
Abstract
Through comparative gene mapping, NICE-3, which is closely linked to tropomyosin 3 in human chromosome 1, was selected to be investigated as a new candidate gene associated with the muscle development in pigs. This gene was sequenced, chromosome mapped, expression analyzed, subcellularly localized, and promoter activity analyzed. After screening and sequencing, porcine NICE-3 was found in a bacterial artificial chromosome clone containing tropomyosin 3. Quantitative reverse transcription–polymerase chain reaction revealed that NICE-3 mRNA was widely expressed, with highest expression levels in longissimus dorsi muscles, followed by heart, biceps femoris, liver, kidney, back fat, and lowest expression levels in spleen, brain, lymph, lung, stomach, and small and large intestines. Fluorescence and confocal microscopy assay demonstrated that the fusion protein, GFP-NICE-3, was distributed throughout the cytoplasm, including the plasma membrane. NICE-3 was mapped to Sus scrofa chromosome 4, in a region of conserved synteny with human chromosome 1, where the homologous human gene is localized. Results of dual reporter gene assays and mutation experiments combined with electrophoresis mobility shift assays showed that the retinoid X receptor might be an important transcription factor affecting the promoter activity of this gene.
Introduction
NICE-3, also known as chromosome 1 open reading frame 43 (C1orf43), is located on region q21.2 of human chromosome 1 (Hsap1) (Zhao et al., 2007) and is linked closely to the gene tropomyosin 3 (TPM3) (Rosati et al., 2006). The TPM3 gene is associated with nemaline myopathy (Penisson-Besnier et al., 2007; Ilkovski et al., 2008; Lehtokari et al., 2008) and congenital muscle fiber type disproportions (Clarke et al., 2008). NICE-3 belongs to the epidermal differentiation complex (Marenholz et al., 2001; Perez-Plasencia et al., 2005), and three alternatively spliced cDNA sequences of this gene have been obtained (Marenholz et al., 2001). There is a predicted peroxisome proliferator response element (PPRE) in conserved elements upstream of the transcription start site. Peroxisome proliferator–activated receptors, binding PPREs in regulatory regions of their target genes, are a family of nuclear receptors that not only broadly regulate nutrient metabolism and energy homeostasis, but also act as cellular sensors of fatty acids and fatty acid derivatives (Lemay and Hwang, 2006).
It has been reported that NICE-3 might play an important role in the maintenance and/or differentiation of human hematopoietic stem/progenitor cells (Zhao et al., 2007). It was not destabilized upon p38 inhibition in lipopolysaccharide-treated THP-1 cells (Frevel et al., 2003) but showed significant upregulation among all primary and metastatic fibrolamellar carcinomas studied (Kannangai et al., 2007). In addition, it was differentially expressed between ductal carcinoma in situ and invasive ductal carcinomas in human breast cancer progression (Abba et al., 2004). Its expression level differed significantly between survivors and deceased patients with lymph node–negative breast cancers (Karlsson et al., 2008), but was not strongly associated with the risk of developing metastases (Mosley, 2007). The corresponding protein product was predicted to be distributed in the cell membrane, or integral to it (Zhang, 2006), or in the cytoplasm and mitochondria (
Materials and Methods
Molecular cloning of the porcine NICE-3 gene
The human NICE-3 mRNA sequence (GenBank accession no. NM_001098616) was used as a probe to carry out BLAST searching in the porcine-expressed sequence tag database (
The porcine NICE-3 mRNA sequence obtained was separated into seven exons after being aligned with the human NICE-3 genome sequence, and primers were designed to obtain a sequence of introns 1, 3, and 5 (NICE-In(1)F/R, NICE-In(3)F/R, and NICE-In(5)F/R; Supplemental Table S1). Primers spanning the exons and introns were designed (NICE-BAC(1)F/R and NICE-BAC(2)F/R; Supplemental Table S1) to screen a porcine bacterial artificial chromosome (BAC) library to obtain the porcine NICE-3 promoter sequence.
Nested PCR primers with the same reverse primer were designed (NICE-TSS(1)F-(6)F and NICE-TSSR; Supplemental Table S1) to confirm the transcriptional start site (TSS) according to Lee et al. (2005), and the nucleotide immediately upstream of the translation initiation codon (ATG) was denoted as −1.
