mL of G418 (Gibco) in 10-cm dishes for ∼10 days. Individual cell colonies were picked up and cultured in 24-well plates and then passaged to 12-well plates.
The single colony was lysed in NP-40 buffer at 55°C for 30 minutes and then at 95°C for 10 minutes. The lysate was used as a template for PCR screening. The primers used in amplifying the target region were as follows. For TetO-FUW-OSKM, forward: 5′-GAGGGAGAC CGAGGAGTTCAA-3′ and reverse: 5′-GCAGCGTATCCACATAGCGTAA-3′. For FUW-M2rtTA, forward: 5′-GGGGAGGGTAAGTGAGGC-3′ and reverse: 5′-GCTGACAGGTGGTGGCAAT-3′. The PCR conditions were 95°C for 5 minutes, followed by 35 cycles of 95°C for 30 seconds, 58°C for 30 seconds and 72°C for 90 seconds, and a final 72°C for 5 minutes. Transgenic HEF cotransfecting with TET-ON vectors had been established following the same steps described above in our laboratory (data not shown).
Generation of urine-derived cell iPS (HUC-iPS) and alkaline phosphatase staining
The transgenic cells (HUCs and HEF) cotransfecting with TET-ON vectors were seeded onto a monolayer of mitomycin C-inactivated mouse embryonic fibroblasts (MEFs) at a density of 1 × 105 cell/well. The MEFs were inactivated with 10 mg/mL mitomycin C for 2.5 hours, resuspended, and seeded in four-well dishes precoated with 0.1% gelatin. The iPS medium was 40% KnockOut™DMEM, 10% KnockOut™SR, 24% DMEM/F12, 24% neurobasal medium, 0.5% N-2 supplement, 1% B-27 supplement, 0.125 μg/mL BSA, 2.5 μg/mL insulin, 1 μmol/L
The transgenic cells were cultured at 38.5°C in a 5% CO2 atmosphere. The culture medium was changed every day. After 3 days, the culture medium was changed and the cell clones were observed. When HUC-iPS was grown, it was passed by mechanical method first, and then by the trypsin method. Alkaline phosphatase (AP) activity of the HUC-iPS at passage 4 was assayed with an AP detection kit (Millipore Sigma) according to the manufacturer's instructions. Reprogramming efficiency refers to AP-positive primary clones with human embryonic stem cell (ESC) morphology before cell passage.
Kidney precursor differentiation of iPS
The iPS (HEF-iPS and HUC-iPS) cells were tested for their potential to differentiate into defined lineages under chemically defined medium conditions in the absence of DOX support. They were differentiated into KPCs after 12 days of culture, as previously described by Takasato et al., in differentiation medium on a Matrigel-coated dish (Takasato et al., 2014).
PCR analysis
Total RNA was extracted from cultured cells at 0, 3, 7, and 12 days of HEF-iPS and HUC-iPS differentiation using Trizol, and cDNA was synthesized using a QuantiTect Reverse Transcription Kit (Vazyme, Nanjing, China). The sequences of the primers and annealing temperatures used are listed in Table 1. Real-time polymerase chain reaction (RT-PCR) analysis was also performed.
The Sequences of the Primers and Annealing Temperatures
Animal models and cell transplantation
A unilateral ischemia–reperfusion-induced AKI model and cell transplantation were established as previously reported (Li et al., 2016; Wan et al., 2015; Zhang et al., 2018). Male C57BL/6 mice aged 8 to 10 weeks were anesthetized using an intraperitoneal injection of chloral hydrate (10%, 0.35 mL/10 g). After a flank incision, the left renal pedicle was bluntly dissected and clamped by a microvascular clamp for 45 minutes. After the clamps were removed, 5 × 105 HUCs, FKPCs (KPCs, 12 days differentiation of HEF-iPS), or UKPCs (KPCs, 12 days differentiation of HUC-iPS) were injected under the capsule of the left kidney with a 32-gauge syringe needle.
