
Abstract
Evodiamine-induced apoptosis has been shown to have anticancer activity by eradication of some carcinoma cell lines. This study was designed to evaluate the effects of evodiamine on the viability of human gastric cancer SGC-7901 cells and to define the cell death pathway. Flow cytometry detection showed that 1.5 μM evodiamine significantly induced SGC-7901 cell apoptosis in a time-dependent manner. This apoptosis was partially inhibited by the pancaspase inhibitor carbobenzoxy-valyl-alanyl-aspartyl-[O-methyl]-fluoro-methylketone, which suggests that evodiamine-induced apoptosis in SGC-7901 cells is partially caspase independent. Further, the total content of sphingomyelin was decreased and expression of acid sphingomyelinase (aSMase) and neutral SMase genes in the SGC-7901cells was upregulated. Protein expression of aSMase, which was exposed to evodiamine, was shown to be increased by western blot analysis and could have been responsible for inducing caspase-independent apoptosis. Our results indicate that evodiamine stimulates upregulation of aSMase expression and hydrolysis of sphingomyelin into ceramide, which might be one of the mechanisms by which apoptosis occurs in SGC-7901 cells.
Introduction
However, the mechanisms that underlie the antiproliferative, antimetastatic, and apoptotic activities of evodiamine are incompletely understood. We postulated that this compound might mediate its effects by causing sphingomyelinases (SMases) to trigger the hydrolysis of sphingomyelin into ceramide, a potent signaling molecule that is critical for induction of apoptosis. The sphingomyelin pathway is initiated by hydrolysis of sphingomyelin to generate the second messenger ceramide through the action of SMases (Bose et al., 1995). Two SMases, acid SMase (aSMase) and neutral SMase (nSMase), play important roles in sphingomyelin hydrolysis (Matmati and Hannun, 2008) and are activated in response to many extracellular stimuli (Marchesini and Hannun, 2004). It has been suggested that ceramide modifies the stress response (Hannun and Luberto, 2000) and functions as a common lipid mediator in programmed cell death. A growing body of literature suggests that response to cellular stress is one function of sphingolipids, which indicates that ceramide-related pathway is a fertile area for the identification of therapeutic targets (Reynolds et al., 2004).
Gastric cancer is one of the most common causes of malignancy-related death worldwide. At present, gastric cancer patients have certain clinical responses to chemotherapy or radiation therapy, but they cannot tolerate it well (Macdonald, 2004); therefore, it has been a focus to explore drugs capable of preventing and treating gastric cancer. SGC-7901, a human moderately differentiated gastric adenocarcinoma cell line (Lin et al., 1984), was employed in our study. We examined the apoptosis-inducing potential of evodiamine in SGC-7901 cells and the evodiamine-induced apoptotic signaling pathway. Evodiamine was still able to induce slight SGC-7901 cell apoptosis after the caspase cascade was blocked by Z-VAD-FMK. This indicated that evodiamine-induced cell death was partly mediated by the caspase-dependent pathway and caspase-independent cell death. Further, we demonstrated that evodiamine could induce upregulation of aSMase expression to hydrolyze sphingomyelin into ceramide.
Materials and Methods
Reagents
Evodiamine was obtained from the Chinese National Institute for the Control of Pharmaceutical and Biological Products, reconstituted in dimethyl sulfoxide (DMSO) (Sigma–Aldrich) to a concentration of 30 mM, and stored at −20°C. Sphingomyelin and thiazolyl blue tetrazolium bromide (MTT) were from Sigma–Aldrich. Annexin V-FITC Apoptosis Detection Kit I was purchased from BD Pharmingen. Z-VAD-FMK was obtained from Promega. Moloney Murine Leukemia Virus (M-MLV) Reverse Transcriptase and Taq DNA Polymerase were purchased from Promega. aSMase antibody and nSMase antibody were purchased from Santa Cruz Biotechnology. PVDF membrane was obtained from Millipore.
Cell lines and cell culture
The human gastric cancer cell line SGC-7901 was obtained from the Shanghai Type Culture Collection of the Chinese Academy of Sciences. Cells were propagated in RPMI-1640 medium supplemented with 10% fetal bovine serum, 100 kU/L penicillin, and 100 mg/L streptomycin at 37°C in a humidified atmosphere of 95% air and 5% CO2 (Li et al., 2006). Anticoagulated blood was obtained from healthy adult volunteers. Human lymphocytes were separated by density centrifugation in cell separation solution (Chuanye) and suspended in RPMI-1640 medium.