Chromosome mapping by IMpRH
The INRA (Institut National de la Recherche Agronomique)–University of Minnesota's 7000-rad radiation hybrid panel (IMpRH) was used for chromosome mapping of the porcine NICE-3 gene with porcine-specific primers (NICE-RHF/R; Supplemental Table S1). Each PCR run consisted of 5 min at 95°C; 35 cycles of 20 s at 95°C, 20 s at 57.9°C, and 20 s at 72°C; and a final 1 min extension at 72°C. Data were analyzed using the IMpRH mapping tool (
Quantitative reverse transcription–PCR analysis of gene expression patterns
To evaluate NICE-3 as a new candidate gene associated with ADG and BFT, the expression level analysis of NICE-3 in longissimus dorsi muscle, biceps femoris, back fat, and other organs was an important method to study its function. Thirteen tissues (longissimus dorsi muscle, biceps femoris, back fat, heart, liver, spleen, lung, kidney, stomach, brain, lymph, large intestine, and small intestine) were obtained from three female mature Lantang pigs (a kind of Chinese indigenous pig breed, 180 days, about 70.4 kg). Total RNA was extracted from the tissues using the Trizol reagent (Invitrogen, Carlsbad, CA) and treated with RNase-free DNase (MBI Fermentas, St. Leon-Rot, Germany). RNA quality was assessed by agarose gel electrophoresis and the ND-1000 Spectrophotometer (NanoDrop Technologies, Wilmington, DE) and was discarded if the 260/280 ratio was not between 1.8 and 2.1. The RNA was then subjected to reverse transcription (RT) into cDNA using M-MLV reverse transcriptase (Promega, Madison, WI). The housekeeping beta actin gene (ACTB) was selected as an internal control for normalization as it was consistently expressed in all tissues of this assay. A negative RT control with primers for ACTB was performed to test if products were derived from DNA. The quantitative PCR (qPCR) conditions consisted of a single cycle of 95°C for 30 s, followed by 40 cycles of 95°C for 5 s, 64°C for 10 s, and 72°C for 15 s, and then a final cycle of 95°C for 10 s and 65°C for 1 min. The reaction was performed with SYBR Premix Ex Taq (TaKaRa Bio, Dalian, P.R. China), using a LightCycler 480 Real-Time PCR System (Roche, Basel, Switzerland) (corresponding primers NICE-RT-F/R and ACTB-F/R; Supplemental Table S1). The efficiency of the qPCR assays was calculated by the LightCycler 480 software program. In this assay, the efficiency of NICE-3 and ACTB gene primers was 2.007 and 1.975, respectively. Relative expression was calculated as a ratio between expression of NICE-3 and expression of the ACTB in the same cDNA sample, according to the following equation: ratio =(ET)CpT(c) − CpT(s) ×(ER)CpR(s) − CpR(c). In this equation, ET and ER represent the efficiencies of qPCR from NICE-3 and ACTB, respectively, CpT(c) and CpT(s) represent the crossing points of the target gene for the calibrator and the samples, respectively, and CpR(s) and CpR(c) represent the crossing points of the reference gene for the samples and the calibrator, respectively.
Subcellular localization of the porcine NICE-3 protein in pig kidney epithelial cells (PK15)
The CDS, encoding porcine NICE-3 protein, was amplified by PCR using the NICE-C-F/R primers (Supplemental Table S1) and subcloned into the HindIII–MluI site of the pEGFP-C1 green fluorescent protein (GFP)–expressing vector (Clontech–Takara Bio, Mountain View, CA) to yield a mammalian expression plasmid, named pNICE-GFP. After sequence verification, the plasmids were extracted using an Endo-free Plasmid Mini Kit (Omega Bio-Tek, Norcross, GA) and transferred into PK15 using Lipofectamine 2000 reagent (Invitrogen) according to the manufacturer's instructions. PK15 cells were cultured in Dulbecco's modified Eagle's medium supplemented with 10% fetal bovine serum (Gibco–Invitrogen, Carlsbad, CA), 100 U/mL penicillin, and 0.1 mg/mL streptomycin under humidified air containing 5% CO2 at 37°C and seeded onto the cover slips in six-well plates.
