
Abstract
Age-related macular degeneration (AMD) is a disease with multifactorial etiology characterized by irreversible loss of central visual acuity. The discovery of susceptive single-nucleotide polymorphisms (SNPs) has progressed our understanding of AMD. Complement factor H (CFH) gene Y402H polymorphism and high-temperature requirement A-1 (HTRA1) LOC387715 gene A69S polymorphisms are the most important SNPs reported in the literature. Determination of genetic risk factors and genotype–phenotype relationship in AMD may result in rapid and cost-effective therapeutic applications for young and old population. In this study, we hypothesized a potential association between CFH gene Y402H and HTRA1 LOC387715 gene A69S polymorphism in Turkish AMD patients. In blood samples from a total of 252 individuals, 147 clinically diagnosed as AMD and the others control, polymorphic sites in CFH, Y402H (Tsp509I T/C), and HTRA1, LOC387715 A69S (FnuHI G/T), were determined by polymerase chain reaction–restriction fragment length polymorphism analysis. There was significant difference between CFH genotypes in the AMD group, TT 21.8%, TC 48.3%, and CC 29.9%, and in the control subjects, TT 45% (p=0.003), TC 41% (p=0.0001), and CC 14% (p=0.0001). Further, the A69S polymorphism of LOC387715 was investigated and found to be significantly associated with AMD. LOC387715 genotypes in the AMD group were GG 30.6%, GT 38.1%, and TT 31.3% and in the control subjects were GG 59% (p=0.027), GT 39% (p=0.0001), and TT 2% (p=0.0001), respectively. We also found that Y402H C and A69S T allele were associated with AMD. This is the first study showing that Y402H and LOC387715 are associated with AMD in Turkish population.
Introduction
AMD is a multigenic, multifactorial disease and its pathogenesis remains poorly understood. However, family-based genome-wide and candidate region linkage studies on the two most promising linkage regions occurring on chromosome 1q31 and 10q26 have contributed to the understanding of its pathogenesis (Fisher et al., 2005). A large number of single-nucleotide polymorphisms (SNPs) including complement factor H (CFH) gene Y402H and high-temperature requirement A-1 (HTRA1) gene LOC387715 were found to be associated with progressed AMD, geographic atrophy, and neovascular disease (Fisher et al., 2005). Y402H (rs1061170) polymorphism results in a T→C change at nucleotide position 1277 on exon 9 of the CFH gene (MIM 134370), changing a tyrosine amino acid at position 402 to a histidine (Y402H). This change may increase risk for AMD (odds ratio (OR) 2.1 to 7.4) (Klein et al., 2005). Hypothetical LOC387715 gene rs10490924 or A69S polymorphism located at chromosomal region 10q26, was identified as the second major AMD susceptibility locus (Scholl et al., 2007). HTRA1, expressed in both the human and mouse retina, is a heat-shock serine protease and is upregulated by cellular stress (Dewan et al., 2006). HTRA1, which resides at chromosomal region 10q26, was implicated as the candidate region for AMD independent of CFH. LOC387715/age-related maculopathy susceptibility-2 (ARMS2) gene A69S polymorphism (rs10490924) is one of the most important reported SNPs and is located in the coding region of the putative genomic locus LOC387715/ARMS2 and 6.6 kb upstream of HTRA1 gene (Yoshida et al., 2007; Yoshimura 2010).
Environmental factors such as age, smoking, light exposure, and body mass index play an important role in the AMD pathogenesis during gene–gene interactions (Katta et al., 2009). AMD has been classified based on specific clinical features as early AMD and late AMD, which were subclassified into dry and wet categories. Early AMD is characterized by presence of soft, indistinct or reticular drusen or any drusen with pigmentary changes in the macular region, whereas late AMD is characterized by geographic atrophy or choroidal neovascularization. Dry AMD, which is the severe form of the disease, accounted for 80%–90% of the cases, and wet AMD accounted for the rest (Hageman et al., 2001).
Genetic predisposition associated with AMD was based on significant number of risk-modifying SNPs, which were identified among protein cascade genes in AMD. The aim of the present study was to investigate the frequency of the Y402H and LOC387715 A69S polymorphisms in Turkish population as a risk factor for AMD-associated vision loss.
