
Abstract
Single-nucleotide polymorphisms (SNPs) are single-nucleotide sequence variations between individuals. Two missense SNPs are present in the human undifferentiated embryonic-cell transcription factor 1 (UTF1) gene and their consequences for UTF1 function are investigated in this study. Expression of the UTF1 gene is restricted to pluripotent cells and UTF1 is a chromatin-associated protein with core histone-like properties. UTF1 further acts as a transcriptional repressor and is required for proper differentiation of pluripotent cells. Two missense mutations in UTF1 are reported: rs11599284, which results in a glycine to an arginine change at amino acid 73, and rs4480453, resulting in a leucine to methionine change at amino acid 275. To study the effects of these two SNPs, P19CL6 mouse embryonic carcinoma cells stably expressing eGFP-hUTF1 constructs containing either one or both SNPs were generated. The single and double SNPs did not alter the localization or transcriptional repressor activity of the protein. Further, the single SNPs did not alter the chromatin association and mobility of hUTF1. However, the double mutant, G73R/L275M, demonstrated a decreased chromatin association, indicating a degree of protein malfunction.
Introduction
UTF1 is expressed in the inner cell mass, primitive ectoderm, and early primitive streak stage of the developing mouse embryo (Okuda et al., 1998). During development, its expression is maintained in the primordial germ cells of the developing embryo (Chuva de Sousa Lopes et al., 2005), and in adults, its expression is restricted to the gonads (Okuda et al., 1998; Chuva de Sousa Lopes et al., 2005; Kristensen et al., 2008; van Bragt et al., 2008). UTF1 is a nuclear protein, excluded from the nucleoli, and it associates with DNA during all stages of the cell cycle, including mitosis (van den Boom et al., 2007; Kooistra et al., 2009). UTF1 is a stable chromatin-associated transcriptional repressor with core histone-like properties (van den Boom et al., 2007; Kooistra et al., 2009). In addition, the requirement of UTF1 to maintain the proliferation rate of embryonic carcinoma and ES cells (Nishimoto et al., 2005; Li et al., 2007; van den Boom et al., 2007) might explain its expression in germ cell neoplasms (Kristensen et al., 2008) and teratoma formation-promoting activity (Nishimoto et al., 2005).
The key domains that are essential for the function of UTF1 are two evolutionary conserved domains (CDs), CD1 and CD2. CD1, which shares high homology to Myb/SANT domains, is responsible for proper nuclear targeting of the protein because deletion of CD1 leads to cytoplasmic mislocalization (Kooistra et al., 2009). CD2, which contains a putative leucine zipper, is responsible for stable chromatin association of the protein (Fukushima et al. 1998; van den Boom et al., 2007; Kooistra et al., 2009). In human UTF1, the CD2 domain is required for both repressor activity and stable chromatin interaction (Kooistra et al., 2009).
Single-nucleotide polymorphisms (SNPs) are major contributors to DNA polymorphisms in a number of eukaryotic species. SNPs that are localized in the promoter regions (Song et al., 1996; Wilson et al., 1997; Knight et al., 1999; Liu et al., 1999; Shin et al., 2000; Bream et al., 2002; Garcia-Barcelo et al., 2005; Szalai et al., 2005; Fushan et al., 2009) and regulatory elements of genes, affecting their expression, are reported, as well as missense or nonsense mutations affecting the function of the encoded protein (Kostrikis et al., 1998; Martin et al., 1998; Winkler et al., 1998; Bream et al., 1999; Knight et al., 1999; El-Omar et al., 2000; Collin et al., 2008). For the UTF1 gene, SNPs are reported in the noncoding regions as well as two missense SNPs in the coding region of the UTF1 gene. These missense SNPs are localized in the CD1 (G73R) and CD2 (L275M) domains, respectively. The heterozygosity is 0.087 for G73R and unknown for L275M. We hypothesized that these missense SNPs may result in mislocalization or reduced repressor activity and/or chromatin association of human UTF1 (hUTF1) protein.
The aim of this study was to determine the functionality of UTF1 variants G73R, L275M, and G73R/L275M. The effect of these SNPs on UTF1 repressor activity was determined in reporter assays. The subcellular localization and chromatin association of hUTF1 variants were determined using immunofluorescence and subnuclear fractionation experiments in combination with strip-fluorescence recovery after photobleaching (FRAP) assays.