Twenty-four hours after transfection, the cells were incubated at 37°C for 30 min in growth medium containing 200 nM MitoTracker Orange (Molecular Probes, Eugene, OR) for mitochondrial labeling, 200 nM Golgi-Tracker Red (Beyotime Institute of Biotechnology, Haimen, China) for Golgi apparatus labeling, or 200 nM ER-Tracker Red (Molecular Probes) for endoplasmic reticulum labeling. After washing with phosphate-buffered saline, the cells were incubated at 37°C for 10 min with 10 μM Hoechst 33342. Then the slides were fixed with formaldehyde, mounted, sealed, and analyzed using an Olympus FluoView FV1000 Confocal Microscope (Olympus, Tokyo, Japan). The results were analyzed using FV1000 Viewer software.
Reporter gene assay and site-directed mutagenesis
A reporter gene assay was carried out to study the porcine NICE-3 promoter. Nested PCR primers with the same reverse primer were designed (NICE-Prom(0)F-(6)F and NICE-PromR; Supplemental Table S1), and the PCR products were subcloned into pGL3-basic vector (Promega) after being digested with MluI and XhoI. After sequence verification, the plasmids were extracted with an Endo-free Plasmid Mini Kit and transferred into C2C12 cells with Lipofectamine 2000 reagent. C2C12 cells were cultured in almost the same conditions as for PK15 cells, but in 24-well plates rather than 6-well plates. The pRL-TK vector was used as the internal control for normalization, and the ratios of reporter gene vector to pRL-TK were 80:1 or 60:1. A pGL3-basic vector was used as the external control.
Twenty-four hours after transfection, the cells were lysed, and firefly luciferase and Renilla luciferase activities were measured using the Dual-Luciferase Reporter Assay System (Promega) according to the manufacturer's instructions. After analyzing the results, fragments with higher activity were selected, and the 5′ sequence was analyzed using the Transcription Element Search System (TESS) web tool (
Electrophoresis mobility shift assay
Electrophoresis mobility shift assay (EMSA) was done using a commercial LightShift Chemiluminescent EMSA kit, according to the manufacturer's instructions (Viagene Biotech, Ningbo, P.R. China). Nuclear extracts for EMSA were prepared according to the manufacturer's instructions (Viagene Biotech). The corresponding probe (NICE-EMSAF/R; Supplemental Table S1) was generated as follows: double-stranded oligonucleotides were end labeled with γ-32P ATP and T4 polynucleotide kinase. In competition experiments, unlabeled competitors containing intact or mutant consensus sequences (NICE-MutationF/R; Supplemental Table S1) were added in 10- or 50-fold excess. Probes were incubated with nuclear extracts from C2C12 cells for 15 min on ice, and the DNA–protein complexes were resolved by electrophoresis on 4% (w/v) polyacrylamide gels.
Results
Molecular cloning and sequence analysis
A 925 bp fragment of the porcine NICE-3 cDNA sequence (GenBank accession no. EU814887) was obtained and used to determine the exon/intron organization. The potential promoter region (GenBank accession no. EU814889), as well as genomic sequence, was obtained from the pig BAC clone (no. 303L14) containing the NICE-3 gene. In the TSS experiment, all forward primers from NICE-TSS(3)F to NICE-TSS(6)F yielded PCR products, whereas primers NICE-TSS(1)F and NICE-TSS(2)F did not (data not shown). This indicated that the TSS might be mapped to region −388 to −347 nucleotide located between the 3′ ends of primers NICE-TSS(2)F and NICE-TSS(3)F.
In total, 12,625 bp of genomic sequence representing about 2067 bp of the 5′-flanking region, about 354 bp of the 5′-untranslated region, 762 bp of the coding region, and 342 bp of the 3′-untranslated region of the porcine NICE-3 gene were obtained. The exon–intron boundaries and respective exon and intron sizes are shown in Table 1. Seven exons of the porcine NICE-3 were confirmed with size ranging from 59 to 538 bp, and the exon structure is similar to that of the human C1orf43 gene transcript variant 3. The sizes of the introns found varied substantially, ranging from 86 to about 3935 bp, and all exon/intron junctions were in accordance with the universal RNA splice criteria (GT-AG rule). The NICE-3 protein is highly conserved in mammals, and the porcine NICE-3 protein sequence shows identities of 98% to equine (GenBank accession no. XP_001496686.1), dog (GenBank XP_852286.1), macaque (GenBank BAE91701.1), and bovine (GenBank NP_001015666.1) sequences, and 97% to human (GenBank NP_001092086.1), rat (GenBank AAH98673.1), and mouse (GenBank NP_080038.1) sequences. A conserved NICE-3 domain was found in the porcine NICE-3 protein sequence (
Chromosome assignment
Using the IMpRH, porcine NICE-3 was mapped to SSC4 and closely linked to SW512 (limit of detection [LOD] score 9.41, 44cR) with 31% retention frequency. The gene was also linked to SW270 (LOD score 6.41, 60cR), S0214 (LOD score 6.02, 62cR), and SW286 (LOD score 5.95, 65cR) in this region. The conserved syntenic region in human is on chromosome 1q21.2, where the human NICE-3 is located. Based on the Radiation Hybrid distance (cR) and reference map (
Expression analysis of porcine NICE-3 gene
qPCR analysis indicated that the porcine NICE-3 gene was ubiquitously expressed in all the tissues examined, with the highest expression in longissimus dorsi muscle, moderate expression in heart, biceps femoris, liver, kidney and back fat, and the lowest amount expression in spleen, brain, lymph, lung, stomach, and small and large intestines (Fig. 1).
