
Abstract
In many cases, bacterial pathogens are close relatives to nonpathogens. Pathogens seem to be limited lineages within nonpathogenic bacteria. Nonpathogenic isolates are generally more diverse and widespread in the environment and it is generally considered that environmental bacteria do not pose a risk to human health as clinical isolates do; this may not be the case with mycobacteria, but environmental mycobacteria have not been well studied. It is documented that several environmental mycobacteria constitute a source for human infections. Diverse mycobacterial environmental isolates are rarely involved in human disease. Environmental mycobacteria may have a role in degradation of different compounds. Environmental mycobacteria have had a long interaction with humans, maybe as long as the human species, and may have contributed to human evolution.
Introduction
Excellent reviews have been published on environmental and nontuberculous mycobacteria (Falkinham, 2003; Primm et al., 2004; van Ingen et al., 2009; Alvarez-Uria, 2010; Kazda et al., 2009) and on the pathogenic mycobacteria (Cosma et al., 2003). The information contained therein will not be repeated here and we will only cover the pertinent literature addressing the new questions posed here.
A Context of Different Environmental and Clinic Bacteria
In a comparative genomic analysis of klebsiella (Reyes-Prieto et al., unpublished), we found that there was an uneven distribution of virulence genes in environmental and clinical klebsiellas. There were virulence genes in klebsiellas obtained from the environment, so it is not assured that environmental bacteria would be completely safe. However, Klebsiella variicola plant isolates (from maize plants) had fewer virulence genes than clinical Klebsiella pneumoniae isolates. A different epidemiological dynamics has been suggested for K. pneumoniae and K. variicola (Martínez et al., 2004). K. pneumoniae isolates are transmitted from human to human and K. variicola isolates most probably from the environment to human. K. variicola isolates fix nitrogen, but this is not a characteristic of K. pneumoniae (Rosenblueth et al., 2004). Nitrogen fixation is advantageous in the environment, especially in association with plants but probably not so in human patients. In Klebsiella, genes determining nitrogen fixation as well as genes involved in cellulose or pectin (natural products existing only in plants) degradation allowed us to recognize that the human-infecting K. variicola should be environmental borne (Martínez et al., 2004). Nitrogen fixing genes may be easily lost when not needed. A K. variicola strain isolated as a maize endophyte promoted increased crop yields when used in agriculture as inoculant for wheat and maize in the United States (Iniguez et al., 2004). This strain was no longer used in agriculture after virulence genes were detected in its genome.
Interestingly, genes that are found in human bacterial pathogens (considered virulence determinants) such as type III secretion systems are also required in plant colonization in Rhizobium (Marie et al., 2001), a well-known plant mutualist that has been safely used in agriculture as plant inoculants for over 100 years. In Azoarcus, a rice endophyte with promising biotechnological applications, cable pili are needed for plant colonization (Böhm et al., 2007), and they are required for epithelium colonization by pathogenic bacteria (Urban et al., 2005). Some plant-associated bacteria with genes for eukaryotic infection could be potential human pathogens and sick people, especially immune-compromised patients, could be at risk. Bacterial species from Stenotrophomonas, Serratia, Klebsiella, and Salmonella enterica Typhimurium that have been reported as plant endophytes (Araujo et al., 2001; Gyaneshwar et al., 2001; Dong et al., 2003; Barac et al., 2004; Martínez et al., 2004; Rosenblueth et al., 2004) are also human pathogens or opportunistic pathogens. The use of potential human pathogens as crop inoculants is not recommended as an agricultural practice (Parke and Gurian-Sherman, 2001; Rosenblueth and Martínez-Romero, 2006).
Bacteria that are able to use many nutrients and are highly resistant to different stresses may be capable to colonize many plants and it is not surprising that some endophytes (associated bacteria inside the plant) may be human pathogens or opportunistic human pathogens (Rosenblueth and Martínez-Romero, 2006); a similar argument has been considered to explain that very successful biocontrol agents are opportunistic human pathogens (Parke and Gurian-Sherman, 2001). Among rhizospheric (on roots) bacteria, opportunistic human pathogens are commonly found as well (Berg et al., 2005).