Materials and Methods
Constructs
eGFP-HA-hUTF1-G73R and eGFP-HA-hUTF1-L275M were generated using the Stratagene Quickchange II XL site-directed mutagenesis kit with peGFP-HA-hUTF1 (Kooistra et al., 2009) as a template and primers F 5′CGGAGCTGCTG CTGAGGACGCTGCTGCAACC 3′ and R 5′GGTTGCAGCA GCGTCCTCAGCAGCAGCTCCG 3′ (G73R) and F 5′GCCGC GCCCCCGTCGATGAACACCGCCCTGC 3′ and R 5′ GCA GGGCGGTGTTCATCGACGGGGGCGCGGC 3′ (L275M), respectively. eGFP-HA-hUTF1-G73R/L275M was generated by transferring a L275M-containing DraIII/BbsI fragment to the eGFP-HA-hUTF1-G73R plasmid. pSG424-hUTF1 G73R, pSG424-hUTF1 L275M, and pSG424-hUTF1 G73R/L275M were generated by ligation of the SNP-containing BstXI/MunI fragments from the respective eGFP-HA-hUTF1 plasmids into pSG424-hUTF1. All SNPs were confirmed by sequencing.
Cell culture and transfections
P19CL6 cells were grown in α-MEM (Invitrogen, Breda, The Netherlands) supplemented with 10% FCS (PAA, Cölbe, Germany), 100 U/mL penicillin (Invitrogen), and 100 μg/mL streptomycin (Invitrogen) at 37°C and 5% CO2. HepG2 cells were grown in DMEM supplemented with 10% FCS (PAA), nonessential amino acids (Invitrogen), 100 U/mL penicillin (Invitrogen), and 100 μg/mL streptomycin (Invitrogen) and maintained at 37°C and 5% CO2. To generate stable lines, P19CL6 cells (75,000) were transfected with 5 μg DNA (eGFP-HA-hUTF1, eGFP-HA-G73R, eGFP-HA-L275M, or eGFP-HA-G73R/L275M) using calcium phosphate coprecipitation. Selection was performed using 600 μg/mL G418. Clones were selected based on GFP expression using immunoblotting. Luciferase reporter assays were performed as described previously (van den Boom et al., 2007; Kooistra et al., 2009).
Microscopy and FRAP
For fluorescence microscopy, 200,000 P19CL6 cells were cultured on glass coverslips and fixed in 2% paraformaldehyde in PBS for 10 min at room temperature. Cells were permeabilized with 0.1% Triton X-100 in PBS and counterstained with DAPI. Images were made using a fluorescent microscope (Axiophot; Carl Zeiss MicroImaging, Sliedrecht, The Netherlands) with a plan-NEOFLUAR 100 × NA 0.70 lens. FRAP experiments were performed as reported previously (van den Boom et al., 2007; Kooistra et al., 2009).
Western blotting and subnuclear fractionation
Western blotting and subnuclear fractionation were performed as described previously (Citterio et al., 2004; van den Boom et al., 2007). Fractions were analyzed using the following primary antibodies: hUTF1 (clone 5G10.2, MAB 4337; Millipore, Amsterdam, The Netherlands), mUTF1 (Eurogentec) (van den Boom et al., 2007), and H2A (acidic patch; Upstate Biotechnology, Amsterdam, The Netherlands). Secondary antibodies used were goat anti-mouse IgG-HRP (Santa Cruz Biotechnology, Santa Cruz, CA), donkey anti-goat IgG-HRP (Santa Cruz Biotechnology), and donkey anti-rabbit IgG-HRP (GE Healthcare, Hoevelaken, The Netherlands).
