
Abstract
Recently, alterations in the expression pattern of PPP2R5C associated with malignant transformation have been characterized, and PPP2R5C overexpression was demonstrated in leukemias. To confirm the role of PPP2R5C in proliferation and its molecular mechanism, three PPP2R5C-siRNAs and a scrambled nonsilencing siRNA control were used to treat Molt-4 and Jurkat T cells. After nucleofection, PPP2R5C expression and biological consequences based on a highly efficient and specific PPP2R5C-siRNA were demonstrated by qRT-PCR, CCK-8 assay, Annexin V/PI, and flow cytometry. The global gene expression profile of PPP2R5C-siRNA-treated Jurkat T cells was established. A significant reduction in the PPP2R5C mRNA level was observed at 24 to 72 h in Molt-4 and Jurkat T cells with all of the PPP2R5C-siRNAs. The proliferation rate of Molt-4 and Jurkat T cells transfected with different PPP2R5C-siRNAs was significantly decreased at 72 h compared with the control (p<0.05). However, the transfected cells did not show a significant increase in Annexin V/PI-positive cells (apoptosis). The highly efficient PPP2R5C-siRNA2 was used to treat Jurkat T cells for gene expression profile analysis. In total, 439 genes were upregulated, and 524 genes were downregulated at least twofold in PPP2R5C-siRNA-treated Jurkat T cells. Changes in signaling pathway genes closely related to the TCR, Wnt, calcium, MAPK, and p53 signaling pathways were observed. In conclusion, the suppression of PPP2R5C by RNA interference could effectively inhibit the proliferation of leukemic T cells, the PPP2R5C-siRNA treatment altered gene expression profiles, and the differential expression of the glycogen synthase kinase 3 beta (GSK-3β), ataxia telangiectasia mutated (ATM), and Mdm2 p53 binding protein homolog (MDM2) genes may play an important role in the effects of PPP2R5C knockdown in Jurkat T cells.
Introduction
In 2001, synthetic small interfering RNAs (siRNAs) were demonstrated to be capable of inducing RNAi in mammalian cells by Thomas Tuschl (Elbashir et al., 2001). This discovery led to a surge in interest in harnessing RNAi for biomedical research and drug development. siRNA is a promising gene-targeting agent that has shown great potential, particularly in the field of cancer treatment (Devi, 2006; Oh et al., 2009; Whitehead et al., 2009). A combination of c-raf and bcl-2 siRNA induced apoptosis in HL-60, U937, and THP cell lines and increased chemosensitivity to etoposide and daunorubicin (Cioca et al., 2003). Based on the finding that B-cell chronic lymphocytic leukemia/lymphoma 11B (BCL11B) overexpression is detected in most T-cell malignancies (Oshiro et al., 2006; Liu et al., 2010; Huang et al., 2011), a previous study demonstrated that the inhibition of BCL11B expression by BCL11B-siRNA leads to apoptosis in Molt-4, Jurkat, and huT78 cell lines (Grabarczyk et al. 2007).
Recently, we characterized the novel chromosomal translocation t(14;14)(q11;q32), which causes rearrangement of the T-cell receptor (TCR) α gene J7 segment (TRAJ7) with the PPP2R5C gene (unpublished data) in a case of Sézary syndrome at the molecular level. PPP2R5C is a regulatory B subunit of protein phosphatase 2A (PP2A), which is a major cellular serine/threonine phosphatase that affects the phosphorylation status of many proteins (Muneer et al., 2002). The PPP2R5C gene encodes five differentially spliced variants: B56γ1, B56γ2, B56γ3, B56γ5, and B56γ6, whereas B56γ4 is identified only in mice. The locus for the functional PPP2R5C gene is at 14q32.2, whereas the nonfunctional B56γ1 pseudogene PPP2R5C is at 3p21.3 (Muneer et al., 2002; Lee et al., 2010). PPP2R5C plays a crucial role in cell proliferation, differentiation, and transformation based on its induction of the dephosphorylation of p53 at various residues (Shouse et al., 2008). It has been reported that the dynamic nuclear distribution of the B56γ3 regulatory subunit controls the nuclear PP2A activity and may be responsible for the tumor-suppressive function of PP2A (Lee et al., 2010). Recently, alterations in the expression pattern of PPP2R5C that are associated with malignant transformation have been characterized in lung cancer, and the PPP2R5C mutation F395C disrupts the B56γ–p53 interaction (Shouse et al., 2010).
We were also the first to find that PPP2R5C overexpression occurs in leukemias, including T-ALL (Zheng et al., 2011). To confirm the role of PPP2R5C in proliferation and its molecular mechanism, we screened highly efficient and specific PPP2R5C-siRNAs, which target different exons in the human T-ALL cell lines Molt-4 and Jurkat, and a highly efficient PPP2R5C-siRNA was used further to evaluate its molecular mechanism in the proliferation inhibition of leukemic T cells.