Materials and Methods
Patients
This case–control study consisted of 147 patients (82 males [56%] and 65 females [44%]; mean age: 70.80±8.6) clinically diagnosed as having AMD and 105 (50 males [48%] and 55 females [52%]; mean age: 64.63±8.5) ophthalmologically evaluated non-AMD patients as control group. The patients were from the West Anatolian Turkish population, were not related, were not from specific subpopulations or ethnicities, and have similar lifestyles. The study was conducted according to the Helsinki declaration and was approved by the Ethics Committee of the Afyon Kocatepe University. All cases and controls were enrolled in this study under informed consent. Selection criteria have been previously described and were modified from Age-Related Eye Disease Study grading system and overall grade was based on the most severely affected eye (Postel et al., 2005). Patients were examined by trained ophthalmologists by slit-lamp examination, biomicroscopy with a handheld 90-diopter lens or fundus contact lens, and 20-diopter indirect ophthalmoscopy of the peripheral retina. Also, 35 mm color fundus photographs as well as stereophotographs of the disc and macula were obtained for all of the patients according to the study by Shuler et al. (2007). Individuals with disease of grade 3 or higher in at least one eye were considered the AMD patient group. Data collection for each individual was standardized by completing questionnaires about medical and familial histories.
Genotyping
Genomic DNA was isolated from whole blood samples using a commercially available DNA extraction kit (Omega, Omega Bio Tek) according to the manufacturer's instructions. Polymorphic sites in CFH Y402H (Tsp509I T/C, rs1061170) and HTRA1 LOC387715 A69S (FnuHI G/T, rs10490924) were determined by polymerase chain reaction–restriction fragment length polymorphism (PCR–RFLP) analysis. The following primer sequences were used for the amplification of the SNP of 1277 T>C (402Y>H) located in exon 9 of the CFH gene producing a PCR product of 241 bp: forward 5′ AGTGAGATGGCAGCTGGCTT 3′ and reverse 5′ GGTAAGGCCTGATCATCTGCA 3′. We set the PCR cycling conditions for the gene as described by Lau et al. (2006). Three genotypes, namely 1277 CC, 1277 TC, and 1277 TT, were analyzed by PCR-RFLP. The amplified DNA fragments were digested by FastDigest Tsp509I (MBI Fermentas) restriction enzyme at 65°C for 5 min. The presence of T at nucleotide position 1277 led to digestion of the 241 bp PCR product into DNA fragments of 60 and 181 bp.
The A69S variant of the LOC387715/HTRA1 gene locus was identified by the primer pair 5′ TCATTGTTATGGTCCTTAGGAAA 3′ (forward) and 5′ TTAGAAAGACATGAACATGCTAGG 3′ (reverse) and the PCR cycling conditions were as described by Pulido et al. (2007). Three genotypes, namely GG, GT, and TT, were analyzed by PCR-RFLP. The 394 bp fragment generated by PCR was digested with the restriction endonuclease FnuHI (New England Biolabs) at 37°C for 4 h. The A (wild type) allele gave bands of 199, 71 (2), and 53 bp. The S (risk) allele eliminated an FnuHI site and gave bands of 270, 71, and 53 bp. The PCR products and restriction fragments were separated in 2% agarose gel containing 0.5% ethidium bromide by electrophoresis and visualized by UVP BioDoc-It gel image system (UVP). LOC387715 homozygous TT genotype was confirmed by DNA sequence analysis from the same amplified SNP region (Fig. 1).