Control mice underwent identical procedures, except the clamping of the renal pedicles and cell transplantation were omitted. The cohorts of mice were sacrificed at 7 days after surgery. Ligation of the right kidney was performed 1 day before sacrificing. The postischemic kidneys and control kidneys were harvested and fixed in 10% formalin until they were needed for further analysis. Blood samples were obtained at the time of euthanasia. Serum creatinine and blood urea nitrogen (BUN) were measured using an automated method.
Histology examination
Kidney sections from paraffin-embedded tissues were prepared at 4 μm thickness and stained with hematoxylin and eosin using standard procedures for histological evaluation. The sections were blind reviewed by a renal pathologist and scored using a previously described semiquantitative scale designed to evaluate the degree of tubulointerstitial injury according to tubular necrosis, tubular dilatation or atrophy, inflammatory cell infiltration, and cellar edema (Cao et al., 2004).
The kidney injury score was estimated at 200 × magnification using six randomly selected fields for each kidney. Injury was graded on a scale from 0 to 4. Higher scores represented more severe damage (maximum score, 4): 0, normal kidney; 1, minimal damage (<5% involvement of the cortex); 2, mild damage (5% to 25% involvement of the cortex); 3, moderate damage (25% to 75% involvement of the cortex); and 4, severe damage (>75% involvement of the cortex).
Statistical analysis
