
Abstract
Ankylosing spondylitis (AS) is a chronic inflammatory disorder primarily affecting the sacroiliac joints and the spine. T-cell immunoglobulin- and mucin-domain-containing molecule 3 (TIM-3) has been established as a negative regulatory molecule that plays a critical role in controlling inflammation. Studies have shown that polymorphisms in TIM-3 gene may be associated with inflammatory diseases. The current study investigated the association between polymorphisms in the TIM-3 gene and susceptibility to AS, and it examined the effects of these polymorphisms on gene expression. Two polymorphisms in TIM-3 −574G/T and +4259T/G polymorphisms were identified by polymerase chain reaction-restriction fragment length polymorphism in 282 AS patients and 298 healthy controls. Results showed that frequency of the TIM-3 −574GT genotype was significantly increased in cases than in controls (Odd ratio [OR]=2.50, 95% confidence interval [CI]: 1.39–4.48, p=0.002). Similarly, TIM-3 −574T allele revealed a positive association with the disease (OR=2.39, p=0.002). The TIM-3 +4259T/G polymorphism did not show any correlation with AS. We further evaluated TIM-3 mRNA and protein levels in CD4+ T cells, CD8+ T cells, and monocytes from subjects carrying different TIM-3 genotypes. Results revealed that subjects carrying polymorphic −574GT genotype had significantly lower TIM-3 mRNA and protein levels in CD4+ T cells, CD8+ T cells, and monocytes than those with wild-type GG genotype. These data suggest that TIM-3 polymorphism is associated with increased susceptibility to AS possibly by downregulating gene expression.
Introduction
A
T-cell immunoglobulin- and mucin-domain-containing molecule 3 (TIM-3) is expressed in Th1, Th17 cells, and CD8 T cells, but not in Th2 cells (Han et al., 2014; Poonia and Pauza, 2014). An Interaction between TIM-3 and its ligand galectin-9 inhibits Th1 and Th17 responses (Gorman et al., 2014) and induces peripheral tolerance (Poonia and Pauza, 2014; Qiao et al., 2014; Rong et al., 2014; Wang et al., 2014), suggesting an inhibitory role of TIM-3 in T-cell responses. TIM-3 expression has also been identified in exhausted T cells during chronic infection (Han et al., 2014; Wang et al., 2014). TIM-3-expressing CD4+ and CD8+ T cells produce reduced amounts of cytokines or are less proliferative in response to antigen (Berrocal Almanza et al., 2013; Han et al., 2013; Fourcade et al., 2014; Gorman et al., 2014). Blockade of the TIM-3 signaling pathway restores proliferation and enhances cytokine production in HIV-1-specific T cells (Berrocal Almanza et al., 2013; Han et al., 2013; Fourcade et al., 2014; Gorman et al., 2014). Recent studies have shown an important role of TIM-3 T cell exhaustion in cancer. Tim-3 and PD-1, another marker of T-cell exhaustion, are co-expressed in CD8 tumor infiltrating lymphocytes in mice bearing transplanted tumors as well as in NY-ESO-1-specific CD8+ T cells in patients with advanced melanoma (Jiang et al., 2013; Han et al., 2014; Ju et al., 2014; Moore et al., 2014). TIM-3+PD-1+ T cells exhibit the most severe exhausted phenotype as defined by failure to proliferate and produce IL-2, TNF-α, and IFN-γ. The effects of targeting TIM-3 in autoimmune diseases have recently been reported (Qiao et al., 2014; Rong et al., 2014; Wang et al., 2014). Studies have revealed that administration of antibody against TIM-3 in proteolipid protein-induced experimental autoimmune encephalomyelitis (EAE) significantly exacerbates the clinicopathological severity of EAE, and increases the number and level of activation of macrophages (Wei et al., 2013; Xiao et al., 2013; Xu et al., 2013). These observations were subsequently extended to show that in vivo treatment with TIM-3-Ig fusion protein promotes a hyperproliferative response in CD4+ cells and increases IFN-γ production (Wei et al., 2013). Several similar studies complemented these findings, and they showed that in vivo blockade of TIM-3 using blocking monoclonal antibodies or fusion proteins exacerbates experimental allergic encephalomyelitis and autoimmune diabetes (Poonia and Pauza, 2014; Qiao et al., 2014).