MTT assay
Cell proliferation was assessed using the MTT assay method. The treatment of living cells with MTT produces a dark blue formazan product, whereas dead cells remain unstained after treatment. MTT (5 g/L) was dissolved in phosphate-buffered saline (PBS). Cells were seeded in 96-well microplates at a density of 20,000 cells/well and incubated for 12 h. The cells were treated for the indicated time with different concentrations of evodiamine. MTT diluted in fresh medium (final concentration: 0.5 g/L) was added to each well and incubated at 37°C for 3 h. After this, the MTT loading medium was replaced with 10 μL DMSO, and the absorbance at 550 nm was measured for each well using an ELX800 absorbance microplate reader from BioTek Instruments. Cell numbers were determined from a standard plot of known cell numbers versus the corresponding absorbance density.
Apoptosis assays
The apoptotic cells were counted by flow cytometry after double staining with propidium iodide (PI) and fluorescein isothiocyanate (FITC)-conjugated annexin V (BD). The cells were washed twice in cold PBS and resuspended in annexin-V–binding buffer at a concentration of 1 × 106 per mL. This suspension (100 μL) was stained with 5 μL annexin V–FITC and 5 μL PI. The cells were gently vortexed and incubated for 15 min at room temperature in the dark. Then, 400 μL of 1 binding buffer was added to each tube. The apoptosis rate was detected using a cytofluorimeter and analyzed by FACScan and CellQUEST software (Becton Dickinson). Cells that were viable and not undergoing apoptosis were annexinV–FITC and PI negative, and cells undergoing apoptosis were annexin V–FITC positive and PI negative.
Real time reverse transcriptase–polymerase chain reaction
The isolation of RNA was carried out using the TRIzol reagent from Invitrogen. Synthesis of cDNA was performed using 50 ng RNA with M-MLV reverse transcriptase. Real-time reverse transcriptase polymerase chain reaction (RT–PCR) was performed to quantify the changes in the gene expression of aSMase, nSMase, and β-actin. Twenty microliters of reaction mixture contained 10 pM forward and reverse primers (Takara), 9 μL of 1 × SYBR Green reaction mixture (TianGEN), and 2 μL template. SYBR Green fluorescence was measured with the ABI 7500 fast (Applied Biosystems). aSMase was amplified using a set of primers (forward primer: 5′-CCGGCCCTTTTGATATGGTGT-3′; reverse primer: 5′-CAGCG GGAGGAGTGGTTGC-3′) to generate an amplicon of 220 bp; nSMase (forward primer: 5′-TGCTGCCTGCTGAAGGAGTGG-3′; reverse primer: 5′-CCTGTGAGGGTCAAAGCCTGTAGT-3′) of 234 bp; and β-actin (forward primer: 5′-AAGGTGACAGCAGTCGGTTGGA-3′; reverse primer: 5′-GAGAAGTGGGGTGGCTTTTAGGA-3′) of 156 bp. The PCR conditions were as follows: initial denaturation at 94°C for 2 min, 35 cycles with denaturation at 94°C for 30 s, annealing at 59°C for 40 s, and elongation at 68°C for 40 s.
Western blot
Expression of aSMase and nSMase was investigated by western blotting (Claus et al., 1996; Opreanu et al., 2010). Protein samples were prepared by boiling lysates in a reducing sodium dodecyl sulfate sample buffer. Proteins were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis, blotted on a PVDF membrane, blocked with 5% nonfat dry milk in TBS-T buffer (10 mM Tris, 150 mM NaCl, and 0.2% Tween 20 [pH 8.0]) overnight at 4°C, and subsequently probed with the appropriate primary antibody. Immunoreactive bands were visualized using a chemiluminescence kit (ECL-Plus; GE Healthcare).
High-performance liquid chromatography analysis
The cell samples (approximately 4 × 106 cells) were collected and washed twice with PBS, and the total lipids were extracted according to standard procedures (Tepper and Van Blitterswijk, 2000). The total amount of sphingomyelin in the extracted lipids was determined by Agilent 1100 high-performance liquid chromatography (HPLC) using methanol:acetonitrile (70:30) to resolve the lipids on a C18 silica column (the lipids were resuspended in N,N-dimethylformamide:methanol (1:1) before injection onto the column) (Modrak, 2005).