Methods
Cell culture
The human T-cell leukemia cell lines Molt-4 and Jurkat (Institutes for Biological Sciences Cell Resource Center, Chinese Academy of Sciences, Shanghai, China) were grown in the Roswell Park Memorial Institute (RPMI) 1640 medium (Gibco-BRL) with 10% fetal calf serum (FCS) (Sijiqing Co.) and maintained in a humidified incubator at 37°C and 5% CO2. All experiments were performed using cells in the exponential growth phase.
siRNA design and synthesis
The siRNAs PPP2R5C-siRNA1, PPP2R5C-siRNA2 (Chinese patent number: ZL 2011 1 0340411.1), and PPP2R5C-siRNA3 (Chinese patent number: ZL 201110337837.1), which target domains in the fourth, sixth, and between the eighth and ninth exons in the PPP2R5C gene (ACCESSION NM_178587), respectively, and a scrambled nonsilencing siRNA control (SC) were designed with online software (
SC, scrambled nonsilencing siRNA control.
Nucleofection
Cells were collected by centrifugation and resuspended at 2.5×106 cells for Molt-4 and Jurkat cells per 100 μL of the appropriate Nucleofector™ kit solution (Amaxa Biosystems) (Huang et al., 2011; Chen et al., 2012; Zha et al., 2012). Malignant T cells were nucleofected with 3 μg of PPP2R5C-siRNAs or a control nonsilencing scrambled (SC) siRNA using the C-005 (Molt-4 cells) or X-001 (Jurkat cells) program of the Nucleofection Device II (Amaxa Biosystems). Mock-transfected cells nucleofected without siRNA were used as a negative control. After nucleofection, the cells were immediately mixed with 500 μL of prewarmed culture medium and transferred into culture plates. The treated cells were incubated at 37°C for 3 days for cell proliferation, apoptosis, and microarray analyses. Three independent experiments for Molt-4 or Jurkat T cells were preformed every 24 h.
RNA isolation, reverse transcription, real-time qRT-PCR, and microarray analysis
Total RNA was isolated from different samples (Molt-4 and Jurkat T cells) using TRIzol (Invitrogen). cDNA for qRT-PCR was synthesized using the Superscript II RNaseH reverse transcriptase kit (Invitrogen).
The expression level of PPP2R5C and the β2-MG reference gene was determined by SYBR Green I real-time PCR. PCR was performed as previously described (Zheng et al., 2011). The sequences for the primers used in qRT-PCR were as follows: PPP2R5C: 5′-GTAATAAAGCGGGCAGC AGG-3′ (forward) and 5′-CAAAGTCAAAGAGGACGCAACA-3′ (reverse) for PPP2R5C gene amplification and β2M: 5′-CAGCAAGGACTGGTCTTT CTAT-3′ (forward) and β2M 5′-GCGGCATCTTCAAAC CTC-3′ (reverse) for β2M gene amplification.
Total RNA (>3 μg) was sent for global gene expression profile analysis using the Affymetrix HG-U133 Plus 2.0 gene chip (Shanghai Biochip Co. Ltd). Affymetrix microarray analysis was performed using the Gene Spring GX11.0 software (Agilent Technologies) (Lai et al., 2011; Huang et al., 2011a; Chen et al., 2012). Probe sets displaying a signal log ratio indicating an increase or marginal increase [i.e., log ratio ≥1(n)] and the detection of an experimental group displaying a signal change with P represented upregulated genes. Conversely, probe sets displaying a signal log ratio indicating a decrease or marginal decrease [i.e., log ratio ≤−1(n)] and the detection of a control group displaying a signal change with P represented downregulated genes. The resulting data were analyzed using the SBC Analysis System. After normalization and correction, the log2 fluorescence intensity value for each gene was obtained (Huang et al., 2011b). The fold change for all genes was calculated by comparing PPP2R5C-siRNA2- and SC-treated Jurkat T cells. A minimum twofold difference was considered significant.
Cell proliferation assays
The proliferation of Molt-4 and Jurkat T cells was indirectly assayed using the CCK-8 kit (Dojindo), which stains living cells. After transfection, approximately 5×104 Molt-4 or Jurkat (100 μL) and control cells were incubated in triplicate in 96-well plates. At 24, 48, and 72 h, the CCK-8 reagent (10 μL) was added to each well and incubated at 37°C for 2 h. The optical density at 450 nm was measured using an automatic microplate reader (Synergy4; Bio-Tek).