Results
Missense SNPs in hUTF1
Two biallelic missense SNPs, G73R and L275M, were found in the hUTF1 gene corresponding to entries rs11599284 and rs4480453 in the National Center for Biotechnology Information (NCBI) SNP database (
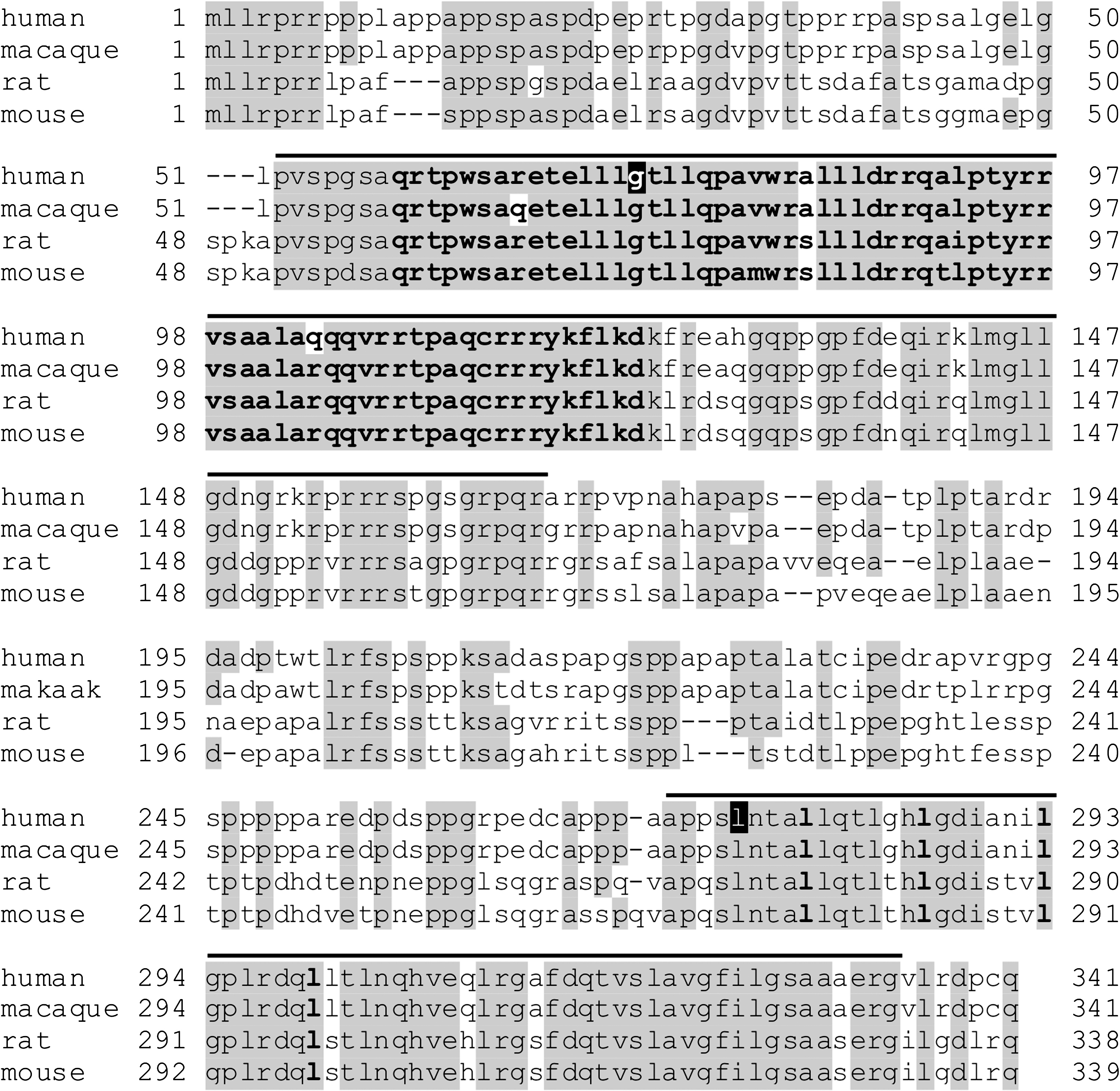
Sequence alignment of undifferentiated embryonic cell transcription factor 1 (UTF1) proteins. Comparison of the amino acid sequence between human, macaque, rat, and mouse UTF1. Amino acids identical in at least three species are boxed in gray. Conserved domain (CD) 1 and CD2 are indicated by lines. Within CD1, the Myb/SANT domain is indicated by bold residues. The first amino acid residues within the seven amino acid repeats of a coiled-coil leucine zipper motif in CD2 are indicated in bold. The single-nucleotide polymorphisms (SNPs) in human UTF1 at positions 73 and 275 are indicated by black boxes.