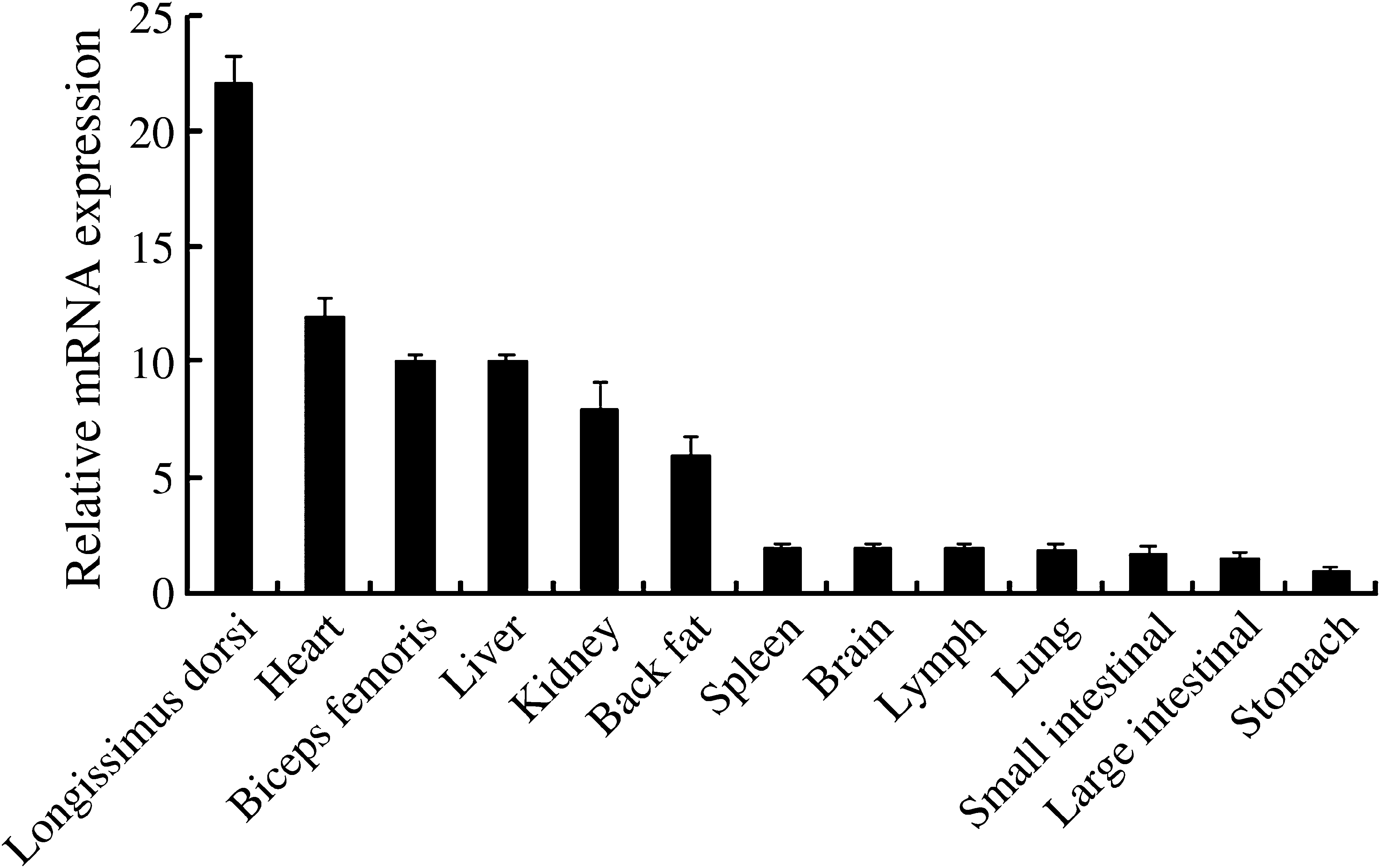
Analysis of porcine NICE-3 gene expression in different tissues using quantitative PCR on RNAs from different tissues including longissimus dorsi muscle, biceps femoris, back fat, heart, liver, spleen, lung, kidney, stomach, brain, lymph, large intestine, and small intestine. The mRNA levels of NICE-3 were normalized to those of the housekeeping gene ACTB and indicated in the respective porcine tissues. Error bars represent the standard deviation (n = 3).