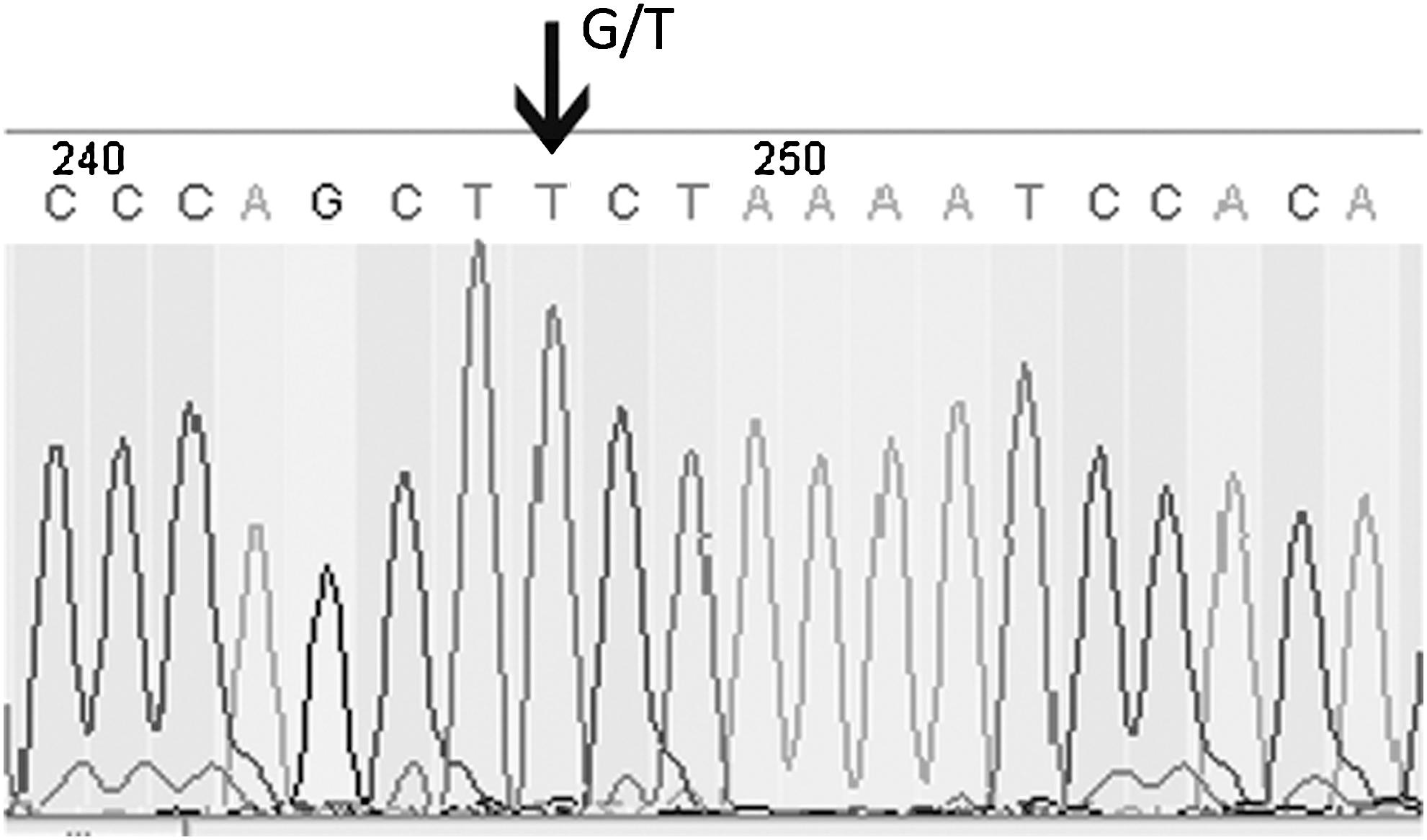
Analysis of the LOC387715 polymorphism. Forward sequence of LOC387715: CTTTATCACACTCCATGATCCCAGCT[G/T]CTAAAATCCACACTGAGCTCTGCTT. Arrow indicates the A69S G>T substitution. The nucleotide sequence GCT[G/T]CTA in the mutated allele is an FnuHI site.
Statistical analysis
Genotype distribution within the groups of the cases and controls was compared with values predicted by Hardy–Weinberg equilibrium using the chi-square test. Hardy–Weinberg equilibrium, based on allele frequencies, was verified with the following formula: p 2 + 2pq + q 2=1. The demographic characteristics such as age and gender were compared with those of controls by Student's t-test and Pearson's chi-square test. The difference of allelic and genotypic frequencies between the case and control groups were also estimated by the Pearson's chi-square test or Fisher's exact probability test. The relationships between genotypes and alleles and AMD risks were determined by obtaining the ORs and 95% confidence intervals (CI) through a logistic regression method. SPSS version 15.0 (SPSS, Inc.) was used to evaluate all statistical analyses. Chi-square values with a probability (p) value of <0.05 were considered statistically significant.