Subcellular localization of GFP-hUTF1 SNPs
Previously, we have reported that both mUTF1 and hUTF1 associate with DNA during all stages of the cell cycle, including mitosis (van den Boom et al., 2007; Kooistra et al., 2009). To assess if hUTF1 SNPs resulted in mislocalization of the protein, P19CL6 cells stably expressing GFP-hUTF1 and GFP-hUTF1 SNPs were generated and analyzed using fluorescence microscopy. GFP localization indicated that the mutations, either alone or in combination, had a similar nuclear localization as wild-type hUTF1 (Fig. 2). Nonhomogeneous localization of GFP-hUTF1 within the nucleus, as well as exclusion from the nucleoli, which are both characteristics of wild-type GFP-hUTF1, was observed with all SNPs (Kooistra et al., 2009). In addition, association of GFP-hUTF1 SNPs with chromosomes during mitosis was also observed (insets in Fig. 2).
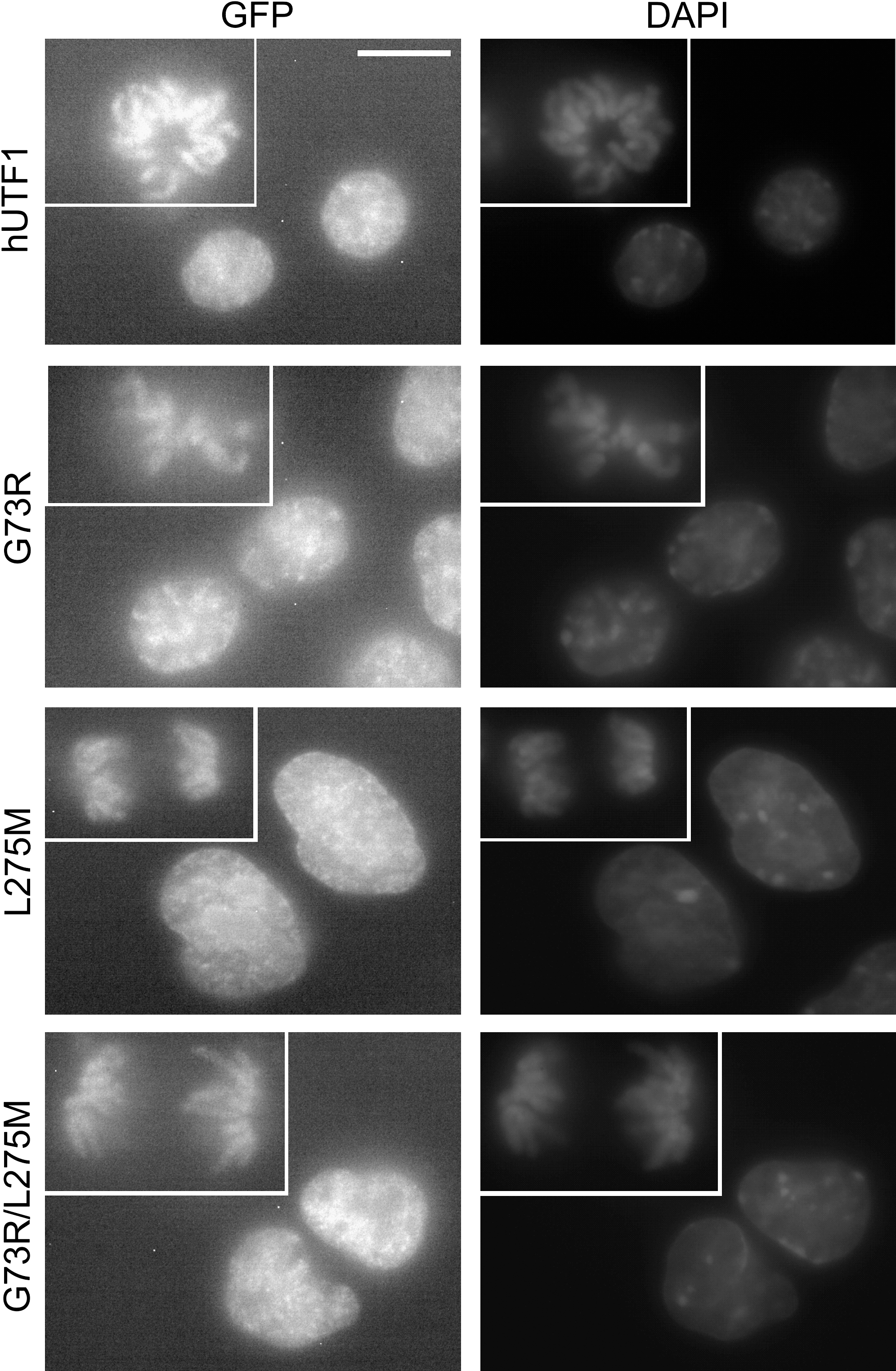
Localization of eGFP-human UTF1 (hUTF1) mutants. Fluorescence microscopy of P19CL6 cells stably expressing eGFP fused to hUTF1 or the hUTF1 SNPs indicated and counterstained with DAPI. Mitotic nuclei are shown in the insets. The scale bar is 15 μM.