Subcellular distribution
Confocal analysis showed that the fusion proteins were mainly distributed throughout the cytoplasm, including the plasma membrane (Fig. 2), and green fluorescence was detected throughout the control cells transfected with the pEGFP-c1 vector alone (data not shown).
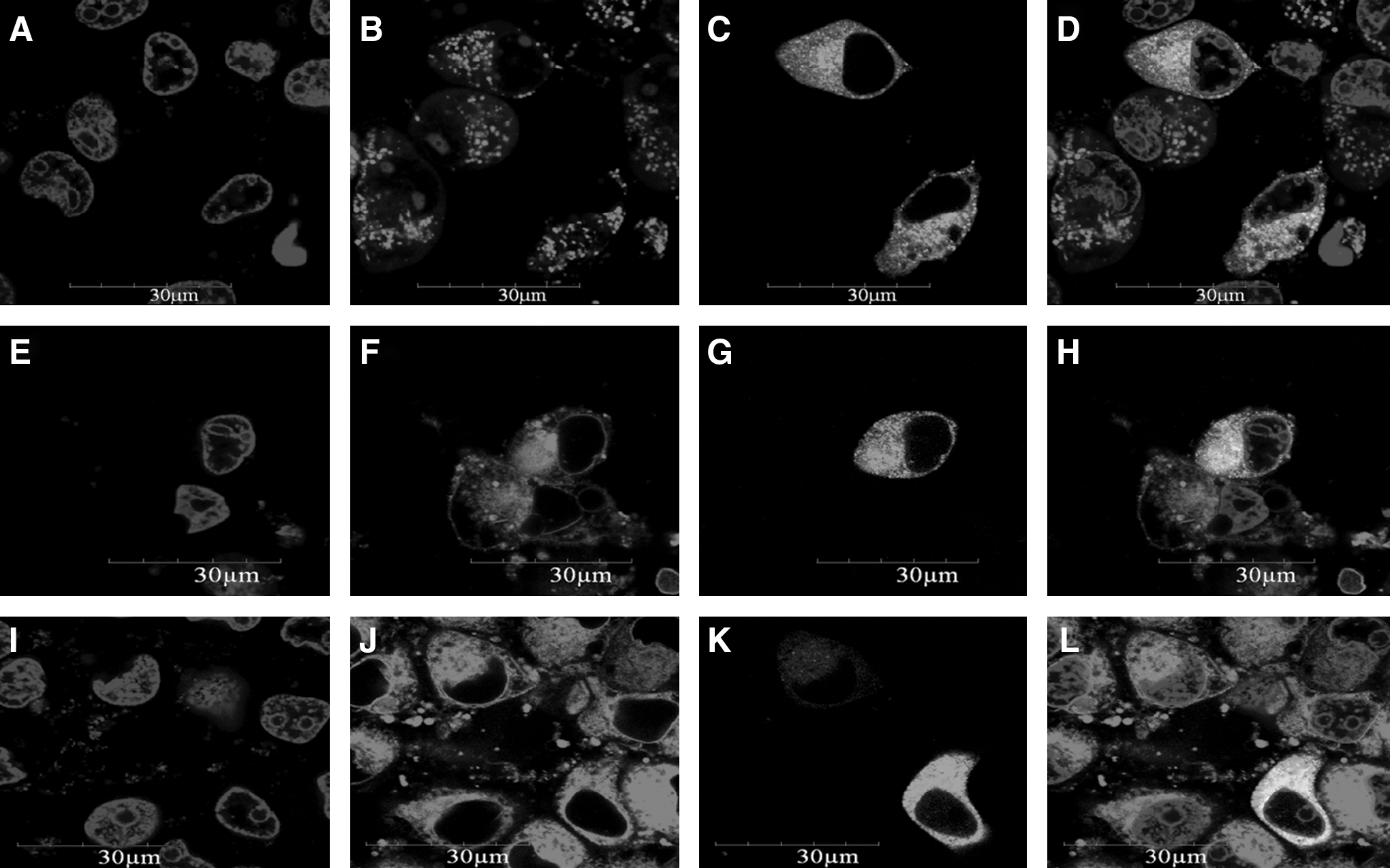
Cellular localization of the pNICE-GFP fusion protein in PK15 cells. Recombinant plasmids were transfected into PK15 cells using Lipofectamine 2000. Nuclei stained with Hoechst 33342 (
Transcriptional regulation of the NICE-3 gene
Transcriptional activity of the NICE-3 gene was examined using the transient transfection method. To determine the regulatory elements responsible for NICE-3 gene expression, a series of deletion constructs of the 5′-flanking region of the gene were generated. These deletion constructs were transfected transiently into C2C12 cells, and the levels of luciferase activity were determined. Figure 3A shows that deletions up to positions −1649 and −1341 resulted in significant changes in promoter activity, relative to the luciferase activity