Apoptosis analysis
Molt-4 and Jurkat T cells transfected with PPP2R5C-siRNAs or Alexa Red Oligo were monitored from 48 to 72 h post-transfection using a Delta Vision high-resolution imaging system (Applied Precision, Inc.). The Molt-4 and Jurkat T cells (5×105) in each group were collected 48 h after transfection and then prepared with FITC-labeled anti-Annexin-V (BD Pharmingen) and propidium iodide (Kaiji) according to the manufacturers' protocol and measured by flow cytometry (Beckman Coulter). The results were analyzed using Windows MDI 2.9 software.
Statistical analyses
Statistical analyses were performed with paired t-tests and one-way analysis of variance using SPSS 13.0 statistical software. Kruskal–Wallis analysis was used to analyze the PPP2R5C gene mRNA levels in different samples. Differences were considered statistically significant at p<0.05.
Results
PPP2R5C-specific siRNAs suppress PPP2R5C expression in Molt-4 and Jurkat T cells
We first verified the transfection efficiency with Alexa Red Oligo-transfected Molt-4 and Jurkat T cells, which was found to be 58.12%±14.14% and 65.2%±10.3%, respectively (Fig. 1). To examine the knockdown of PPP2R5C expression in Molt-4 and Jurkat cells after siRNA treatment, PPP2R5C mRNA expression was analyzed by qRT–PCR 24, 48, and 72 h after nucleofection. The expression level of PPP2R5C was significantly decreased in Molt-4 and Jurkat T cells treated with all three PPP2R5C-siRNAs compared with the control (p<0.05) (Fig. 2) and the expression levels of PPP2R5C were obviously decreased (Figure S1; supplementary materials are available online at
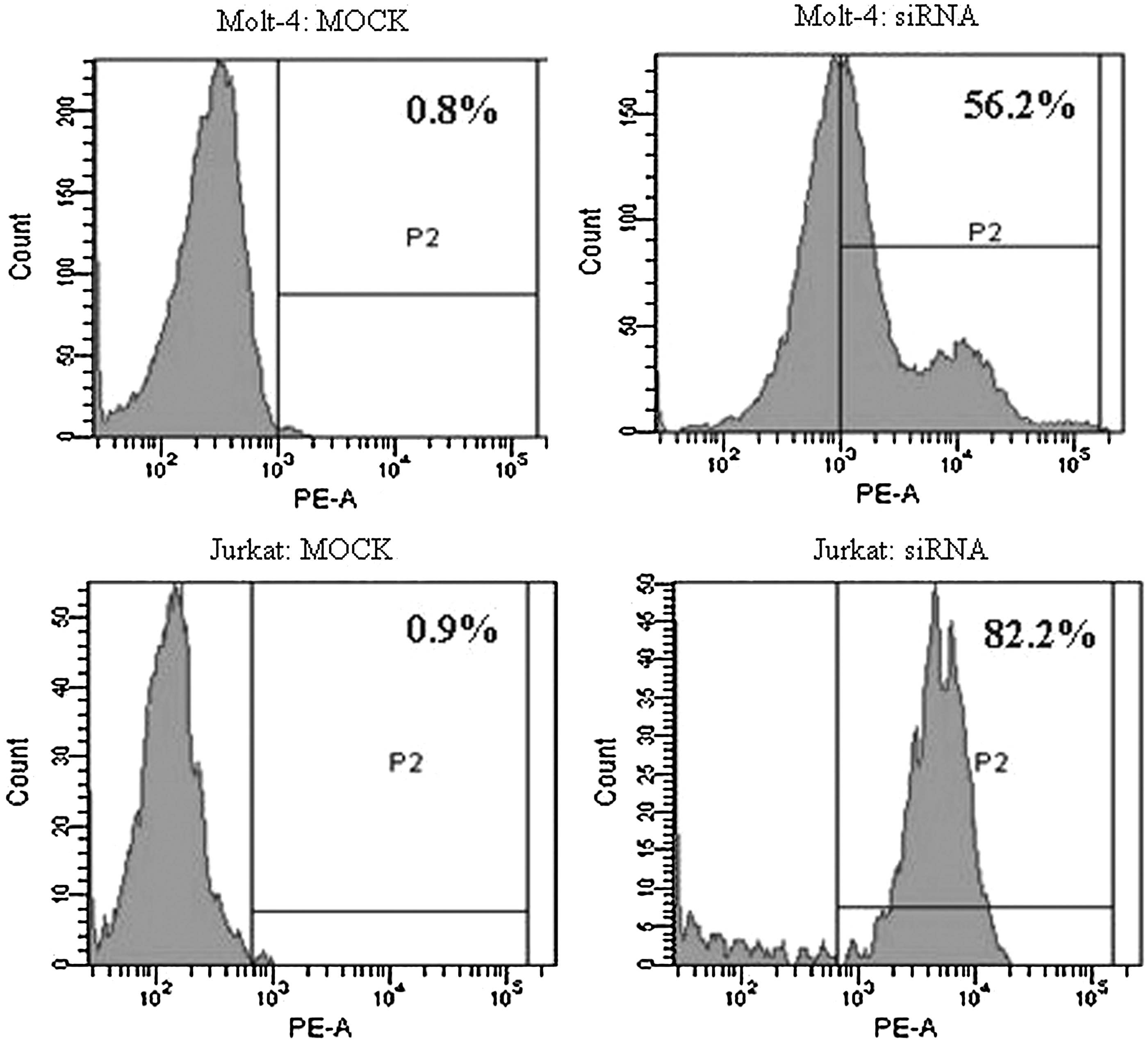
Alexa Red Oligo-transfected Molt-4 and Jurkat T cells and MOCK transfected control cells 10 h after transfection as measured by FCM (positive cells are shown in the P2 domain).