Transcriptional repression by hUTF1 SNPs
To determine if transcriptional repression by hUTF1 was affected by the SNPs, luciferase reporter assays were performed. HepG2 cells were transiently transfected with fusions of the DNA binding domain of the yeast transcriptional activator protein GAL4 with hUTF1 and mutants in combination with an upstream activating sequence-luciferase reporter construct and a LacZ reporter as an internal control to correct for differences in transfection efficiency. Wild-type hUTF1 repressed reporter gene activity approximately sevenfold but the SNPs G73R, L275M, and G73R/L275M also repressed transcription to a similar degree. These data indicate that these mutations, either alone or together, do not perturb transcriptional repression of reporter gene activity by hUTF1 (Fig. 3).
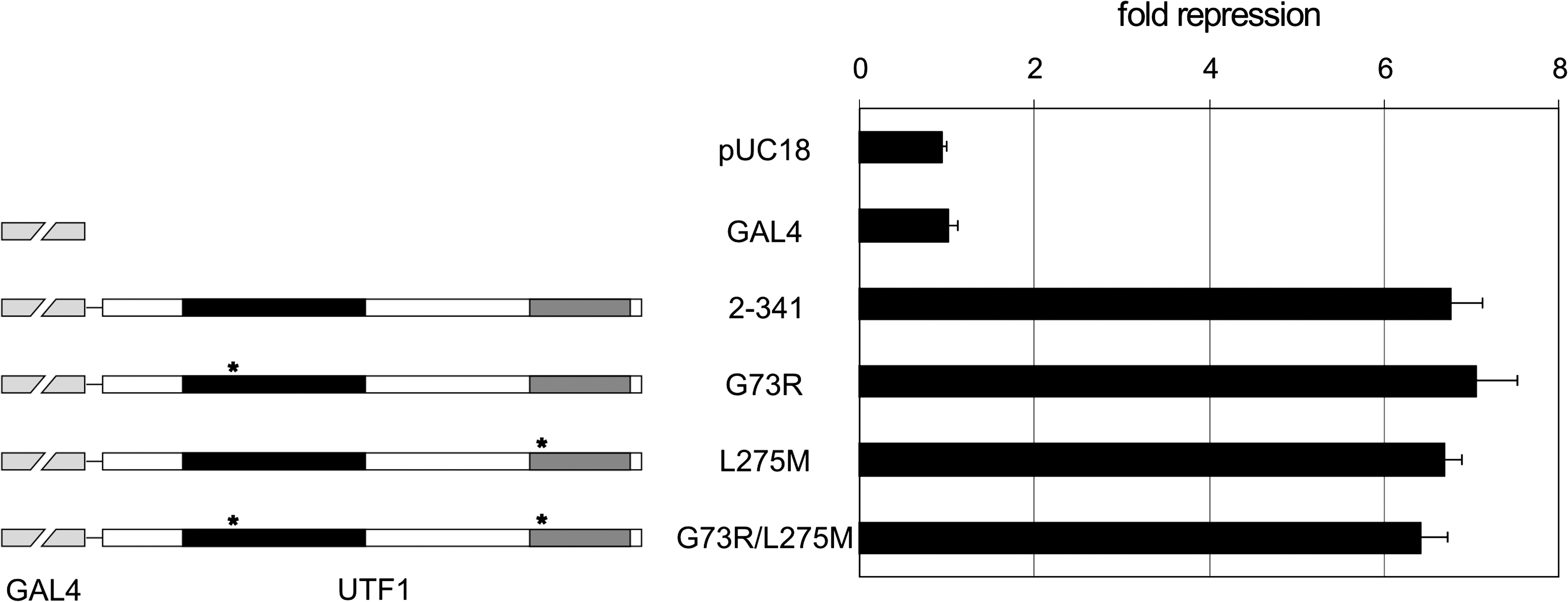
hUTF1 and hUTF1 SNPs repress transcription. A schematic representation of the GAL4-hUTF1 constructs used in reporter assays (left) and transcriptional repressor activity of different GFP-hUTF1 mutants (right). The CD1 and CD2 domains are indicated to scale by black and dark gray boxes, respectively. The location of point mutations G73R in CD1 and L275M in CD2 are indicated by asterisks. Different GAL4-hUTF1 mutants, a constitutively active TK-luciferase reporter containing five GAL4-binding sites (upstream activating sequence-TK-Luc), and a LacZ reporter as internal standard (pDM2-LacZ) were cotransfected into HepG2 cells. Transfections were repeated three times with independent DNA preparations, and in each experiment, all constructs were transfected in triplicate. A typical transfection is depicted as mean fold repression with standard deviations, with respect to GAL4 alone.