Results
A total of 147 cases of AMD whose ages ranged from 48 to 87 years (70.8±8.62) were evaluated. Patients were diagnosed according to the International Classification System and subdivided into three AMD degeneration grades as early AMD (grade 3, extensive intermediate drusen or any large, soft drusen [≥125 μm], including drusenoid retinal pigment epithelial detachment; n=19, 12.9%), geographic atrophy (grade 4, area of retinal pigment epithelium atrophy with sharp margins, usually visible choroidal vessels, ≥175 μm diameter; n=6, 4.1%), and exudative AMD (grade 5, including nondrusenoid pigment epithelium detachment, choroidal neovascular membrane, subretinal hemorrhage or fibrosis, or a photocoagulation scar consistent with treatment of AMD; n=122, 83%) (Postel et al., 2006). In the control population, ages were 64.63±8.5 (mean±standard deviation) and the mean age of controls was significantly lower than that of AMD cases by 6.07 years (p=0.001). To determine whether there was a population substructure in the AMD and control groups, we evaluated Hardy–Weinberg equilibrium for the CFH Y402H and LOC387715 A69S polymorphisms. We found that CFH Y402H genotype frequencies were in Hardy–Weinberg equilibrium in the AMD group (p=0.74) and also in the control group (p=0.49). The genotype frequencies of LOC387715 A69S were in Hardy–Weinberg equilibrium in the control group (p=0.87) and heavily skewed in all cases of the AMD group (p=0.004) (Tables 1 and 2). The genotype and allele frequency distributions of the Y402H and LOC387715 polymorphisms in patients and control subjects are shown in Table 3. There were significant differences between CFH genotypes in the AMD group (TT 21.8%, TC 48.3%, CC 29.9%) and control subjects (TT 42.9%; TC 42.9%, p=0.007; CC 14.3%, p=0.0001). The CFH 1277 C risk allele was significantly more frequent in the AMD group as 54.1% (p=0.0001) than in controls as 35.7% and the OR was 2.12 (95% CI: 1.47–3.05). The CC genotype was 29.9% in the AMD group and 14.3% in the control group (p=0.0001). The results confirmed the significant correlation (p<0.001) between the CC genotype and AMD (OR=4.12; 95% CI: 1.96–8.65). Additionally, the TC genotype conferred an increased risk (OR=2.22; 95% CI: 1.23–4.00) and significant association (p=0.007).
HWE, Hardy–Weinberg equilibrium.
Compared with TT versus TC, CC, and TC+CC.
Compared with GG versus GT, TT, and GT+TT.
Reference genotype/allele.
OR, odds ratio; CI, confidence interval.
Second, we investigated the polymorphism of A69S at LOC387715. rs10490924 SNP was also found to be significantly correlated with AMD. There were significant differences between LOC387715 genotypes in the AMD group (GG 30.6%, GT 38.1%, TT 31.3%) and the control group (GG 56.2%; GT 37.1%, p=0.027; TT 6.7%, p=0.0001). The frequencies of G and T alleles were 49.7% and 50.3%, respectively, in the AMD group, whereas in the control, their frequencies were 74.8% and 25.2%, respectively. The A69S thymine risk allele was significantly increased in the AMD group as 50.3% (p=0.0001) than in controls (25.7%) and the OR was 3.00 (95% CI: 2.04–4.42). The TT genotype was 31.3% in the AMD group and 6.7% in the control group (p=0.0001). The results confirmed the highly significant correlation (p<0.001) between the TT genotype and AMD (OR=8.61; 95% CI: 3.55 to 20.87). The GT genotype conferred a slightly increased risk (OR=1.88; 95% CI: 1.07 to 3.30) and less significant correlation (p=0.027). Our results showed that AMD cases were 2.12-fold to carry at least one copy of the C risk allele (p=0.0001), resulting in H402 compared with controls. At the same time, AMD cases were threefold to carry one copy of the T risk allele (p=0.0001), resulting in serine compared with controls.