The data are expressed as mean ± standard deviations. A statistical analysis was performed using the Student t-test when only two value sets were compared. A one-way analysis of variance followed by Dunnett's test or Bonferroni correction was conducted when the data involved three or more groups. The differences were evaluated using SPSS10.0 software (SPSS, Chicago, IL). p < 0.05 was considered statistically significant.
Results
Isolation and identification of urine-derived renal cells from patients with AKI
Isolation of HUCs from urine involves minimal processing (Fig. 1A). Urine cells grew and formed primary cell clones (P0) with epithelial-like cell morphology for 2 days culturing, and formed larger clones after 6–7 days culturing, as shown in Figure 1B. Similar to the human proximal TEC line (HK2), the HUC has strong proliferative ability and can be passaged stably in vitro (Fig. 1C).
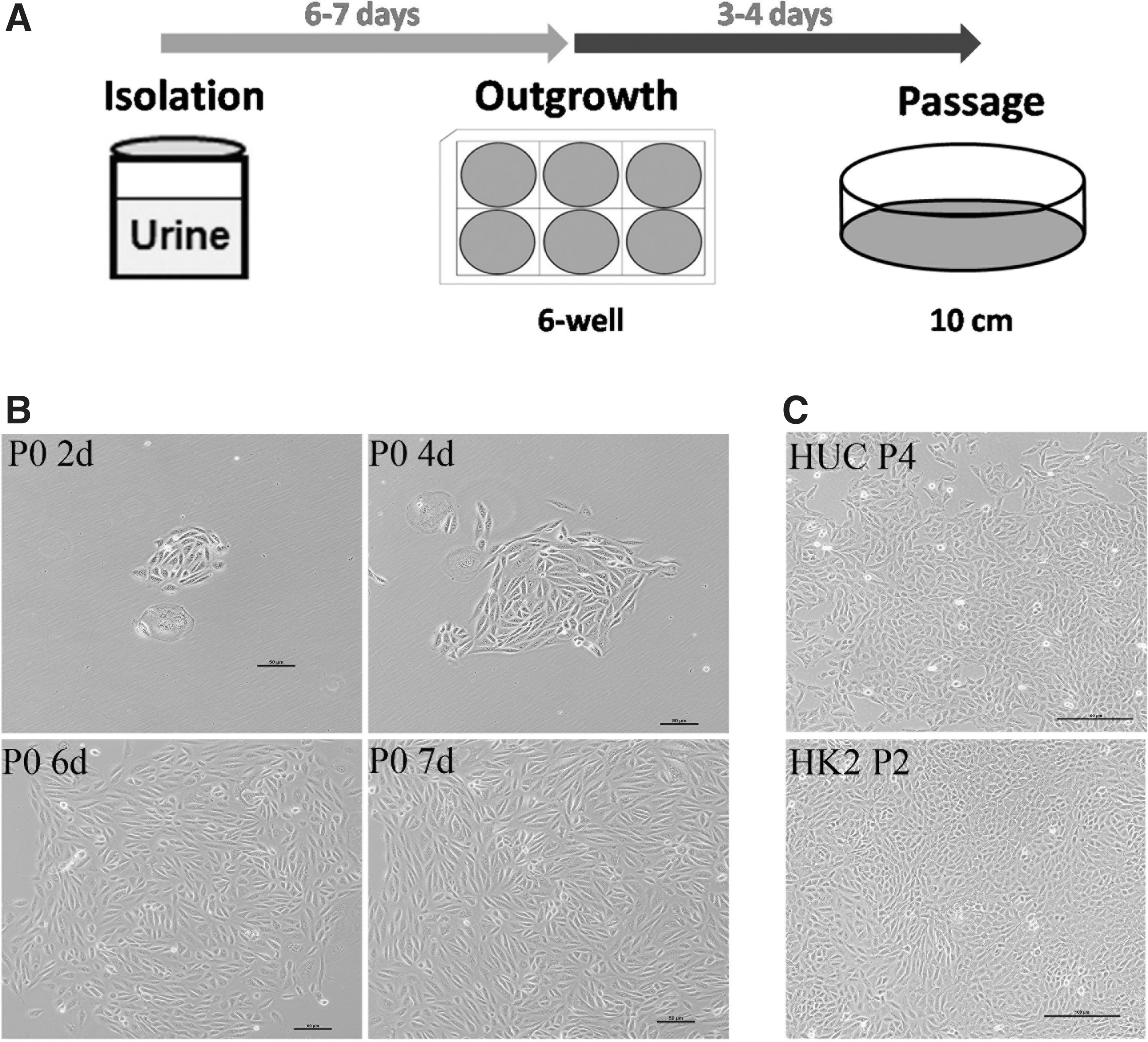
Schematic isolation of human urine-derived cell (HUC) from patients with AKI and morphology of HUC clone.
To determine whether urine cells displayed a normal karyotype, we analyzed the chromosomes of the different cell lines at different passages. Our results demonstrated that the urine-derived cells displayed a continued normal 46XY karyotype (Fig. 2A, B). In addition, renal proximal TECs-specific markers (Pax8, E-cadherin, and AQP1) were positively expressed in the cultured cell lines consistent with the HK2 cell line by immunofluorescence staining (Fig. 3A, B). The renal development-related genes (Pax2, WT1, Six1, and Six2) were also found to be expressed in the urine kidney cells and human embryo kidney cells (HEK293, positive control cell lines) (Fig. 4).

Karyotype analysis of urine-derived renal cell from different patients.