Statistical analysis
All results were confirmed in at least three separate experiments. The data are expressed as the mean ± standard error. Data of the representatives were analyzed for statistical significance by the Student's t-test. A p-value of <0.05 was considered significant.
Results
Effects of evodiamine on cell growth
The MTT assay was used to quantitatively determine the effect of evodiamine on the proliferation of SGC-7901 cells and human lymphocytes. Cells treated with 0.1% DMSO were used as the control. As shown in Figure 1A, evodiamine caused SGC-7901 cell death at concentrations ranging from 0.1 to 15 μM during 36 h of culture and significantly decreased proliferation of SGC-7901 cells in a dose- and time-dependent manner. However, evodiamine did not cause significant lymphocyte death with the same treatment (Fig. 1B).
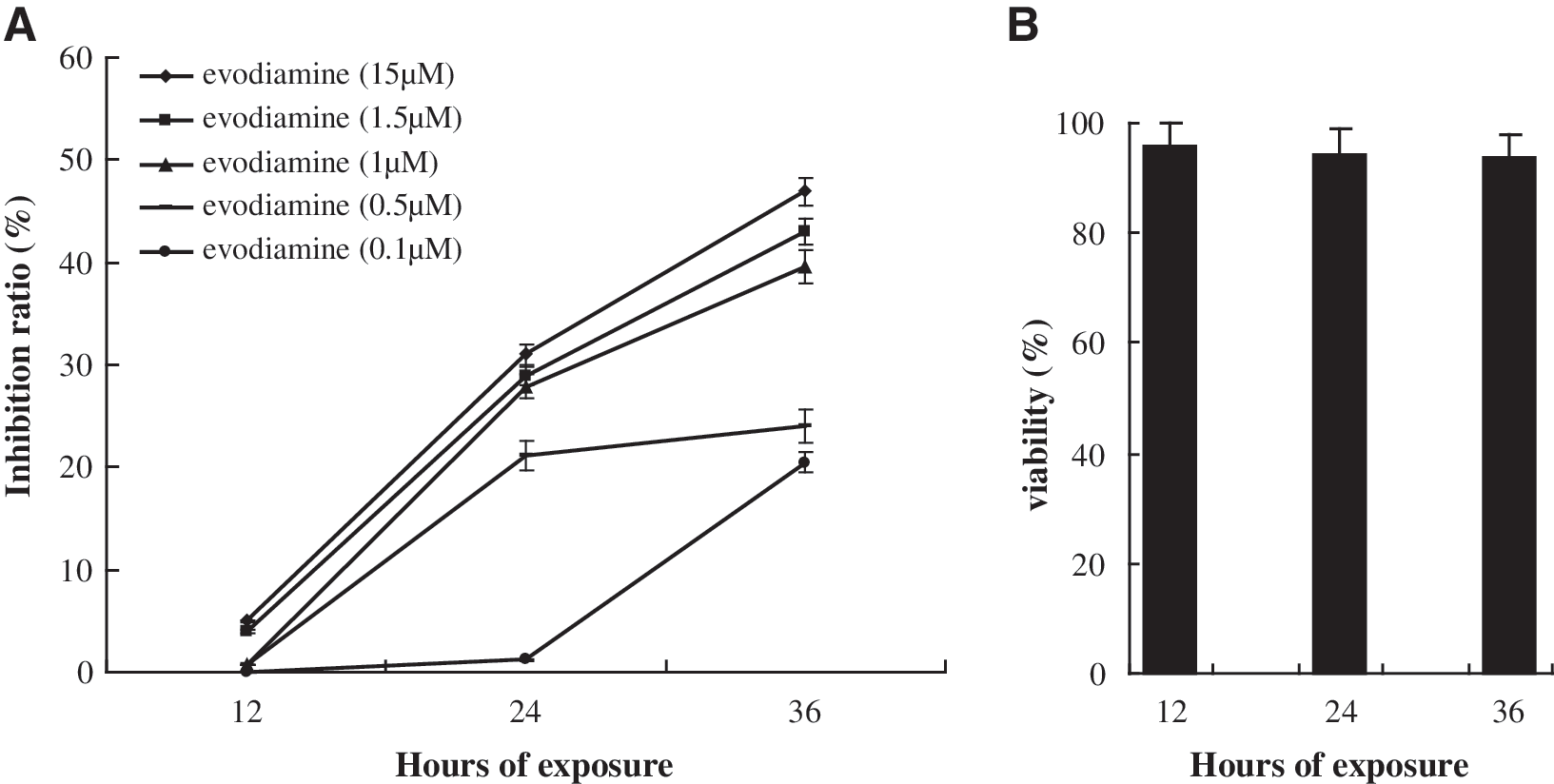
Effects of evodiamine on the proliferation of SGC-7901 cells and human lymphocytes by thiazolyl blue tetrazolium bromide assay.