Subnuclear fractionation of GFP-hUTF1 SNPs
Recently, we reported that hUTF1, like mUTF1, is tightly associated with chromatin, with dynamic properties similar to core histones: an immobilization time in the order of minutes and a large immobilized fraction (>90%) (Kooistra et al., 2009). To assess whether the hUTF1-SNPs affected chromatin binding, resulting in an altered distribution over subnuclear fractions, P19CL6 cell lines stably expressing G73R, L275M, and G73R/L275M were analyzed. Stable cell lines that express relatively low levels of the transgene were used, to avoid aberrant fractionations. As we reported earlier, eGFP-hUTF1 and the endogenous mUTF1 protein localized exclusively to the strongly DNA-associated protein fraction, which also contained core histone H2A (Kooistra et al., 2009). SNP G73R and L275M also exclusively localized to the ammonium sulphate fraction. However, a hUTF1 mutant containing both SNPs (G73R/L275M) not only localized to the strongly DNA-associated fraction but was also detected in the free diffusing fraction as well, indicating the presence of an increased proportion of mobile hUTF1 molecules (Fig. 4A).
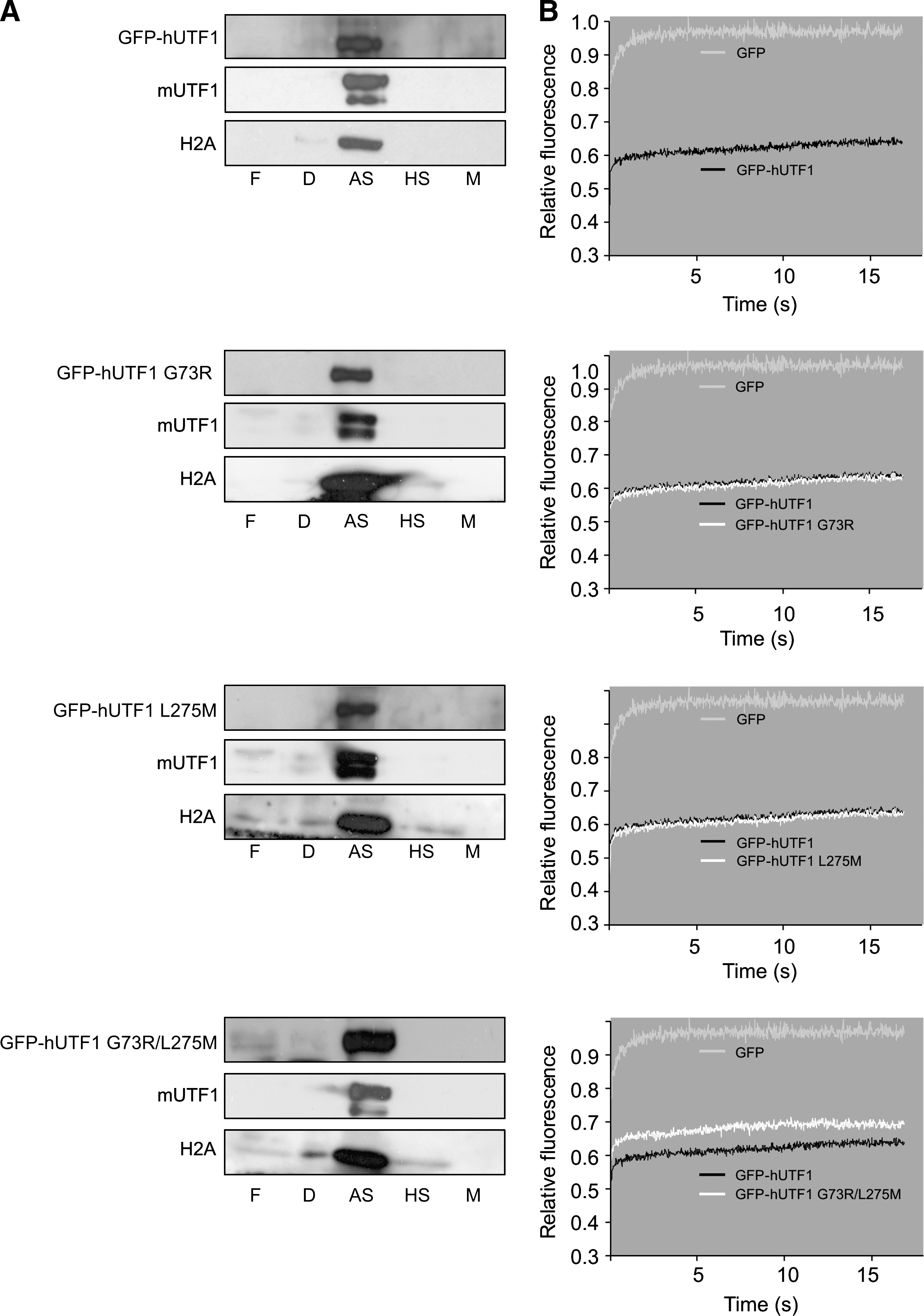
Analysis of subcellular localization and mobility of eGFP-hUTF1 SNPs. Subnuclear fractionations (
Mobility of GFP-hUTF1 SNPs in living cells
To confirm the stable chromatin association of hUTF1 containing single SNPs and the mobility of a fraction of the G73R/L275M mutant, we analyzed the dynamic properties of these proteins in living cells using FRAP. In P19CL6 cells that stably express low levels of GFP-hUTF1 fusions, these GFP fusion molecules were bleached in a small strip spanning the nucleus, and subsequent fluorescent recovery in the strip was measured (van den Boom et al., 2004). The mean fluorescence intensity in the strip of several cells was plotted against time relative to the prebleach level. GFP-expressing cells showed a fast recovery of fluorescence in the strip (Fig. 4B, light gray wavy line), indicating a highly mobile protein. Fluorescence