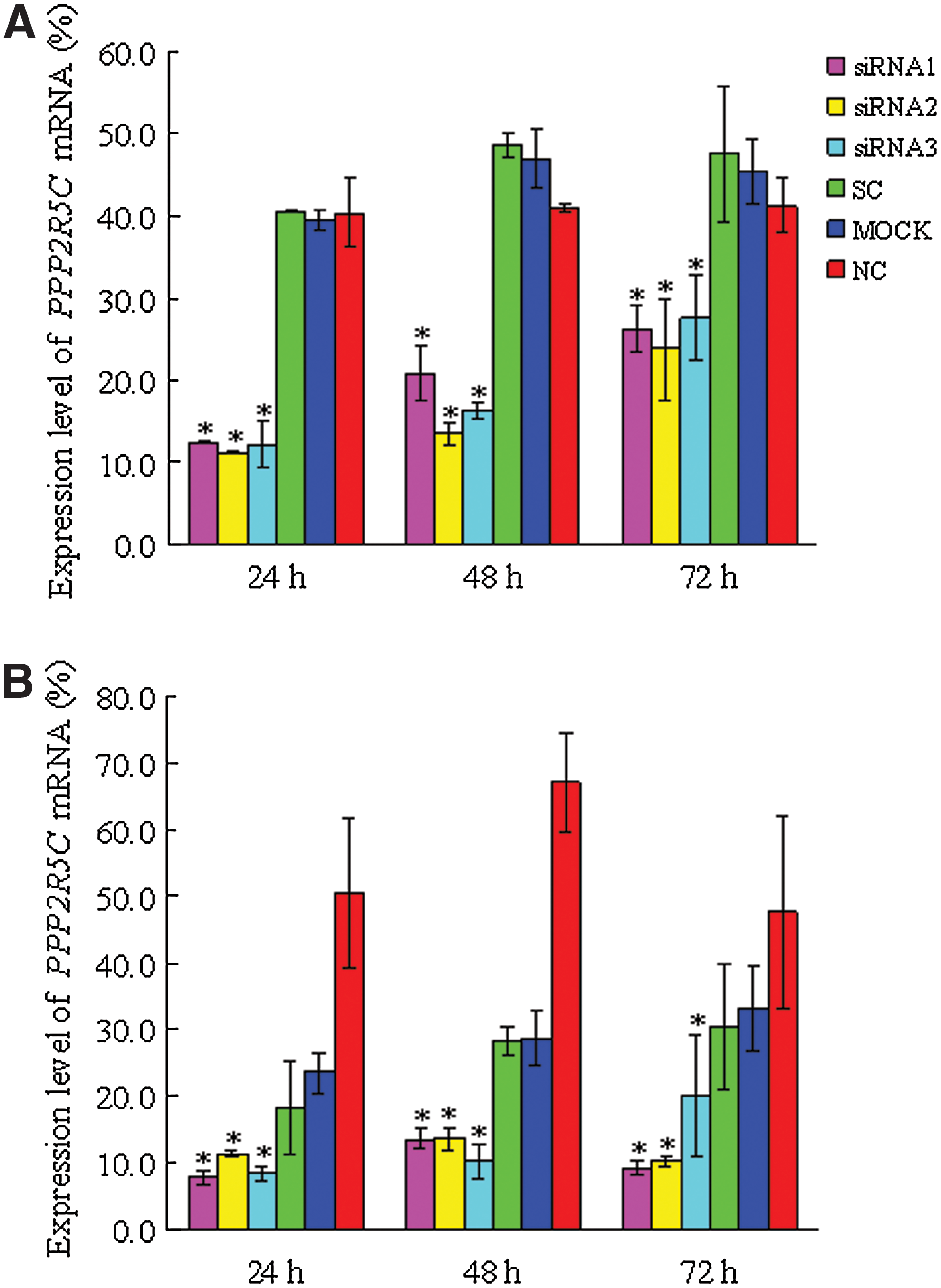
Inhibition of PPP2R5C expression in Molt-4 and Jurkat cells by RNA interference. The expression of PPP2R5C in Molt-4
PPP2R5C suppression inhibits the proliferation of Molt-4 and Jurkat T cells
All three PPP2R5C-siRNAs were used to assess biological consequences. The proliferation rate of Molt-4 and Jurkat T cells transfected with different PPP2R5C-siRNAs was significantly decreased at 72 h compared with the control. In comparison with scrambled nonsilencing RNA-treated cells, Molt-4 and Jurkat T cells transfected with PPP2R5C-siRNAs had a significantly inhibited proliferation rate (p<0.05) (Fig. 3). The inhibition rate was 49.23%±2.12% for Molt-4 T cells treated with siRNA2 at 72 h and 48.87±0.69 and 47.12%±0.72% for Jurkat cells treated with siRNA1 and siRNA2, respectively, at 72 h. However, the transfected cells did not demonstrate a significant increase in Annexin V/PI-positive cells (apoptosis) at 48 or 72 h, and the apoptosis rate in cells from different groups was 12.80%±2.73% (siRNA1), 2.11%±2.75% (siRNA2), 13.35%±3.66% (siRNA3), 8.76%±2.19% (SC), 11.73%±2.11% (MOCK), and 6.60%±1.88% (NC) for Molt-4 T cells at 48 h, and 12.79%±1.39% (siRNA1), 22.41%±1.76% (siRNA2), 18.40%±6.36% (siRNA3), 11.48%±2.77% (SC), 16.60%±4.37% (MOCK), and 7.38%±2.45% (NC) for Jurkat T cells at 48 h. We have also checked the change of cell cycle of the siRNA-treated T cells by flow cytometry; there were no significant differences between different groups at 48 and 72 h after siRNA treatment and controls either in Jurkat or Molt-4 T cells (Figures S2 and S3).
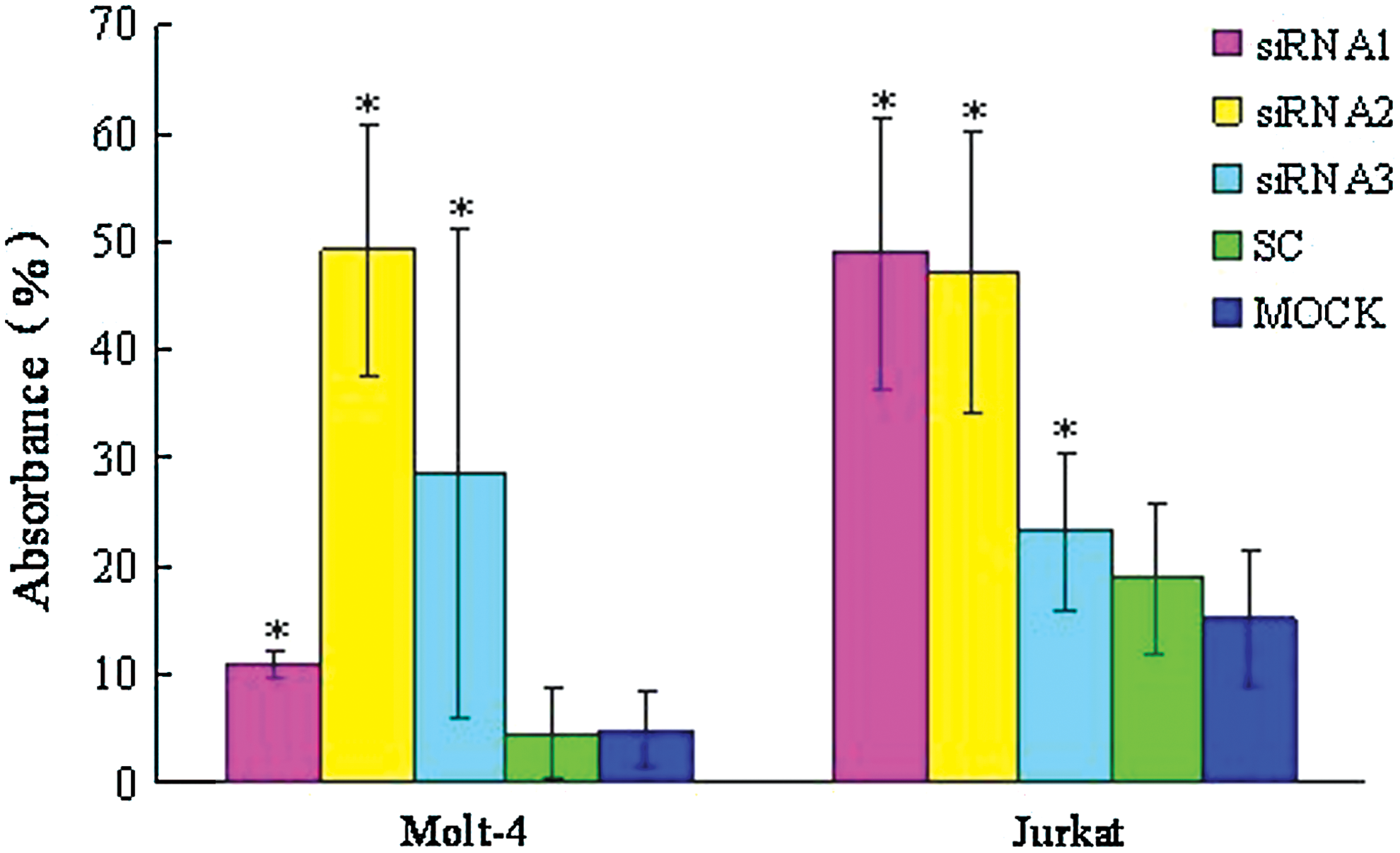
The inhibition of the proliferation of Molt-4 and Jurkat T cells transfected with PPP2R5C-siRNAs at 72 h as measured by the CCK-8 method. *p<0.05 compared with scrambled, nonsilencing siRNA-treated cells. Color images available online at