Evodiamine-induced apoptosis of cells
Exposure of SGC-7901 cells to evodiamine and evodiamine plus Z-VAD-FMK was measured by flow cytometry using double staining with PI and FITC-conjugated annexin V. Evodiamine induced apoptosis of SGC-7901 cells in a time-dependent manner. This evodiamine-induced cell death was only partially prevented by the pancaspase inhibitor Z-VAD-FMK (Fig. 2). These results suggested that evodiamine-induced cell death was partly mediated by a caspase-dependent pathway and caspase-independent cell death in the presence of Z-VAD-FMK.
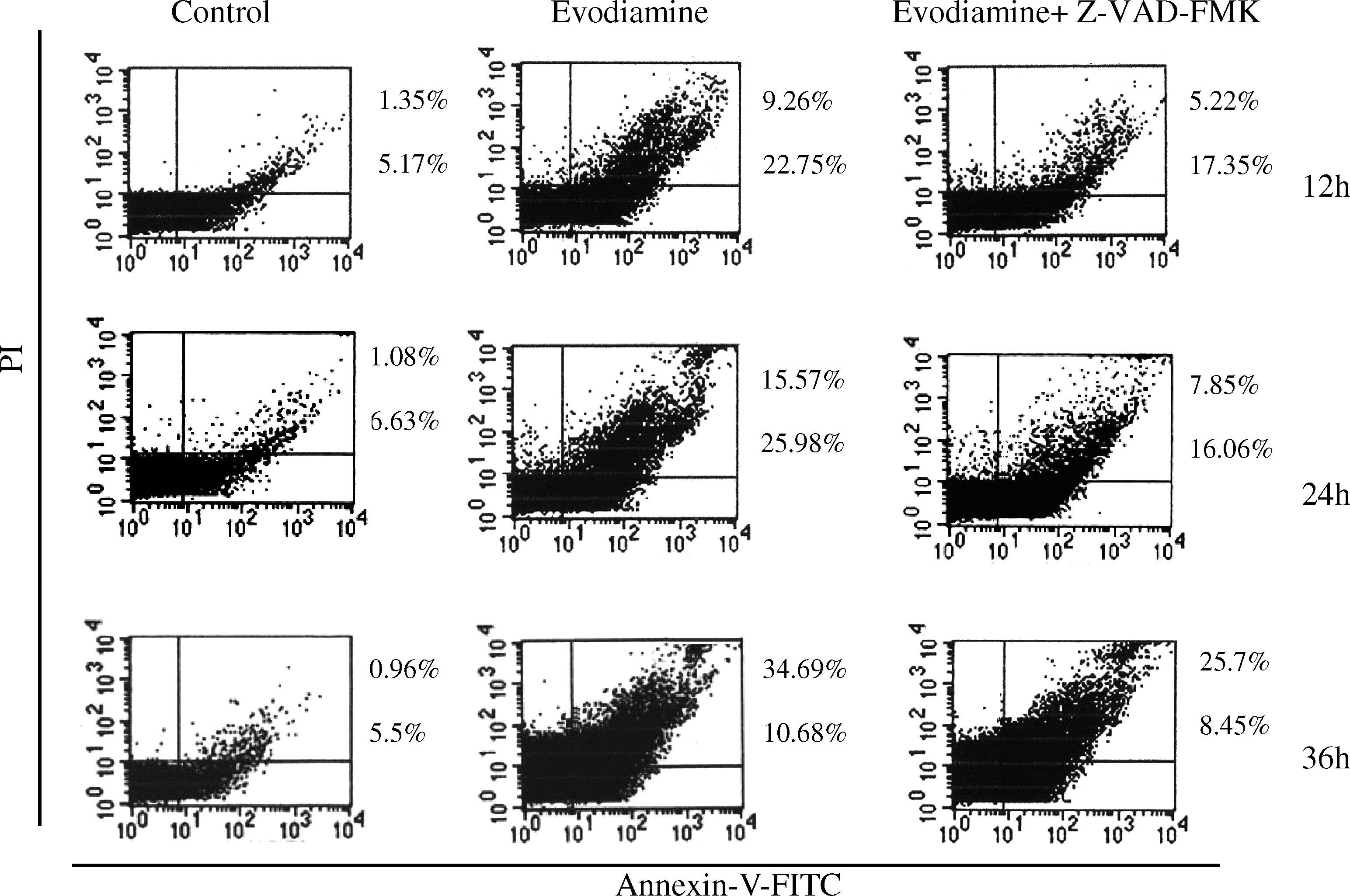
Caspase-dependent and -independent apoptosis of SGC-7901 cells. The SGC-7901 cells were treated with 1.5 μM evodiamine alone or incubated with 20 μM Z-VAD-FMK before challenge with 1.5 μM evodiamine for 12, 24, and 36 h and then harvested and stained with FITC-conjugated annexin V and PI. Cell death was determined by flow cytometry. Bottom right quadrant: annexin V+/PI−, early apoptotic cells; top right quadrant: annexin V+/PI+, late