of the longest NICE-3 promoter reporter construct (nucleotides −2421 to −122). The promoter activity was lost or enhanced in constructs containing nucleotides −1649 to −122 or −1341 to −122, respectively. After analyzing the sequence from nucleotides −1341 to −1077, two potential transcription factor binding sites were selected to be mutated, and these were scored 14 and 16 by TESS. One was a retinoid X receptor (RXR) element-binding site, at position −1218 to −1212, and the corresponding mutated plasmid was named M1. Another was a CCAAT/enhancer-binding protein element-binding site, at position −1150 to −1143, and the corresponding mutated plasmid was named M2. Promoter activity of M1 was significantly less than M2 (Fig. 3B). In an attempt to identify transcription factors that have the potential to interact with the promoter region of NICE-3, a synthetic double-stranded DNA probe for RXR was subjected to a gel mobility shift assay. As shown in Figure 4, DNA–protein complexes were formed with the nuclear extracts from C2C12 cells. The specificities of these complexes for the sequence were shown in a competition experiment, in which the complexes were competed by a 10- or 50-fold excess of unlabeled wild-type probe (C-RXR) and by a 10- or 50-fold excess of unlabeled mutation probe (M-RXR). The results revealed that the difference between wild RXR and the mutated RXR affected their binding with RXR, so the mutated nucleotide must be the core nucleotide of the RXR element-binding site, and the interaction of RXR with a RXR element-binding site was necessary for basal constitutive promoter activation.
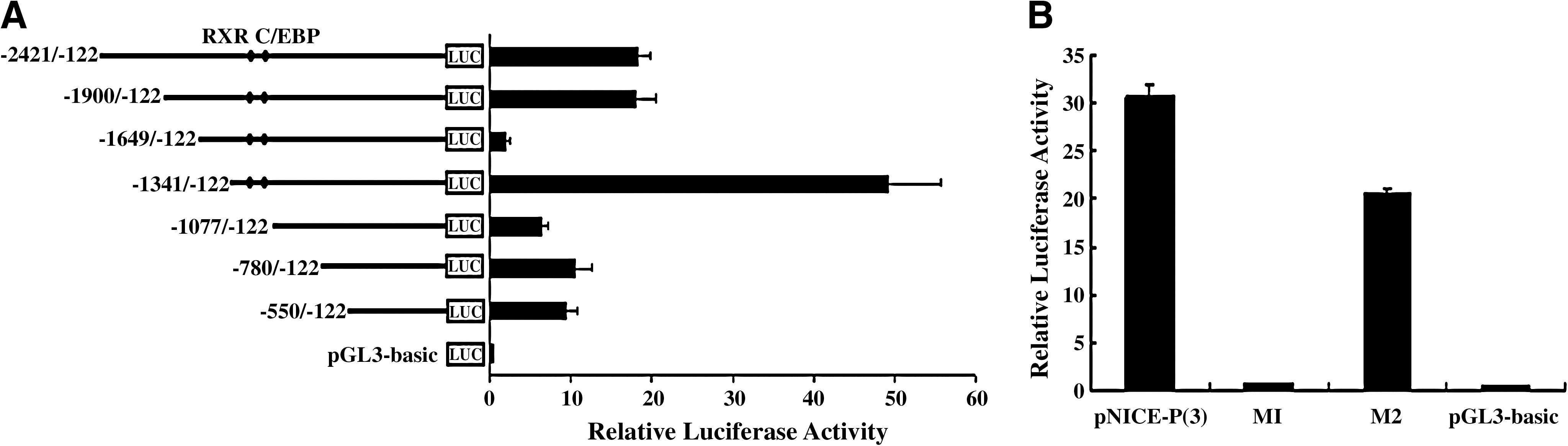
Reporter gene analysis of porcine NICE-3 gene promoters in C2C12 cells. (
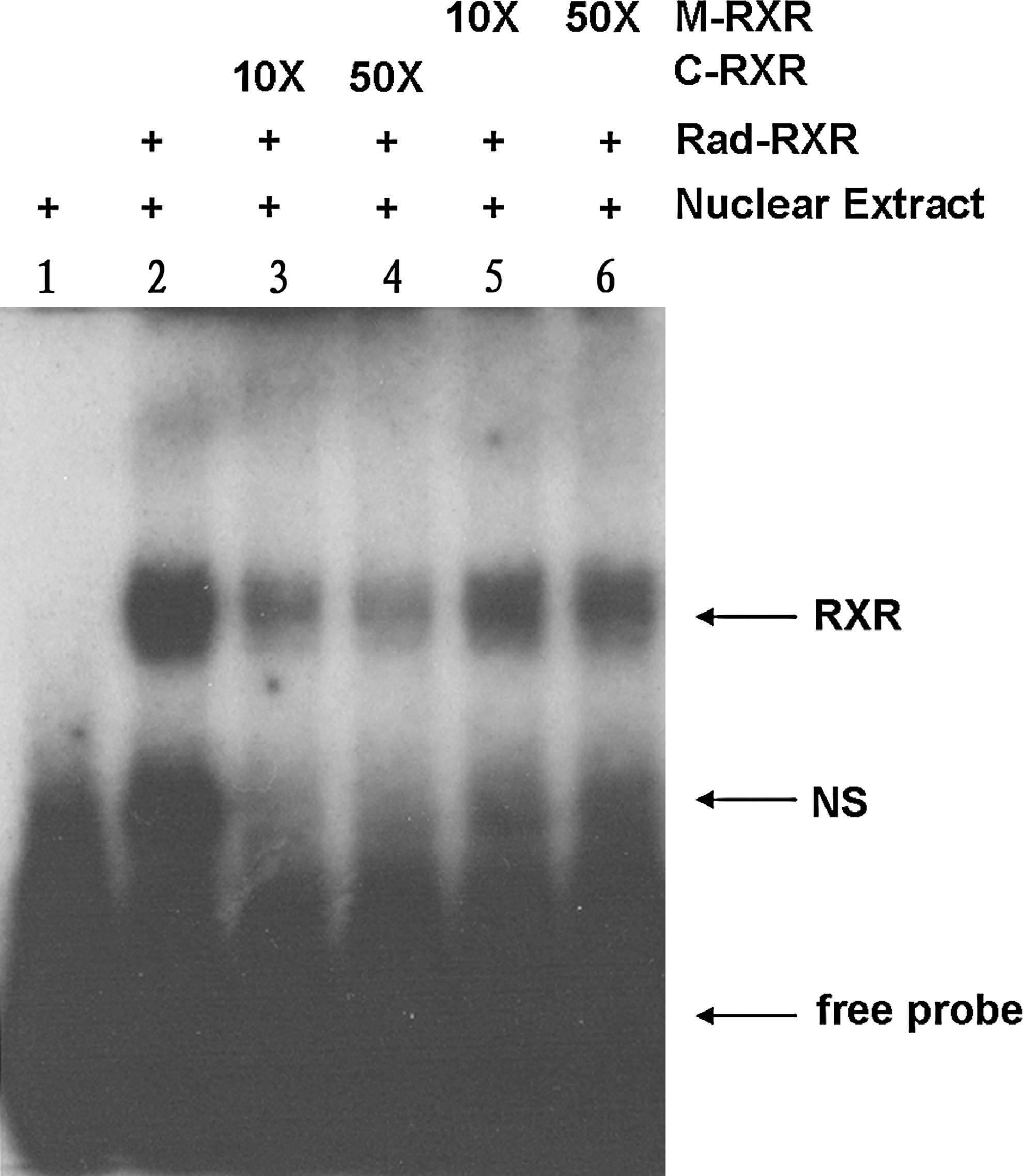
Specific DNA–protein binding to putative RXR oligonucleotides in porcine NICE-3 promoter. Radiolabeled probe (Rad-RXR) was incubated, or not, with nuclear extracts of C2C12 cells (lane 2 and 1). Competitive assays were performed using 10- and 50-fold molar excesses of unlabeled oligonucleotides (C-RXR in lanes 3 and 4, and M-RXR in lanes 5 and 6). Specific protein–DNA complexes (RXR) are indicated by arrows. NS, nonspecific.