Global gene expression profile in PPP2R5C-siRNA2-treated Jurkat T cells
To elucidate the gene expression profile of Jurkat T cells after PPP2R5C suppression, global gene expression analysis was performed by comparing the transcriptome profiles of PPP2R5C-siRNA2- and SC-treated Jurkat cells using Affymetrix HG-U133 Plus 2.0 arrays. There was a clear dysregulation in global gene expression in Jurkat T cells with PPP2R5C knockdown as demonstrated by the degree of differential expression between the PPP2R5C-siRNA2 and SC groups (Fig. 4A). Subsequently, the dysregulated genes were clustered based on their differential expression (twofold up or down) and visualized using a heatmap (Fig. 4B). Gene Spring GX software analysis demonstrated that 439 genes (greater than twofold: 304 genes, greater than fourfold: 74 genes, greater than eightfold: 61 genes) were upregulated, and 524 genes (greater than twofold: 369 genes, greater than fourfold: 86 genes, greater than eightfold: 69 genes) were downregulated at least twofold in PPP2R5C-siRNA2-transfected Jurkat T cells compared with the SC control. Changes in signaling pathway genes closely related to cell proliferation, TCR signaling, Wnt signaling, calcium signaling, MAPK signaling, and p53 signaling were further analyzed by pathway analysis using the SBC Analysis System (Table 2).
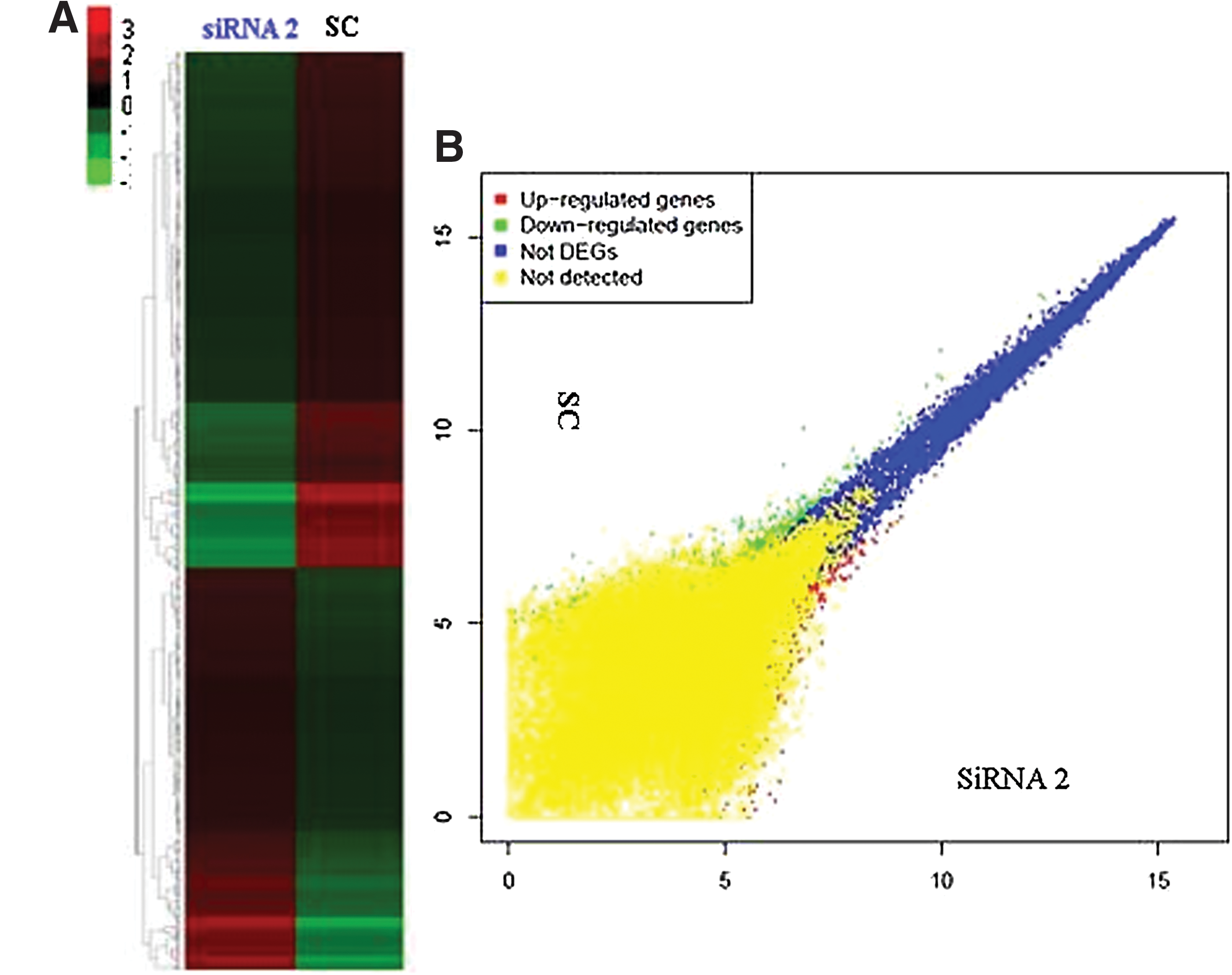
Microarray analysis after PPP2R5C knockdown in Jurkat T cells.
Discussion
PPP2R5C plays a crucial role in cell proliferation, differentiation, and transformation based on its induction of the dephosphorylation of p53 at various residues (Shouse et al., 2008) and may be responsible for the tumor-suppressive function of PP2A (Lee et al., 2010). To confirm the role of PPP2R5C siRNA on the inhibition of leukemic T cells and its potential as a therapeutic agent, we compared different PPP2R5C-siRNAs that target different exon sequences and screened leukemic T cells with a highly efficient and specific PPP2R5C-siRNA.