It has been reported that single-nucleotide polymorphisms (SNPs) in TIM-3 gene may be associated with various diseases such as gastric cancer and rheumatoid arthritis (Xu et al., 2011, 2012). However, functions of these TIM-3 polymorphisms remain unknown. In the current study, we investigated associations between two TIM-3 polymorphisms (−574G/T and +4259T/G) and the risk of AS, and examined the effects of these SNP on gene expression in different immune cells.
Materials and Methods
Patients and controls
A total of 282 AS and 298 healthy controls were recruited. Patients were considered eligible if they had been diagnosed with AS using the modified New York Criteria. Patients with familial Mediterranean fever, psoriasis, psoriatic arthritis, inflammatory bowel disease, or a significant history of psychiatric disease; those younger than 18 years; and those who refused to participate in the study were excluded. Due to ethical and legal reasons, the psychiatric patients were excluded. Healthy controls were selected from hospital blood bank donors. Those who came to the blood bank to voluntarily donate blood and who agreed to participate in the study were recruited.
To exclude the possible effects of ethnicity, only Han Chinese were included in this study. Informed consent was obtained from all study participants according to the Helsinki Declaration. This study was approved by the institutional review boards of the Putuo Hospital and the Changzheng Hospital.
DNA extraction and genotyping
DNA extraction and genotyping were conducted using previously published methods (Cao et al., 2010; Cai et al., 2012; Tong et al., 2012; Bai et al., 2013; Song et al., 2013; Xu et al., 2011, 2012). In brief, the TIM-3 −574G/T and +4259T/G polymorphisms were detected by the polymerase chain reaction–restriction fragment length polymorphism (PCR–RFLP) assay. PCR was performed in a total reaction volume of 20 μL containing 2 μL of 10X PCR buffer (Qiagen, Inc., Hilden, Germany), 1.5 mM MgCl2, 0.5 μM of each primer, 0.2 mM dNTP, 1.2 U Taq polymerase (Qiagen, Inc., Hilden, Germany), and 200 ng of genomic DNA. After an initial denaturation at 95°C for 5 min, the DNA was amplified for 35 cycles at 94°C for 30 s, 55–60°C for 40 s, and 72°C for 45 s, with a final elongation at 72°C for 10 min on the Gene-Amp PCR System 9700 (PE Applied Biosystems, Foster City, CA). PCR products containing the three polymorphic sites were then digested with the restriction enzymes TaqI and PstI (New England Biolabs, Beverly, MA), respectively, by using the conditions recommended in the manufacturer's instructions. The digested PCR products were fractionated on 2% agarose Tris–borate–EDTA gel (Agarose 1000; Gibco BRL, Rockville, MD) and stained with ethidium bromide. To confirm the genotyping results, more than 10% of PCR-amplified DNA samples were examined by DNA sequencing. Results between PCR and DNA sequencing analysis were 100% concordant.
Real-time quantitative reverse transcriptase–PCR and western blot
Real-time quantitative reverse transcriptase–PCR (RT–PCR) was performed using the Exicycler 96 real-time RT–PCR system (A-2060; BIONEER, Daejeon, Korea) and SYBR Green mastermix (SY1020; Solarbio, Beijing, China). Amplification was performed under the following conditions: 95°C for 10 min, followed by 40 cycles of 95°C for 10 s, 60°C for 20 s, and 72°C for 30 s. Experiments were performed in triplicate in the same reaction. The results of the real-time quantitative RT–PCR experiments were calculated using the 2[−Delta Delta C(T)] method (Rong et al., 2014). Western blot was conducted according to a previously published method (Rong et al., 2014).