Determination of renal TEC-specific marker expression in human urine-derived cell (HUC) and human proximal TEC line (HK2).
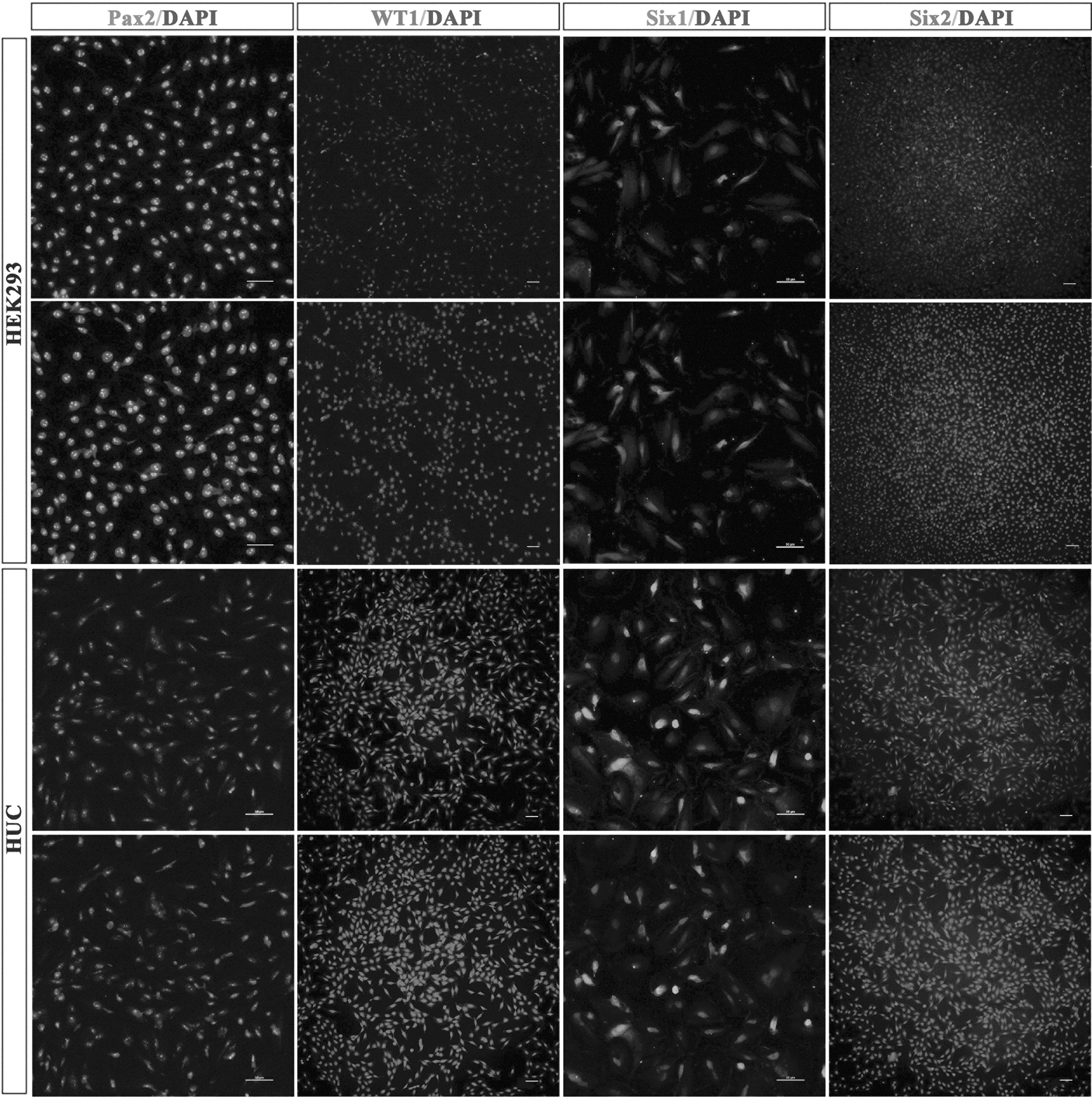
Nephrogenic molecules were detected in human urine-derived cell (HUC) and human embryo kidney cell (HEK293). Immunofluorescence analysis of Pax2, WT1, Six1, and Six2 in HUC and HEK293. Bar = 50 μm.
These results suggest that the isolated urine cells were derived from TECs and have some characteristics of renal progenitor cells. To test the nephrogenic potency, we also attempted to differentiate HUCs into TECs, as previously reported (Arcolino et al., 2016). For 1 day induction, cells became elongated and spindle shaped, accompanied by many cell deaths. After 4 days of culture, there were few differentiated cells alive, as shown in Supplementary Figure S1. Moreover, the differentiated cells had very limited proliferative capacity (data not shown).
Establishment of transgenic cell line with TET-ON vectors
To establish a transgenic cell line with TET-ON vectors (Fig. 5A), we transfected linear TetO-FUW-OSKM plasmids and FUW-M2rtTA plasmids (Fig. 5B) into urine-derived renal cells using a Lonza nuclear transfection kit. After 8 days of drug screening, the clones grew out, and the clones were a regular round shape with tightly arranged cells expressing red fluorescent protein, as shown in Figure 5C. The drug-resistant single cell colony was passaged, and the cell lysate derived from the same colony was used as a template for PCR screening. A total of 59 clones were selected in our study; only 5 clones were cotransfected with TetO-FUW-OSKM and FUW-M2rtTA vectors, as shown in Figure 5D.