Exogenous siRNA delivery always results in a transient RNAi effect. In general, RNAi effects are detected between 24 to 72 h after siRNA transfection. We demonstrated that the siRNAs effectively silenced PPP2R5C post-transcriptionally, and the control siRNA had no obvious influence 72 h after nucleofection. These results were not only confirmed by real-time PCR, but also by microarray analysis, which showed that PPP2R5C was downregulated 2.49- and 2.77-fold by two probe sets. siRNAs targeting different exon domains had different efficacies for PPP2R5C gene silencing and subsequent biological consequences. PPP2R5C-siRNA2, which targets exon six, demonstrated robust knockdown, whereas PPP2R5C-siRNA1 and PPP2R5C-siRNA3 had different effects on the different leukemic cell lines.
There are no reports regarding the effects of the suppression of PPP2R5C on changing cell biological functions. In this study, we demonstrated that the suppression of PPP2R5C by RNAi effectively inhibited the proliferation of the Molt-4 and Jurkat cell lines. However, unlike other reported siRNAs, such as BCL11B-siRNA and Notch1-siRNA (Guo et al., 2009; Huang et al., 2011), the suppression of PPP2R5C by RNAi could not significantly induce apoptosis, which was confirmed by gene expression profile analysis demonstrating that only a limited number of apoptotic genes are altered. Therefore, it is interesting to analyze the molecular mechanisms of the PPP2R5C siRNA-mediated suppression of cell proliferation.