The Amazon River, the water of which is considered suitable for human consumption, contains Chromobacterium violaceum, which confers the river its color by producing violacein. It has been reported that C. violaceum or Agrobacterium (plant pathogen relative to Rhizobium) may cause serious human disease in rare cases (Alnor et al., 1994; Teoh et al., 2006). Nevertheless, clinical agrobacteria are phylogenetically distant from other agrobacterial species (Aujoulat et al., 2011) and a similar situation seems to occur in Burkholderia. In burkholderias, most of the highly pathogenic bacteria are a distinct phylogenetic group from the mostly nonpathogenic plant and other environmental isolates (Caballero-Mellado et al., 2004). We have isolated Acinetobacter as plant endophytic bacteria from bean seeds (López-López et al., 2010) that are not closely related to the clinical Acinetobacter isolates (Dijkshoorn et al., 2007). Only some strains of Vibrio cholerae are highly virulent and there is a larger diversity of nonpathogenic environmental isolates (Faruque et al., 2004). Most nonpathogenic mycobacteria are phylogenetically separated from the slow-growing mycobacteria that contain mostly pathogens (M. tuberculosis, M. leprae, M. bovis). This contrasts to rhizobia, wherein no human pathogens occur in the slow-growing group (Bradyrhizobium), but have been detected in the fast-growing bacteria related to Agrobacterium, which produce tumors in plants (Aujoulat et al., 2011). Afipia, close relatives to bradyrhizobia, have been regularly recovered from various clinical samples and may be associated with nosocomial infections (La Scola et al., 2002).
Previously described examples illustrate that some clinical isolates are distinct from environmental populations. On the other hand some environmental isolates and human pathogens may be undistinguishable. In Burkholderia, clinical strains have been reported to group with rhizospheric and bioremediation strains (Baldwin et al., 2007). Burkholderia vietnamiensis is a nitrogen-fixing rice isolate that has been used for rice fertilization (Govindarajan et al., 2008). Recently, its environmental use has been banned in many countries because of its belonging to the cepacia complex that poses a high risk for people with cystic fibrosis (Holmes et al., 1998). Similarly, environmental and clinical Pseudomonas aeruginosa were equally virulent in model hosts (Vives-Flórez and Garnica, 2006).
Environmental Mycobacteria and Human Disease
Mycobacteria are very diverse, with over 140 species currently reported. In recent years, a large number of new mycobacterial species have been described from the environment. There are indications that environmental mycobacterial diversity is much broader than we suspected (Primm et al., 2004; our own unpublished results). There is compelling evidence that human mycobacterial infections may be acquired from the environment (Wolinsky, 1979; Primm et al., 2004; De Groote et al. 2006; van Ingen et al., 2009). However, it is clear that as mycobacteria seem to be everywhere human contact with environmental mycobacteria is inevitable (the same holds for P. aeruginosa) and the mild transient infections that mycobacteria may produce may be asymptomatic and self-cured. Nevertheless, nontuberculous mycobacteria are increasingly encountered as human pathogens (van Ingen et al., 2009). Around 20% of the species have been isolated both from clinics and the environment. Around 30% of mycobacterial species (the numbers may vary in the different surveys, and many environmental isolates have not been designated as formal species) have not been isolated from clinics but only from the environment. Many have not been characterized with regard to their effects on human health. Mycobacterial species considered to be innocuous (Tortoli, 2003) are indicated in a phylogenetic tree (Fig. 1). Many environmental isolates belong to the group of fast-growing mycobacteria (Pitulle et al., 1992), but they have also been detected in the slow-growing group.