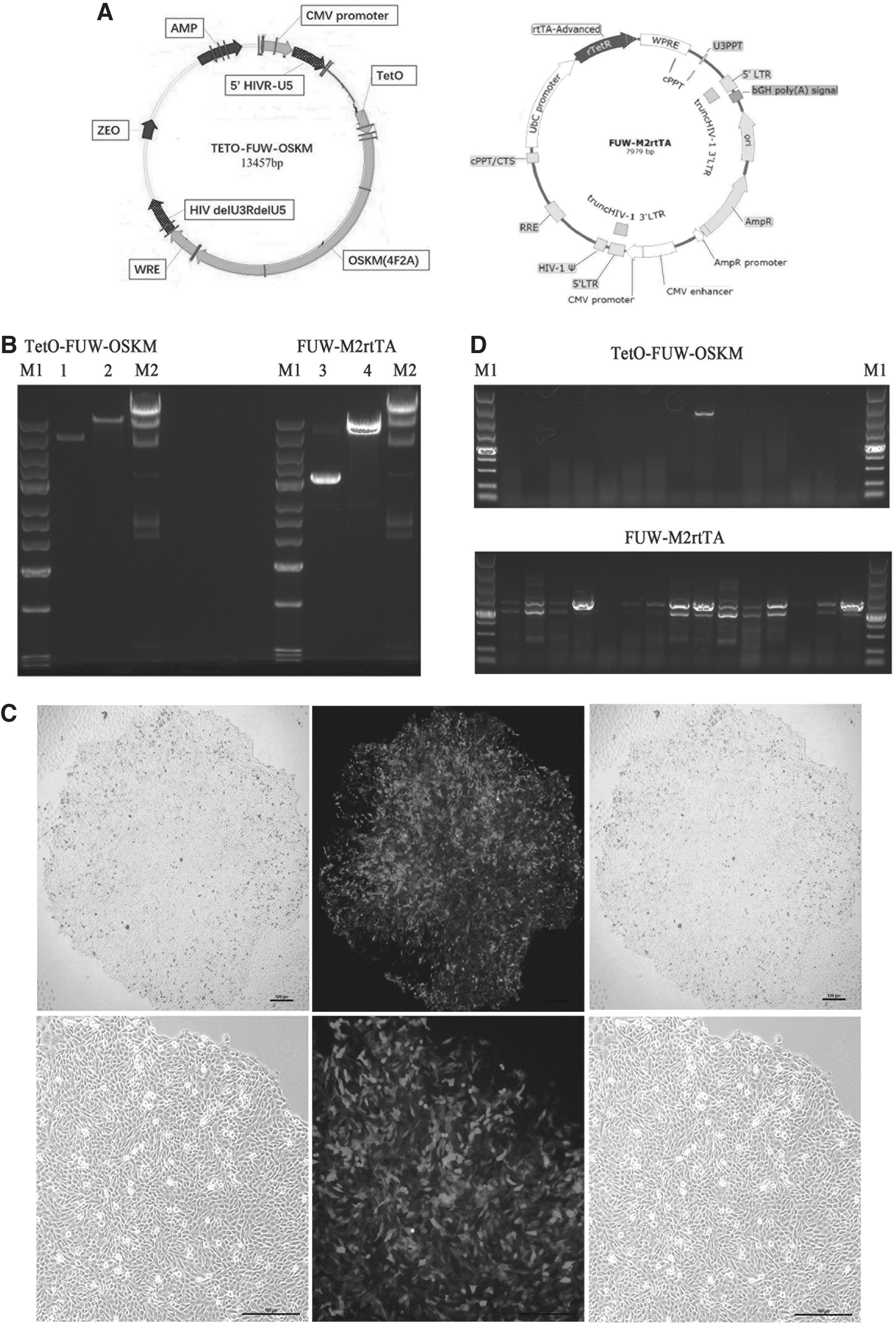
Transgenic cell lines with TET-ON vectors were established.
Generation and identification of urine-derived iPS (HUC-iPS) cells
After being induced by stem cell medium, the HUCs and HEF cotransfecting with TET-ON vectors showed domed morphology and suffered from single-cell passaging (Fig. 6A). The reprogramming efficiency of transgenic HUCs was higher than that of transgenic HEF (0.095 ± 0.007 vs. 0.075 ± 0.006, n = 4) (Fig. 6B). These established iPS lines exhibited many characteristics near naive pluripotency, including AP staining (Fig. 6A) and immunofluorescence for human ESC pluripotency markers (OCT4, SOX2, and TRA-1-60) (Fig. 6C).