Discussion
The DNA sequence of the porcine NICE-3 gene was obtained for the first time by sequencing a BAC clone, and the gene was mapped to SSC4, closely linked to the microsatellite marker SW512. In addition, this BAC clone (no. 303L14) also contains porcine TPM3 gene that is associated with normal development of muscle (Clarke et al., 2008; Lehtokari et al., 2008). Interestingly, porcine TPM3 is also closely linked to marker SW512 with a LOD score of 7.48 using the IMpRH in our previous work (Zhu, 2006). SSC4 is the first porcine chromosome analyzed for QTLs (Andersson et al., 1994), and many QTLs correlated with pork meat quality and other traits have been mapped on this chromosome. The marker SW512 is related with QTLs for ADG, BFT, eye muscle area, linoleic fatty acid content, and oleic fatty acid contents (Rohrer et al., 1994; Andersson-Eklund et al., 1998; Walling et al., 1998; Malek et al., 2001). The qPCR results showed that porcine NICE-3 mRNA was most highly expressed in the longissimus dorsi muscle. All these results support this hypothesis that the porcine NICE-3 gene could be involved in muscle development.
There is a predicted PPRE in the promoter of porcine NICE-3 gene, just as in the human NICE-3 gene, but it does not play an important role in transcriptional regulation. Results of promoter activity analysis indicate that potential positive regulatory elements seem to be located in the region from nucleotides −1341 to −1077, which is essential for a high level of expression of the NICE-3 gene, and potential negative regulatory elements seem to be located in the region from nucleotides −1649 to −1341, which is essential for low-level expression of the NICE-3 gene. Site-directed mutagenesis and EMSA indicated that transcription factor RXR might play an important role in transcriptional regulation of porcine NICE-3 gene. RXR is a unique member of the family of nuclear hormone receptor proteins, for it has the ability to form many heterodimers as well as homodimers. The family of RXRs acts in a number of different nuclear receptor–based signal transduction pathways, including cellular and organ differentiation, apoptosis, and many metabolic processes (Szanto et al., 2004; Desvergne, 2007). Moreover, RXRs, by forming heterodimers with other nuclear receptors, might play important roles in epidermal keratinocyte proliferation and differentiation (Feng et al., 1997; Li et al., 2000). Meanwhile, the NICE-3 protein family consists of several eukaryotic NICE-3 and related proteins. The gene coding for NICE-3 is part of the epidermal differentiation complex that comprises a large number of genes that are of crucial importance for the maturation of the human epidermis (Marenholz et al., 2001), while the function of NICE-3 is still unknown. These observations indicate that porcine NICE-3 gene might play some role in epidermal proliferation and differentiation.
In conclusion, porcine NICE-3 was sequenced; its chromosomal localization and tissue distribution were identified. Tissue expression pattern of this gene was analyzed, as well as mechanisms of transcriptional regulation. Based on the results, we get a first insight of potential biological functions of the gene, and data presented in this study will lead to a better understanding of the biological roles of the NICE-3 gene and will help further study.
Footnotes
Acknowledgments
This research was funded by Major State Basic Research Development Program, China (2004CB117506) and the National Natural Science Foundation of China-Guangdong Key Project (u0731003). The authors are grateful to Dr. Ning Li, Dr. XiaoXiang Hu, and Li Min Ren of state key laboratories for AgroBiotechnology of CAU for helps in screening pig BACs.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.