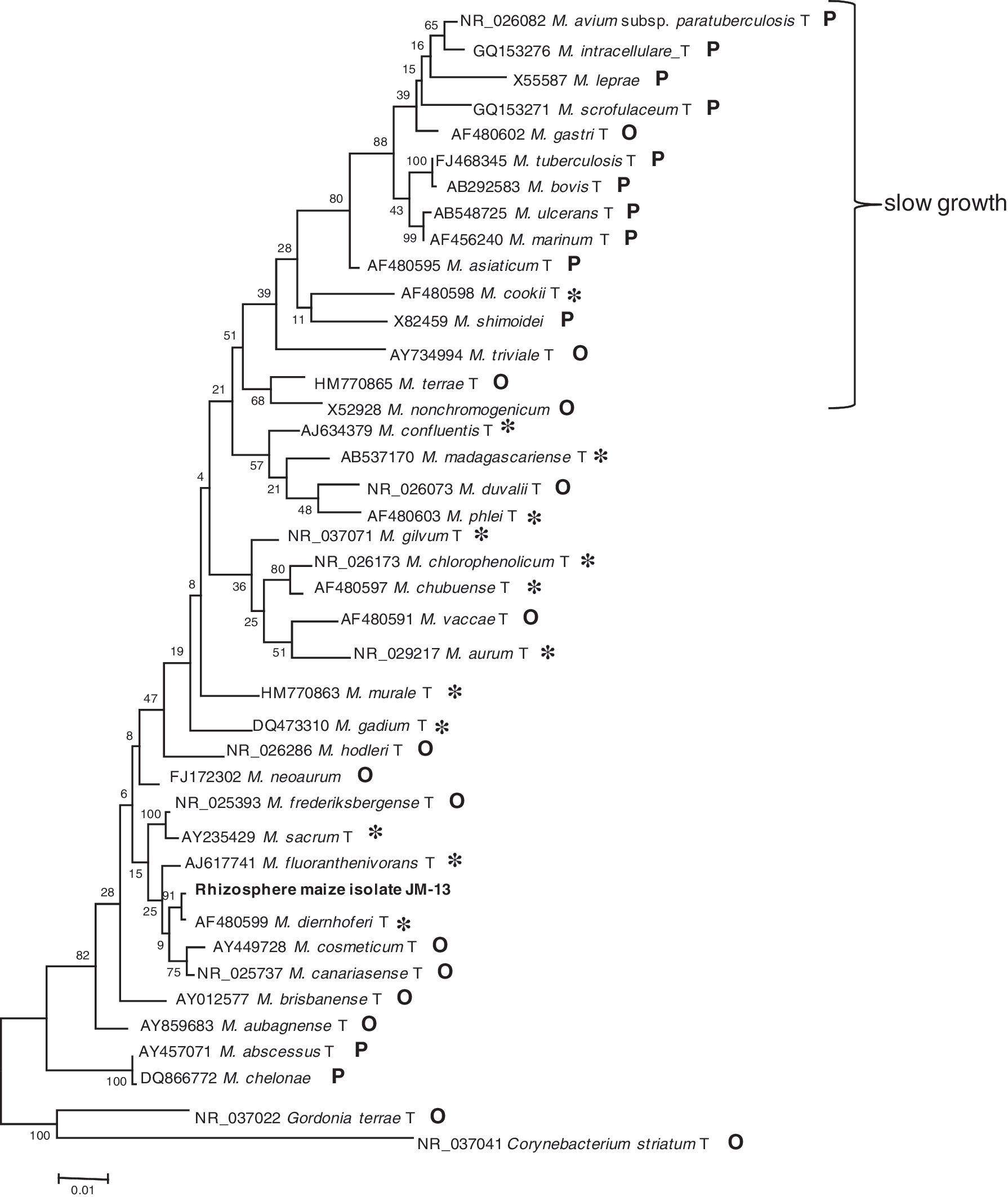
16S rRNA phylogeny of some Mycobacterium, species constructed with maximum likelihood under model GTR and rooted with Gordonia terrae and Corynebacterium striatum. T, type strain; *, nonpathogenic, purely environmental; O, oportunistic or rare pathogenic; P, highly pathogenic. Bolded terms indicate maize rhizosphere.
Environmental mycobacteria are found in soil, rivers, oceans, pipes distributing drinking water (Falkinham et al., 2001; Le Dantec, 2002; Dailloux et al., 2003; September et al., 2004; Young et al., 2005), or sewage and have also been found in bath showers and other manmade sites. It has been recommended that people with lung diseases avoid mycobacterial aerosols (Falkinham, 2003) from contaminated sources. Hospital shower mycobacteria are the same strains as those found in patients and have been recognized as a source of nosocomial infections (Falkinham et al., 2008).
Mycobacterium avium is found in soil and dust and may survive for a long time outside hosts (Whittington et al., 2004). Recently, an increase (not due to better sampling or detection procedures) has been found in infections with M. avium in the aging population from The Netherlands with chronic obstructive pulmonary disease (van Ingen et al., 2010a). M. avium strains have been isolated from patients with AIDS (Bellamy et al., 2004) and seem to be a major cause of death. The high incidence of M. avium infections is correlated with high numbers of these bacteria in environmental or drinking water (Glover et al., 1994; Pierce, 2009), but dust from potting soil (especially peat) may also be a source of human infections (De Groote et al., 2006; De Groote and Huitt, 2006). High incidence of mycobacterial infections have been found in gardeners. The use of protective antidust masks is recommended. Genomic islands have been identified in M. avium subspecies paratuberculosis, which is implicated in cases of Crohn's disease, but are not present in M. avium subspecies avium, which infects immunocompromised patients (Wu et al., 2006). Environmental or human M. avium are not the same populations as those obtained from birds (Johansen et al., 2009).