apoptotic cells. Values correspond to the percentage of cells in those quadrants. Two additional studies yielded equivalent results. Z-VAD-FMK, carbobenzoxy-valyl-alanyl-aspartyl-[O-methyl]-fluoro-methylketone; FITC, fluorescein isothiocyanate; PI, propidium iodide.
Role of aSMase upon exposure to evodiamine
To determine whether aSMase and nSMase were involved in evodiamine-induced apoptosis of SGC-7901 cells, we tested the levels of mRNA by real-time RT-PCR and protein levels by western blotting. The results of real-time RT-PCR show upregulation of aSMase and nSMase gene expression. aSMase gene expression was clearly higher than that of nSMase (Fig. 3A). Western blotting verified the protein expression of aSMase and nSMase; the results also showed that aSMase protein levels were higher than those of nSMase (Fig. 3B). These results indicated that evodiamine triggered the SGC-7901 cells to stimulate upregulation of aSMase.
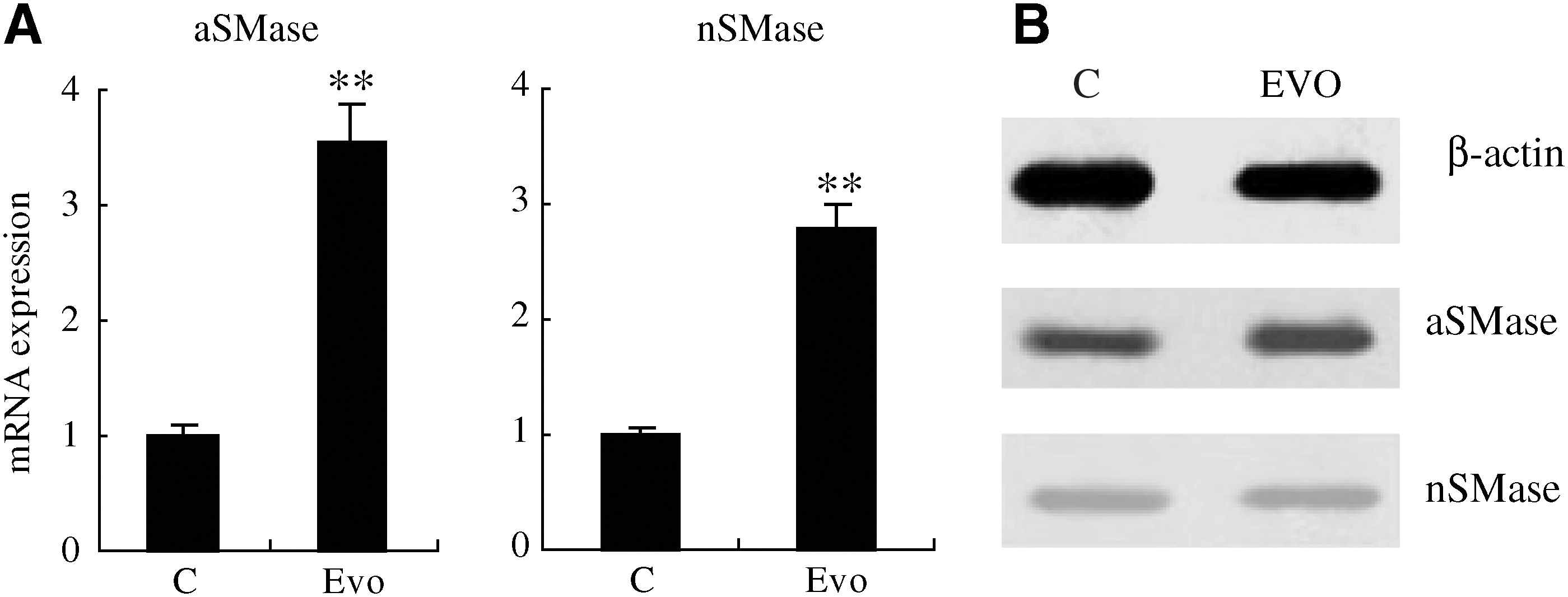
Expression levels of aSMase and nSMase genes in SGC-7901 cells.