in the strip did not recover to prebleach levels as a result of the permanent bleaching of a fraction of the molecules. In contrast to GFP, GFP-hUTF1 (Fig. 4B, black wavy line) showed only little recovery after bleaching, indicating that the vast majority is long-term immobilized, at least for the duration of the FRAP experiments. G73R and L275M (Fig. 4B, white wavy lines) SNPs showed nearly identical curves to the wild-type GFP-hUTF1 protein, indicating that the GFP-hUTF1 protein carrying these SNPs is still immobilized by strong association to chromatin. However, the double mutant G73R/L275M did not reach full recovery but showed a 7% increase in fluorescence recovery in the strip compared with GFP-hUTF1, indicating a reduced binding efficiency (Fig 4, white line). The rate of fluorescence recovery of G73R/L275M after the initial influx resembled that of GFP-hUTF1, implying that the residence time of G73R/L275M was not affected. Summarizing, the strip-FRAP and subnuclear fractionation data indicate that the double SNP G73R/L275M results in a partial loss of hUTF1 function because a fraction of the protein is not tightly associated with chromatin but free diffusing.
Discussion
Recently, we reported that both the mouse and human UTF1 proteins are strongly associated with chromatin, expressed in pluripotent cells, and able to repress transcription (van den Boom et al., 2007; Kooistra et al., 2009). Further, UTF1 is required for proper murine ES cell differentiation because ES cells depleted of UTF1 failed to properly initiate differentiation programs (van den Boom et al., 2007). The aim of this study was to determine if reported missense SNPs in the coding region of the human UTF1 gene have a deleterious effect on the function of hUTF1. Fluorescence microscopy indicated that all hUTF1 mutants were nuclear localized and repressed reporter gene activity. Both subnuclear fractionation and strip-FRAP showed that the combination of SNPs G73R and L275M resulted in an increased mobile fraction, and hence, less-functional hUTF1 protein.