To characterize the effects of PPP2R5C knockdown in Jurkat T cells, we analyzed gene expression profiles by gene chip. Data from global gene expression analysis demonstrated that approximately 1000 genes were differentially expressed; thus, we attempted to find genes involved in signaling pathways related to the downregulation of PPP2R5C.
Genes altered in the TCR signaling pathway
TCR engagement initiates intracellular signaling cascades that lead to T-cell proliferation, cytokine production, and differentiation into effector cells (Wilkinson et al., 2005). Therefore, the TCR signaling pathway is the most important pathway in T-cell activation and proliferation (Laky et al., 2006; Zha et al., 2013), and according to the KEGG (Kyoto Encyclopedia of Genes and Genomes) website, there are 108 genes involved in the TCR signaling pathway (Pathway: map04660) (Kanehisa, 2002; Zha et al., 2013). Our results demonstrated that only three genes, p21 protein (Cdc42/Rac)-activated kinase 1 (PAK1), mitogen-activated protein kinase kinase kinase 8 (MAP3K8), and glycogen synthase kinase 3 beta (GSK-3β), were downregulated in PPP2R5C-siRNA-treated Jurkat T cells (Table 1). All three genes are also involved in the MAPK signaling pathway, and GSK-3β is involved in the cell cycle pathway and is related to tumor proliferation (Varisli et al., 2011). The downregulation of GSK-3β is thought to decrease the phosphorylation of NFAT, while MAP3K8 downregulation effects the inhibition of activator protein-1 (AP-1), which is critically modulated by post-translational modifications, particularly phosphorylation, which mediates not only TCR, but also MAPK cascades (Torgersen et al., 2008). Therefore, our results indicate that the effect of the proliferation inhibition of Jurkat cells by PPP2R5C downregulation might be due to a common cell proliferation pathway, such as the MAPK pathway, rather than the TCR signaling pathway.