For both SNPs, a total of 147 AMD cases and 105 controls were evaluated. Among 105 controls, 1 (0.95%) sample was homozygous for the risk alleles at both SNP loci. However, there were 15 (10.2%) double homozygous in the AMD group. Also, there were 9 (6.12%) double homozygous wild genotype in the AMD group. The highest percentage for AMD cases was 20.4% for the group who carried at least 1 risk allele (CFH +/− LOC +/−) at both loci. The lowest frequency for AMD cases was 6.12% for the group who lacked both of the variant genotype (CFH −/− LOC −/−). For the control group, the highest frequency was for the group that lacked both of the variant genotypes (CFH −/− LOC −/−) and the lowest frequency was for the group that was homozygous for both AMD risk variants (CFH +/+ LOC +/+; Table 4). If an individual had both homozygous risk genotypes CC (Y402H) and TT (LOC387715), compared with the nonrisk genotype TTGG, joint analysis of ORs for the two variants showed that the risk of AMD was 48-fold (p=0.0001), with all the other joint ORs ranging from 2.5 to 19 (Table 4).
Eight genotype combinations are compared with the nonrisk genotype combination TTGG. The risk alleles are C (Y402H) and T (LOC387715). All the patients with AMD (n=147) are compared with the controls (n=105).
Ref., reference; AMD, age-related macular degeneration.
The statistical power was calculated using the QUANTO 1.2 program (
Discussion
Y402H (rs1061170) and LOC387715 (rs10490924) have been implicated to play a role in determining the genetic risk for AMD (Hageman et al., 2001; Shuler et al., 2007). We also found that 1277 T>C (402 Y>H) was associated with AMD in a Turkish population. The 1277 C allele was associated with a 2.12-fold increased risk for the AMD group. Participants who carried homozygous 1277 C allele were at 4.12-fold higher risk and participants who carried heterozygous 1277 C allele were at 2.22-fold higher risk for AMD than participants who carried no 1277 C allele. On the other hand, A69S T allele was also found to be associated with AMD in the same population. The A69S T allele was associated with a 3.00-fold increased risk for AMD group. Participants, who carried homozygous T allele, were at 8.61-fold higher risk and participants who carried heterozygous GT alleles were at 1.88-fold higher risk for AMD than participants who carried no T allele. The low frequency of the homozygous risk allele (6.7%) might have resulted in deviation from Hardy–Weinberg equilibrium in the control group, whereas the AMD group remained in equilibrium. It should be noted that biological and nonbiological reasons could cause deviation from Hardy–Weinberg equilibrium for a population (Lee 2003). For the control group, it could be attributed to nonrandom selection, because enlisted subjects had been selected from characteristically healthy samples. Because no genetic data were reported from Turkey on the Y402H and LOC387715 polymorphisms in Turkish AMD patients, we could not compare the prevalence of these polymorphisms in our study population. The prevalence of AMD seemed to vary around the world by ethnic group, but direct comparison among epidemiological studies could not be performed because of differently designed studies. Souied et al. (2005) reported ORs of 3.0- and 6.93-fold in French neovascular AMD patients with the TC and CC genotypes, respectively. In a population-based study in Latinos, Tedeschi-Block et al. (2007) suggested that CFH was likely to be involved in the pathogenesis of bilateral, but not unilateral, early AMD. They reported ORs of 1.2- and 1.6-fold with the TC and CC genotypes, respectively. According to the study by Varma et al. (2004), non-Hispanic Whites had a higher prevalence and age-specific rates of chroidal neovascularization than Latinos. ORs revealed by Simonelli et al. (2006) in the Italian population were 1.4- and 3.9-fold in AMD patients with the TC and CC genotypes, respectively. Gotoh et al. (2006) found that the frequency of the C allele of rs1061170 was much lower (0.04) in Japanese controls than in Caucasians (0.45) and concluded that CFH did not appear to be a primary hereditary risk factor to AMD in Japanese. Baird et al. (2006) found 1.86- and 9.26-fold increase of risk for TC and CC genotypes, respectively, for Anglo-Celtic ethnic population in Australia. It was reported that in U.S populations, the presence of TC genotypes resulted in an increased risk of AMD of between 2.45- and 4.6-fold and CC genotypes resulted in an increased risk between 3.33- and 7.4-fold (Klein et al., 2005; Zareparsi et al., 2005; Baird et al., 2006). In the present study, the ORs were 2.22- and 4.12-fold with the TC and CC genotypes, respectively, for Y402H polymorphism similar to reports for Caucasian populations (Table 5).