Changes of sphingomyelin contents
Upregulation of aSMase in SGC-7901 cells was affected by evodiamine; therefore, we investigated the content changes of sphingomyelin, the substrate of aSMase, to determine whether aSMase was activated in these cells. As shown in Figure 4, sphingomyelin was reduced in cells exposed to evodiamine compared with the controls, and D609, an aSMase inhibitor, was able to inhibit hydrolysis of sphingomyelin. The data indicate that aSMase played a crucial role in the hydrolysis of sphingomyelin to ceramide in SGC-7901 cells exposed to evodiamine.
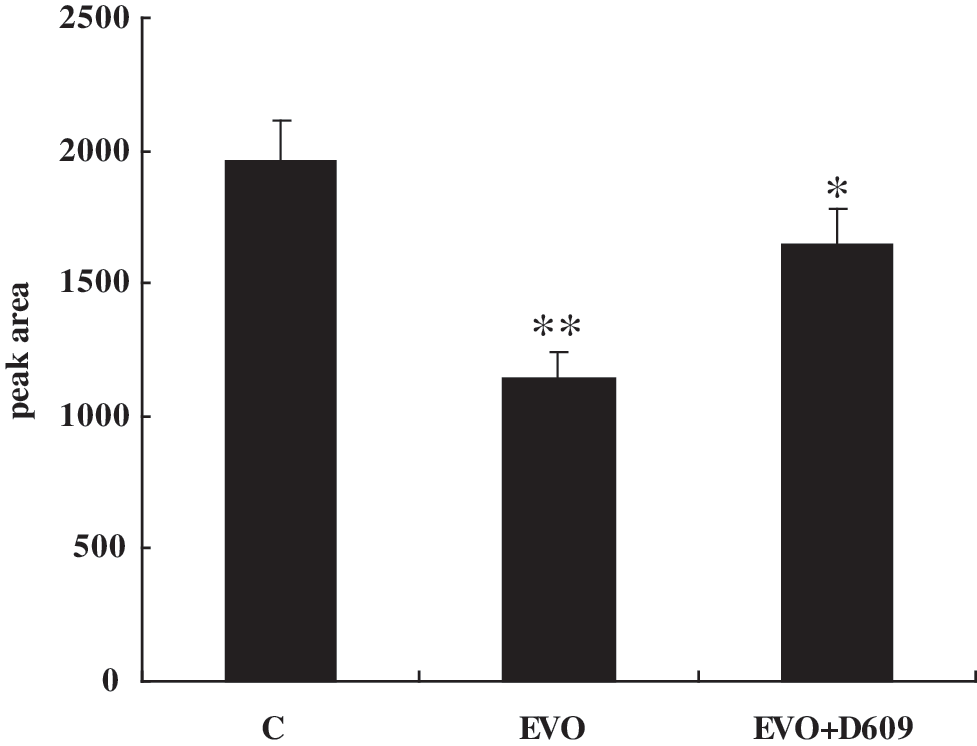
Changes of sphingomyelin in SGC-7901 cells exposed to evodiamine. Sphingolipids were extracted from control (C), evodiamine (EVO), and evodiamine plus D609 (EVO + D609) stimulated cells. Aliquots of the lipid extracts used for determining total phospholipid and sphingomyelin contents were determined by high-performance liquid chromatography. The SGC-7901 cells treated with 1.5 μM evodiamine showed a decrease in sphingomyelin, whereas the SGC-7901 cells incubated with 100 μM D609 before the challenge with 1.5 μM evodiamine did not show an obvious decrease in sphingomyelin. The results indicated that sphyingomyelin was hydrolyzed by aSMase during exposure of SGC-7901 cells to evodiamine. *p < 0.05; **p < 0.01 versus control.