Genes altered in the p53 signaling pathway
p53 is an important tumor suppressor protein, in humans, whose main functions are involved in preventing genome mutations and regulating the cell cycle. In response to DNA damage and other stress signals, p53 is highly modified post-translationally, which increases its stability and promotes its activation and nuclear localization (Shouse et al., 2010; Yang and Phiel, 2010; Isin et al., 2012). B56γ-containing PP2A is an important p53 regulatory enzyme that directly regulates DNA damage-induced p53 stabilization. In response to DNA damage, B56γ-containing PP2A triggers Thr55 dephosphorylation on p53 and promotes its transcriptional activation (Li et al., 2007; Shouse et al., 2008). In addition, B56γ-containing PP2A could dephosphorylate Bcl2 and prevent the proteasome-dependent degradation of Bcl2, which inhibits p53 and resists apoptosis (Lin et al., 2006). Thus, B56γ-containing PP2As regulate the p53 network at multiple levels. In this study, the expression of p53 was not directly downregulated in Jurkat T cells by inhibiting PPP2R5C expression; however, the MDM2 gene, which is an important negative regulator of the p53 tumor suppressor, was upregulated 16.51-fold (Uhrinova et al., 2005), while the ataxia telangiectasia mutated (ATM) gene, a positive regulator of the p53 that triggers p53 Ser15 phosphorylation and promotes its transcriptional activation, was downregulated 2.1-fold (Ditch et al., 2012). Recently, it is proved that ATM directly phosphorylates and specifically regulates B56γ3, B56γ2, and B56δ, after DNA damage, the phosphorylation of B56γ3 at Ser510 leads to an increase in B56γ3-PP2A complexes, and direction of the PP2A phosphatase activity toward the substrate p53, activating its tumor-suppressive functions (Shouse et al., 2011). Both the MDM2 and ATM proteins act as an E3 ubiquitin ligase, which binds to the N-terminal domain of the p53 protein and inhibits p53 transcriptional activation (Vassilev et al., 2004). This result suggests that PPP2R5C knockdown in Jurkat T cells may lead to a decrease in the transcriptional activation of the p53 protein and p53 function by downregulating ATM and upregulating MDM2.
Genes altered in the canonical Wnt/β-catenin signaling pathway
The canonical Wnt
Genes altered in the other signaling pathways
In this study, we also analyzed the differential regulation of genes involved in the MAPK, calcium, apoptosis and cell cycle signaling pathways. Many genes were upregulated or downregulated; however, it is hard to conclusively determine which are the direct results of PPP2R5C knockdown in Jurkat T cells. This finding is in agreement with results demonstrating insignificant differences in apoptosis and cell cycle genes in Jurkat T cells after PPP2R5C knockdown.
In conclusion, our findings provide evidence for the effect of proliferation inhibition in malignant T cells by PPP2R5C knockdown and characterize its relative molecular mechanism. PPP2R5C might be involved in the p53 and canonical Wnt/β-catenin signaling pathways. The differential expression of the GSK-3β, ATM, and MDM2 genes may play an important role in the effects of PPP2R5C downregulation. Moreover, GSK-3β plays a direct role in the stimulatory modification of the p65/RelA transcription factor, which targets NF-κB p50/52 in NF-κB signaling. Overall, PPP2R5C downregulation results in the inhibition of Jurkat T-cell proliferation, which involves numerous pathways, and decreased GSK-3β may be a critical factor in this process. However, further validation of the differentially regulated genes and their relative protein expression levels is needed.
Authors' Contributions
Y.L. contributed to the concept development and study design. Y.C., S.L., Q.S., X.Z., H.Z., L.Y., S.C., X.W., and B.L. performed the laboratory studies. Y.L. and Y.C. coordinated the study and helped draft the manuscript. All authors read and approved the final manuscript.
Footnotes
Acknowledgments
This work was supported by Grants from the National Natural Science Foundation of China (30871091 and 91129720), the Collaborated grant for HK-Macao-TW of the Ministry of Science and Technology (2012DFH30060), the Guangdong Science & Technology Project (2012B050600023), Science and Technology Innovation Key Project of Guangdong Higher Education Institutes (kjcxzd1013), and the Key Discipline Construction Foundation of Jinan University.
Disclosure Statement
The authors declare that they have no competing interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.