Generation and identification of HUC-iPS and HEF-iPS.
Differentiation of iPS under chemically defined conditions
The iPSs (HEF-iPS and HUC-iPS) could also be induced to undergo successful differentiation under defined conditions. They differentiated into kidney precursors when cultured in defined medium without DOX for 12 days (Fig. 7A). The expression of the primitive streak stage-related gene MIXL1, intermediate mesoderm stage-related genes LHX1, and metanephric mesenchyme stage-related genes HOXD11 were detected by RT-PCR when differentiated at different time points (days 0, 3, 7, and 12) in HUC-iPS. We found that the expression of these genes was upregulated in the kidney precursors compared with HUC-iPS at day 0 (Fig. 7B).

HUC-iPS and HEF-iPS were directed to differentiate into kidney precursor.
RT-PCR analysis showed that the expression of MIXL1, LHX1, and HOXD11 in KPCs increased significantly at days 3, 7, and 12 compared with HUC-iPS at day 0. Furthermore, the expression of MIXL1 was highest when differentiated at day 3, the expression of LHX1 peaked on the seventh day of differentiation, and after 12 days of differentiation, the expression of HOXD11 reached its highest level (Fig. 7C).The expression pattern of these genes in the process of differentiation was similar to their activation during kidney development.
To investigate whether HUC-iPS had higher kidney differentiative capacity, we compared significant activated genes (MIXL1, LHX1, and HOXD11) mRNA expression when cells were differentiated at days 3, 7, and 12 between the HUC-iPS group and the HEF-iPS group. The mRNA expression of LHX1 and HOXD11 was significantly higher when cells were differentiated at days 7 and 12 in the HUC-iPS group (Fig. 7D). Furthermore, the immunofluorescence analysis showed that kidney precursor markers (Pax2, WT1, and Six2) were positively expressed in differentiated cells (derived from HEF-iPS or HUC-iPS) at day 12 (Fig. 7E). These results demonstrated that the kidney precursors were obtained by the defined method, and the differentiation efficiency of kidney precursors in the HUC-iPS group was higher than that in the HEF-iPS group.
Rescue effect of KPCs in an AKI model
To determine whether KPCs could repair renal damage upon AKI, we transplanted KPCs under the renal capsule after IRI (Fig. 8A). Our observations revealed that mice receiving FKPCs (derived from HEF-iPS) or UKPCs (derived from HUC-iPS) transplants showed improved renal function as measured by biochemical parameters, including significantly reduced serum creatinine (163.1 ± 10.94, 149.3 ± 6.49 vs. 211.4 ± 13.76, n = 4) and BUN (81.26 ± 3.36, 79.21 ± 1.73 vs. 94.57 ± 3.75, n = 4) levels, compared with mice receiving HUC transplants (Fig. 8B, C). Histological analyses further revealed lowered levels of tubular dilation and necrosis, reduced number of inflammatory cells, and reduced cellar edema in mice receiving KPCs than in mice receiving HUCs (Fig. 8D, E). Thus, these results indicated that transplantation of KPCs promoted kidney repair after AKI.