Discussion
It has been recently reported that evodiamine can induce apoptosis of tumor cells through various mechanisms, including caspase-dependent and -independent pathways (Fei et al., 2003; Liao et al., 2005; Lee et al., 2006a). The sphingomyelin pathway is a ubiquitous signaling system and plays the role of a second messenger, mainly through generation of ceramide during hydrolysis of sphingomyelin by SMase (Dbaibo et al., 2001). It also regulates signaling processes such as apoptosis, proliferation, differentiation, and growth delay, of which the regulation of apoptosis is most important (Carpinteiro et al., 2008; Kitatani et al., 2008). In previous studies on the induction of apoptosis of tumor cells by evodiamine, there has been no report on the metabolic pathway of sphingomyelin. This study aimed to investigate whether evodiamine induced the apoptosis of gastric cancer SGC-7901 cells through the sphingomyelin pathway and its possible mechanisms.
Through our preliminary experiments, it was found that evodiamine significantly inhibited the proliferation of SGC-7901 cells and the effects were dose and time dependent, but evodiamine had little antiproliferative effect on normal human lymphocytes. It was shown by combination of annexin V with flow cytometry that evodiamine could effectively induce apoptosis of SGC-7901 cells in a time-dependent manner. When evodiamine was added after the addition of the caspase inhibitor, Z-VAD-FMK, apoptosis of SGC-7901 cells was weakened but not completely blocked. It was shown that, in addition to the caspase apoptotic pathway, other apoptosis pathways were also involved in the apoptosis of SGC-7901 cells induced by evodiamine.
Sphingomyelin metabolism also induces apoptosis. Sphingomyelin exerts its apoptotic function downstream, mainly through the generation of ceramide by hydrolysis of aSMase and nSMase (Bose et al., 1995). Stimulated by many factors, SMase may have an increased activity and thus the level of sphingomyelin will be reduced. To confirm the effects of treatment of SGC-7901 cells with evodiamine on hydrolysis of sphingomyelin, expression of aSMase and nSMase genes was detected using real-time RT-PCR. We found that the expression of both genes was upregulated compared with that of the control group. It was confirmed through western blotting that protein levels of nSMase did not clearly change, and aSMase was still significantly expressed, which suggested that the reduction in sphingomyelin content induced by evodiamine was mainly due to the upregulated expression of aSMase. Sphingomyelin content in SGC-7901 cells was determined by HPLC. The content of sphingomyelin in the cells treated with evodiamine was significantly lower than that in the control group, which indicated that the decomposition of sphingomyelin by aSMase in the induced cells was increased because of the action of evodiamine.
Further, SGC-7901 cells were treated with evodiamine after addition of the aSMase inhibitor D609 (Cogolludo et al., 2009). It was determined by HPLC that the sphingomyelin content was not significantly reduced, which suggested that hydrolysis of sphingomyelin by aSMase was significantly decreased after inhibition of the enzyme. These results convincingly showed that the apoptosis of SGC-7901 cells induced by evodiamine was due to upregulated expression of aSMase after stimulation by evodiamine, which promoted hydrolysis of sphingomyelin and led to generation of ceramide, thereby inducing apoptosis of the cancer cells.
In conclusion, our data provide experimental evidence that evodiamine is a potent inhibitor of human SGC-7901 gastric cancer cell proliferation and that it induces cell death through caspase-dependent and -independent apoptosis. Evodiamine stimulates upregulation of aSMase expression, resulting in sphingomyelin being hydrolyzed into ceramide, which might be one of the mechanisms that enables evodiamine to trigger SGC-7901 cell apoptosis. Thus, evodiamine could be developed as a potential chemotherapeutic or chemopreventive agent against human gastric carcinoma.
Footnotes
Acknowledgment
This study was supported by a grant (No. 2009-60) from the Administration of Traditional Chinese Medicine, Guizhou Province, China.
Disclosure Statement
No competing financial interests exist.