Earlier we reported that the CD1 domain in human UTF1 is required for proper nuclear targeting of the protein. Deletion of CD1 as well as a W63G mutation resulted in cytoplasmic mislocalization of UTF1 (Kooistra et al., 2009). The SNP G73R is localized in the CD1 domain of UTF1, in close proximity to the W63 residue, and both W63 and G73 are localized in the first helix of the helix-turn-helix motif of the CD1 domain and completely conserved in UTF1 from different species. Further, we previously observed that the mutation of residues W63 E67 in mouse UTF1 resulted in an increased fraction of mobile protein, but with a residence time similar to that of a wild-type when bound to sites of affinity. As the G73 residue is very close to these residues, fully conserved across species, and in the first helix of the Myb/SANT domain, it was very unexpected that no effect of the G73R SNP on UTF1 functionality was observed in the assays used. However, it is possible that the G73R SNP results in a dysfunctional hUTF1 protein with altered target gene specificity. As a consequence, this mutation could affect human embryonic development, most likely at the blastocyst stage or during primordial germ cell differentiation.
SNP L275M is localized in the CD2 domain of UTF1, which contains a putative leucine zipper. Mutations of L293P and L300P resulted in a highly mobile protein, indicating that the leucine zipper is required for stable long-term chromatin interaction of hUTF1 (Kooistra et al., 2009). SNP L275M, however, had no effect on chromatin association in terms of residence time and mobility of the protein in FRAP assays. The evidence that this missense mutation does not have a strong detrimental effect on hUTF1 function could be due to the location of this SNP in the leucine zipper alpha helix chain. A leucine zipper is characterized by a leucine at every seventh position (Landschulz et al., 1988). The leucines at positions 293 and 300 are at the teeth of the zipper that stabilize its conformation. L275, however, is not a “teeth” leucine, and mutation of this residue might not strongly interfere with the structure of the leucine zipper. Alternatively, previous data indicated that the CD2 domain is required for long-term immobilization of UTF1 on chromatin, presumably through protein–protein interactions. It is possible that SNP L275M affects the affinity or specificity of hUTF1 for interaction partners and hence have a detrimental effect on early embryonic human development.
Although single mutants G73R and L275M seemingly did not alter the functionality of hUTF1, the presence of both SNPs clearly affected hUTF1 function. This suggests that, although the effects of each separate SNP are not detectable, when both are present they have a synergistic effect. A fraction of the hUTF1 proteins carrying both the G73R and L275M mutations was detected in the free diffusing fraction in subnuclear fractionations, indicating the presence of an increased proportion of mobile molecules. This observation was supported by the strip-FRAP experiments where an increased mobile fraction was detected with G73R/L275M GFP-hUTF1. When immobilized, the residence time of this mutant was very similar to wild-type hUTF1, indicating that these mutations probably affect the association to sites of affinity but not the duration of its interaction with these sites. Whether this increased mobile fraction is a consequence of altered target sequence specificity or altered interaction partners of this mutant hUTF1 remains to be determined.
We previously reported that deletion of hUTF1 CD2 resulted in a complete loss of repressor activity in reporter assays. Deletion of CD1 or mutation of the leucine zipper had no effect on hUTF1 repressor activity, indicating that the hUTF1 repressor domain is localized in its CD2 but does not require the leucine zipper (Kooistra et al., 2009). When G73R was introduced in hUTF1, as expected, no effect on the transcriptional repressor activity in the reporter assays was observed. SNP L275M, however, is located in CD2, but also no effect of this mutation of hUTF1 repressor activity was detected. The double SNP, G73R/L275M, also repressed reported constructs such as wild-type hUTF1, where an increased mobile fraction was detected in subnuclear fractionations and strip-FRAP assays. In the reporter assays, GAL4 is used as a DNA targeting domain that binds the upstream activating sequence. GAL4-hUTF1 fusions are thus not dependent on hUTF1 for DNA binding and thereby the GAL4 domain probably masks the effect of the double SNP on chromatin association by hUTF1. Moreover, the frequency of the SNP L275M is not known, so it is at present not possible to determine the frequency of the simultaneous presence of SNPs G73R and L275M.
Summarizing, these data show that the human UTF1 gene contains two missense SNPs that, when both present, affect the function of hUTF1. If that results in an altered target gene specificity, chromatin structure or differentiation defects in pluripotent cells remain to be elucidated.
Footnotes
Acknowledgment
This work was supported by the Groningen Biomolecular Sciences and Biotechnology Institute.
Disclosure Statement
No competing financial interests exist.