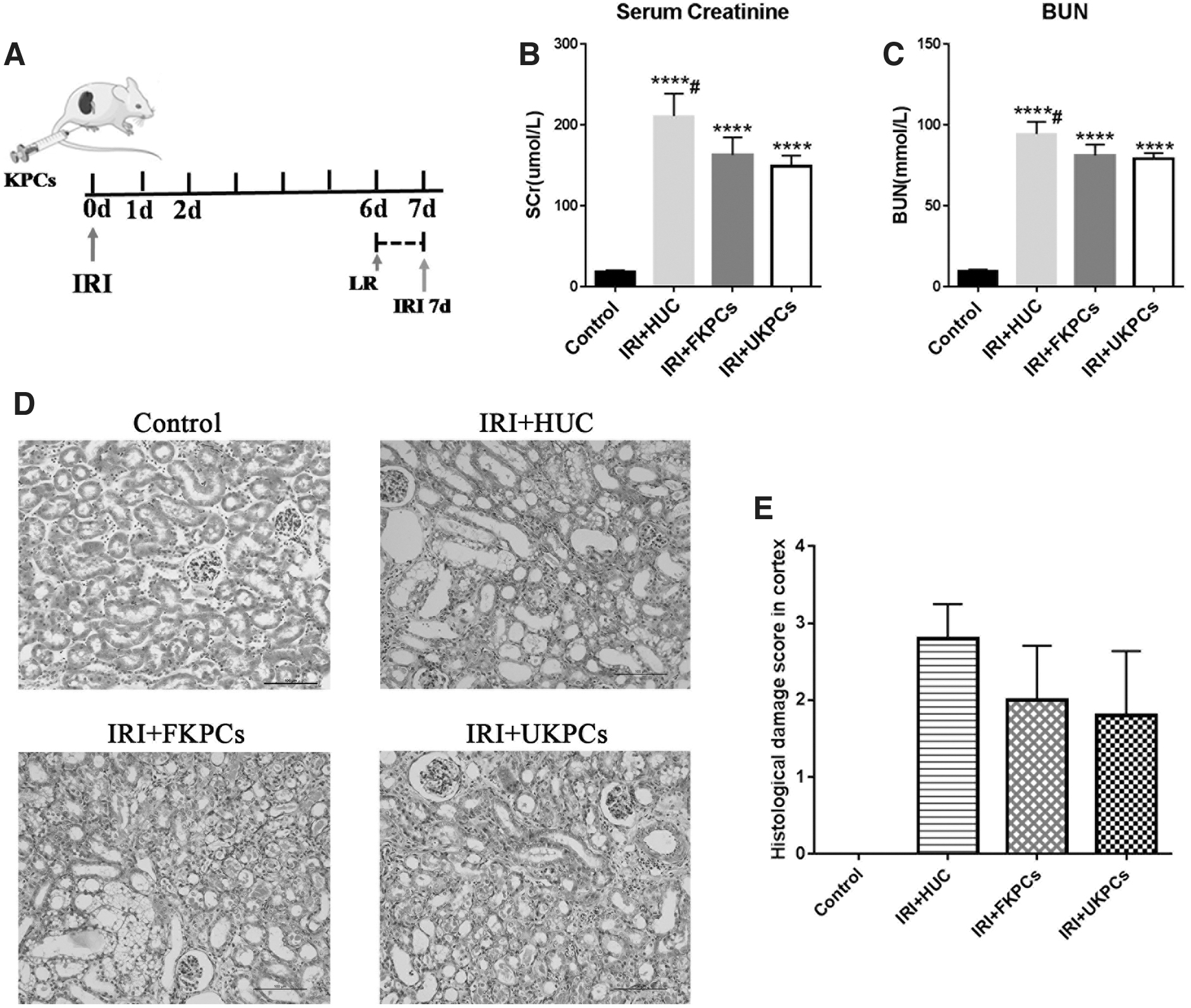
KPC transplants improve kidney function in IRI model.
Discussion
In the process of AKI, the glomerular filtration rate decreases rapidly accompanied by the accumulation of metabolic waste, which often leads to chronic kidney disease (CKD), end-stage kidney disease, and even death (Fan et al., 2017). At the early stage of kidney injury, damaged TECs induce the infiltration of immune cells through damage-associated molecular patterns (Rabb, 2012). Interaction between TECs and immune cells promoted endogenous mechanisms for the repair of the epithelium (Kumar, 2018).
Under the condition of severe injury, endogenous repair was disordered and the remodeling of the extracellular matrix was poor, leading to the progression of renal fibrosis (Goligorsky, 2019). These lines of evidence have suggested that endogenous repair of AKI has played a positive role in kidney recovery. Studies reported that the application of stem cells could promote cell proliferation and migration, as well as inflammatory suppression, which contributed to the repair of the epithelium in AKI models (Tögel et al., 2005).