Mycobacteria have also been found in swimming pools (Leoni et al., 1999). Otomastoiditis has been detected in children and is caused by Mycobacterium abscessus, and it is attributed to their swimming in pools (van Ingen et al., 2010b). Finding mycobacteria in such environments is related to their resistance to chloride (Taylor et al., 2000) and the capacity of mycobacteria to make biofims (Schulze-Robbecke et al., 1992; Ojha et al., 2005). Bacteria in biofims are highly resistant to different antimicrobials (Stewart and Costerton, 2001).
Pathogens may have their reservoirs in the environment and this may be a source of human infections, as is the case with Mycobacterium ulcerans, which thrives in water plants and aquatic insects (Marsollier et al., 2002, 2004; Mosi et al., 2008).
In the most acid river in the world, the Tinto River in Spain, a novel species was identified resembling Mycobacterium shimoidei (Garcia-Moyano et al., 2007). M. shimoidei was found as a rare clinical isolate (Tsukamura, 1988) and has been also found in water distribution systems. From the Thames River in London, Mycobacterium fluoranthenivorans was identified (Boden et al., 2008); this species had been isolated from contaminated soil with polycyclic aromatic hydrocarbons from an abandoned gas-producing mine in Germany (Hormisch et al., 2004).
The Han River in Seoul Korea was studied with a culture-independent approach using a restriction enzyme analysis of the hsp65 gene obtained by PCR and its sequence (Lee et al., 2008). M. gordonae, M. terrae, M. kumamotonense, M. peregrinum, M. intracellulare, M. holsaticum, M. aichiense, M. phocaicum, M. arupense, M. brisbanense, M. chubuense, M. gilvum, M. lentiflavum, M. gadium, and other nonclassified Mycobacterium sp. were identified. River-borne mycobacteria include bacteria that are not innocuous.
Using culture-dependent and -independent approaches (with rpoB- and hsp65-specific primers and sequencing), we analyzed mycobacteria (Sachman-Ruiz et al., 2009) from two rivers in Mexico, one highly polluted (Apatlaco River) and the other not (Tembembe River). A large diversity of fast-growing mycobacteria and also M. avium was detected in the nonpolluted Tembembe River and in the Apatlaco River, which crosses an important urban area in Cuernavaca, Morelos in Mexico. It is unknown whether mycobacteria, especially M. avium, from the polluted Apatlaco River, which is in close contact with a large population, may be a source of human infections. Our data highlighting a potential risk prompted the authorities to start building collectors and sewage plants for these waters.
Mycobacteria have been reported in a river at the Mexican-United States border, Rio Grande (Bland et al., 2005). Mycobacterial presence was correlated with enterobacterial numbers meaning that this river carried bacteria excreted by humans. In the United States, new rules allow 0 enterobacteria per 100 mL potable water.
In developed countries, lymphadenitis in babies and small children are due to mycobacterial infections in mandibular and lymph nodes (Wolinsky, 1995); surprisingly, there has been a shift in the species involved, from Mycobacterium scrofulaceum to M. avium (Wolinsky, 1995). This shift has been related to the increases of M. avium in water (Primm et al., 2004).
Mycobacteria in Human Evolution
As there is now evidence that different rivers (Boden et al., 2008; Lee et al., 2008; Sachman-Ruiz et al., 2009) and lakes (van Ingen et al., 2009) around the world naturally contain mycobacteria, it seems probable that humans who have drunk water from rivers and lakes for hundreds of thousand years have ingested mycobacteria for most of our living history. As mycobacterial composition may not be the same everywhere and may depend on environmental conditions, local human populations may have been selected for their resistance to the local mycobacteria. In a sense, this must have shaped human evolution, and this can also be applied to many other bacteria of which we know little about. A closer contact with soil and soil mycobacteria may have arisen with agriculture, dating from only around 10,000 years ago, followed by a rise in contact with farm animals and their bacteria.
Mycobacteria in Plants
Mycobacteria have been found as wheat endophytes (Conn and Franco, 2004). When studying the bacterial community associated with plant roots we found mycobacteria related to Mycobacterium diernhoferi in the maize rhizosphere (Fig. 1). This species is considered nonpathogenic and has been previously found together with Mycobacterium vanbaalenii, Mycobacterium mageritense, Mycobacterium austroafricanum, Mycobacterium chubuense, or Mycobacterium chlorophenolicum in contaminated soils (Cheung and Kinkle, 2001).