ORHET refers to comparison of TC with TT, and ORHOM refers to comparison of CC with TT. C allele is the risk allele. ORHET refers to comparison of GT with GG, and ORHOM refers to comparison of TT with GG. T allele is the risk allele.
(1) Sepp et al., 2006; (2) Magnusson et al., 2006; (3) Haines et al., 2005; (4) Seddon et al., 2007; (5) Chu et al., 2008; (6) Bergeron-Sawitzke et al., 2009; (7) Pulido et al., 2007; (8) Seitsonen et al., 2008; (9) Rivera et al., 2005; (10) Schmidt et al., 2006; (11) Ross et al., 2007; (12) Hughes et al., 2007; (13) Hageman et al., 2005; (14) Conley et al., 2005.
Rivera et al. (2005) reported ORs as 2.69- and 8.21-fold in Caucasian AMD patients with the GT and TT genotypes for the second risk factor gene LOC387715, respectively. ORs revealed by Pulido et al. (2007) in the Caucasian population were 3.35- and 7.75-fold in AMD patients with the GT and TT genotypes, respectively. Jiang et al. (2009) reported a case–control study in a Chinese cohort of individuals with exudative AMD and observed 3.4- and 10.76-fold increased risk with GT and TT genotypes, respectively. They concluded that a higher frequency of homozygous risk genotypes and risk haplotype and a lower frequency of the protective haplotype in the Chinese might be the cause of higher prevalence of exudative AMD. In our study, the frequency of homozygous risk genotypes was 8.61 and the frequency of protective genotype was 1.88, which were in concordance with Caucasian and Chinese risk frequencies. In the comparison of the control group and AMD group allele distribution, the OR of the T risk allele was 3.0-fold (p=0.0001). For the AMD group, G and T allele frequencies were 49.7% and 50.3%, respectively. But in the control group, G and T allele frequencies were 74.8% and 25.2%, respectively, which demonstrated the protective role of the G haplotype. CFH Y402H and LOC387715 polymorphisms were involved in macular degeneration in Caucasian European AMD patients. A hypothesis proposed was that inflammation and other factors of the immune response might play an important role in the stimulation and development of AMD (Hageman et al., 2001). CFH protects the host arterial wall from excess complement activation. By altering this ability, vessel damage might trigger neovascularization, causing drusen deposits in the macula of AMD patients (Penfold et al., 2001; Hageman et al., 2005). The LOC387715/HTRA1 locus was a second major locus contributing to AMD pathogenesis (Rivera et al., 2005), but the results indicated an independent contribution of effects of risk alleles at the LOC387715 and CFH gene locus to the overall disease risk (Scholl et al., 2007). Heritability for AMD was estimated to range from 46% to 71% (Scholl et al., 2007); first-degree relatives of AMD patients were at increased risk (OR, 2.4-fold) for AMD. Even when the patient was affected at a young age, late AMD lifetime risk was increased (OR, 4.2-fold) (Seddon et al., 1997; Klaver et al., 1998). The prevalence of AMD varies among different ethnicities (Frank et al., 2000). Caucasians were more often affected with AMD than black people, supporting the data of the studies showing that AMD begins earlier and occurs more frequently in less-pigmented eyes (Weiter et al., 1986). This Turkish population reflected the characteristics of Caucasian populations, and the prevalence of genetic risk factors for AMD in the present study supported the results of AMD genetic risk frequencies in Caucasian populations.
Conclusion
Determination of high-risk genotypes and phenotypes may prove valuable to the development of point-of-care technology for the treatment of AMD and may become a model for personalized medicine. If these genetic associations are confirmed, rapid and cost-effective assays can be developed for determining whether an individual has the T allele at LOC387715 and/or the C allele at Y402H, and based on an individual's phenotype as well as genotype, preventive therapeutic treatments may be recommended (Postel et al., 2005). In our study, the control group should be investigated in a longitudinal study to determine whether their eyes develop AMD in the future.
Footnotes
Acknowledgment
This study was supported by the Afyon Kocatepe University Research Fund with code number 08.TIP.08.
Disclosure Statement
There is no conflict of interest or competing financial disclosure statement between authors.