In ischemia–reperfusion (IR)-induced AKI models, some studies have demonstrated that mesenchymal stem cells (MSCs) inhibit inflammation by downregulating the expression of proinflammatory cytokines (interleukin [IL]-1β, tumor necrosis factor-alpha, and interferon-gamma), and upregulating the expression of anti-inflammatory cytokines IL-10. MSCs also promote cell proliferation and migration by increasing the secretion of cytokines (bFGF and transforming growth factor-alpha). These results imply that MSCs rescue after AKI may effectively ameliorate kidney damage (Tögel et al., 2005). However, MSCs derived from bone marrow are relatively difficult to obtain and have a limited ability to proliferate in vitro, which limits their application.
Through reprogramming the human adult cells, patient-specific iPSCs were generated in large quantities. Moreover, iPSCs are characterized by their proliferation ability and autocrine or paracrine function, making them possible to be the best candidate cells for renal injury rescue.
Lee et al. confirmed that the injection of iPSCs derived from MEFs into an IR-induced AKI rat model can significantly improve renal function by suppressing renal inflammation. They further found that iPSCs could differentiate into a variety of cell types in vitro, but failed to differentiate and replace the damaged TECs during kidney repair (Lee et al., 2012). Similarly, in the study of MSCs for AKI rescue, MSCs also did not directly differentiate into TECs (Burst et al., 2010). It was previously reported that transplanted MSCs could differentiate into endothelial cells and smooth muscle cells, supporting the formation of new blood vessels (Chen et al., 2008).
Li et al. (2016) found that kidney progenitor cells derived from fetuses or iPSCs not only showed robust nephrogenic potential in vivo, but also promoted kidney recovery by paracrine effects in an AKI model. These studies suggested that the origin and nephrogenic potency of stem/progenitor cells might be key factors for differentiation of TECs in the process of kidney repair.
In this study, we obtained urine-derived renal cells from patients with AKI, followed by establishing HUC-iPS, and then differentiated iPS into KPCs. Subsequently, we demonstrated that transplantation of KPCs showed more rescue effects than transplantation of HUCs in the AKI model. We hypothesized that the KPCs might be able to differentiate into TECs in the immune microenvironment through their renal cell-specific gene expression pattern, and replace injured TECs to participate in repair. Meanwhile, the lower rescue effect of HUC transplants was probably due to their lower differentiation potency.
It is well known that urine is a noninvasive cell source for generating pluripotent cells (Zhou et al., 2011). In our study, we isolated urine-derived renal cells with epithelioid morphology from patients with AKI, and established patient-specific iPS, paralleling with previous findings (Zhou et al., 2012). It was beneficial to minimize immune rejection for autologous cell therapy in AKI. Owing to AKI often leading to CKD, cell therapy has become an effective way to reduce kidney damage and inhibit the transition of AKI to CKD by regulating anti-inflammatory and promoting endogenous repair during the process of AKI. Our differentiation study confirmed that the nephrogenic potential of HUC-iPSCs was higher than that of HEF-iPSCs in vitro. UKPCs showed more rescue effects than FKPCs in vivo. Based on our findings, UKPCs might be the best candidate cells for cell therapy in AKI.
Furthermore, patient-specific iPSCs are considered to be the best cell model of renal genetic diseases and drug screening in vitro (O'Neill and Ricardo, 2013; Robinton and Daley, 2012). The HUC-iPS established in our study, carrying disease-related genotypes, could be a model for patient-specific AKI. Recent studies have revealed that kidney organoids derived from human iPSCs have a variety of structures, such as glomeruli and renal tubules (Homan et al., 2019; Van den Berg et al., 2018). In our next study, HUC-iPS will also be used to generate kidney organoids for the study of kidney regeneration.
In summary, our study was the first to obtain urine-derived renal cells from patients with AKI, followed by establishing the HUC-iPS line. We discovered that urine-derived renal cells with epithelioid morphology had expression of renal TEC-specific markers and renal development-related genes. Through transfection with TET-ON and induction, HUC-iPS exhibited many pluripotent characteristics, including AP staining and immunofluorescence for ESC pluripotency markers, and higher iPS reprogramming efficiency than HEF-iPS. Notably, HUC-iPS could be induced to undergo directional differentiation of UKPCs under defined conditions, and UKPC transplants could reduce kidney injury after AKI. We successfully established HUC-iPS from patients with AKI and provided unique insights for stem cell therapy in AKI.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.