Upon leaving Cuernavaca, the Apatlaco River is used for irrigation of a large agricultural area where maize and other crops are grown. We wondered whether mycobacteria could proliferate in plants irrigated with mycobacteria-contaminated water. A pending question in mycobacterial research is which are the actual sites of mycobacterial replication. Bacteria form biofilms on plant roots and mycobacteria seem to be seed species for biofilm formation in different niches. We found no mycobacteria on maize roots irrigated with the contaminated river water (from the Apatlaco River crossing Cuernavaca) in laboratory assays using culture-dependent and -independent approaches. A large diversity of other bacteria was encountered in the irrigated roots but not in control plants (not shown).
Beneficial Mycobacteria
Kazda et al. (2009) published a review on the biological role of mycobacteria in the environment. Some mycobacteria have been recognized as having biotechnological advantages because of their capacity to degrade petroleum-contaminated soils and may be used in bioremediation (Cheung and Kinkle, 2001; Coleman and Spain, 2003; Hormisch et al., 2004; Leys et al., 2005; Hennessee et al., 2009).
Among the fast-growing mycobacteria, Mycobacterium vaccae is used in vaccination (Yang et al., 2010). Environmental mycobacteria could stimulate immunity against pathogenic mycobacteria and help build resistance to them (Black et al., 2001; Fine, 2001); they possibly prevent asthma or allergies (Wang and Rook, 1998; Black, 2001). The health impacts of environmental bacteria were extensively reviewed by Primm et al. (2004). It was also reported that environmental mycobacteria interfere with the response to vaccination (Brandt et al., 2002; Young et al., 2007; Mendoza-Coronel et al., 2011).
Environmental Mycobacteria Predisposition for Human Infections
It seems that several environmental isolates have genes that predispose them to infect humans. General characteristics of mycobacteria are their resistance to stress, extreme temperatures, acidity, chlorides, and starvation. These may be predisposing characteristics to resist defense responses during infections. Are fast-growing mycobacteria less tolerant to such stresses?
Mycobacteria are old prokaryotic lineages, and for a long time, they have probably constituted prey for protozoa. Many are able to survive inside amoebas, and consequently, they have a long experience colonizing and persisting inside eukaryotic cells (Harriff et al., 2008).
Mycobacteria, like other bacteria, have genetic plasticity, their genes may be lost or acquired, and genetic variation may determine degrees of pathogenicity (Brosch et al., 2001; Bhatt et al., 2002; van der Sar et al., 2004; Wang and Derbyshire, 2004; Gutierrez et al., 2005). Environmental mycobacteria could be recipients of pathogenicity determinants from pathogenic mycobacteria (Picardeau and Vincent, 1997).
Conclusion
Are environmental mycobacteria a threat to human health? This is still an open question and it depends on the environmental mycobacteria and as a corollary on the environment. Many environmental mycobacteria remain unknown. The human impact on mycobacterial ecology and diversity has been recognized, with human activities causing a selection in the environment of more stress-resistant mycobacterial species such as M. avium in place of M. scrofulaceum (Primm et al., 2004), and the culprit here may be the choride (added by humans) in water. For some environmental mycobacteria, there is no doubt that they are pathogens, and others may be less virulent and could even prevent pathogen infections (Fine, 2001). As discussed above, mycobacteria seem to have a great capacity to colonize humans and animals and have a long and extensive history of interactions with humans. In most mycobacterial species, the genetic basis of pathogenicity is unknown. It is difficult to definitively ascertain whether a particular strain or a newly found species from the environment may be a potential or an opportunistic pathogen. The fact that an environmental bacterium is not found in clinics does not warrant the conclusion that it is not pathogenic. Unless adequate pathogenicity tests are performed with strains to be used in bioremediation or other biotechnological applications, there should be restrictions to handle them or to deliver them to the environment. We still know very little about mycobacteria. “More information about the pathogenesis and treatment of NTM (nontuberculous mycobacteria) is needed … physicians and scientists should join efforts to … studies of specific species … in the different groups of human hosts” (Alvarez-Uria, 2010).
Footnotes
Acknowledgments
This study was supported by a grant from CONACyT (No. 48490). The authors thank Dr. M. Dunn for critically reading the manuscript and Maritza Merino Flores for her valuable technical help.
Disclosure Statement
No competing financial interests exist.