Statistical analysis
The SPSS statistical software package ver.13.0 (SPSS, Inc., Chicago, IL) and the Prism 5.0 were used for statistical analysis. Demographic data between the study groups were compared by the chi-square test and by the Student's t-test. Hardy–Weinberg equilibrium was analyzed using the chi-square test. For SNP analysis, genotype and allele frequencies of TIM-3 were compared between groups using the chi-square test, and odds ratios (OR) and 95% confidence intervals (CIs) were calculated using unconditional logistic regression. Student's t-test was used to compare mRNA and protein levels between different genotypes. p-values less than 0.05 were considered significant.
Results
Characteristics of the subjects
A total of 282 AS cases and 298 controls were recruited for the present study. All subjects were ethnic Chinese. Demographic and other selected characteristics of the cases and controls are presented (Table 1). Cases and controls did not show statistically significant differences with regard to age (p>0.05) and sex (p>0.05). Disease status was assessed using validated measures of disease activity (Bath AS Activity Index; BASDAI) and function (Bath AS Functional Index; BASFI). The mean scores were 3.56 and 3.62, respectively.
TIM-3 polymorphisms and susceptibility to AS
Genotype and allele frequencies of the TIM-3 −574G/T and +4259T/G polymorphisms in AS cases and controls are summarized in Table 2. The genotype distributions of these SNPs among the controls were in agreement with the Hardy–Weinberg equilibrium (p>0.05). Status of linkage disequilibrium between the two SNPs showed that they were not linked with each other (D′<0.01). As for the TIM-3 −574G/T SNP, prevalence of GT genotype was 13.8% in patients and 6.0% in controls (OR=2.50, 95%CI: 1.39–4.48, p=0.002). The −574T allele also revealed significantly increased frequencies in patients than in controls (OR=2.39, 95%CI: 1.39–4.22, p=0.002). As for the TIM-3 +4259T/G SNP, prevalence of TT genotype and TG genotype was 92.9% and 7.1% in patients and 93.6% and 6.4% in controls. No significant differences in the frequencies of +4259 genotypes or alleles were found between patients and controls (Table 2). These results indicate that TIM-3 −574G/T polymorphism is associated with increased susceptibility to AS in the Chinese population. We further analyzed haplotypes of the two polymorphisms between cases and controls (Table 2). Results presented a strong association between TT haplotype (−574T and +4259T) and increased risk of AS (p=0.002).
p-Value<0.05.
TIM-3 polymorphisms and gene expression
It has been reported that TIM-3 −574G/T and +4259T/G polymorphisms can be associated with various diseases, whereas our data also showed a correlation between −574G/T SNP and AS. However, functions of these polymorphisms have not been researched. We investigated effects of −574G/T and +4259T/G polymorphisms on TIM-3 gene expression by quantitative real-time RT–PCR and western blot. We first examined the effect of −574G/T polymorphism, and in order to avoid the potential interference from +4259T/G polymorphism, healthy donors carrying this genetic variant were excluded. A total of 20 patients and 15 controls carrying Tim-3 −574GG genotype, as well as 20 patients and 15 controls carrying Tim-3 −574GT genotype were included to assess the effect of Tim-3 −574G/T polymorphism on mRNA and protein levels of Tim-3. Data showed that mRNA level of TIM-3 in CD4+ T cells, CD8+ T cells, and monocytes was significantly decreased in subjects carrying −574GT genotype than in those with GG genotype (Fig. 1A), in which mRNA level of TIM-3 in CD4+ T cells revealed the most striking decrease by 60% than those in CD8+ T cells, and monocytes (Fig. 1A). Similarly, protein level of TIM-3 in CD4+ T cells, CD8+ T cells, and monocytes was significantly downregulated in subjects carrying the polymorphic −574GT genotype, in which CD4+ T cells presented the most decrease (Fig. 1B). Using the same method, we investigated the effect of +4259T/G on gene expression. A total of 16 patients and 14 controls carrying Tim-3 +4259TT genotype, as well as 16 patients and 14 controls carrying Tim-3 +4259GG genotype were included to assess the effect of Tim-3 +4259T/G polymorphism on mRNA and protein levels of Tim-3. However, no significant changes were found in CD4+ T cells, CD8+ T cells, or monocytes among TT and TG genotypes (Fig. 2). Student's t-test was used to compare mRNA and protein levels between different genotypes.
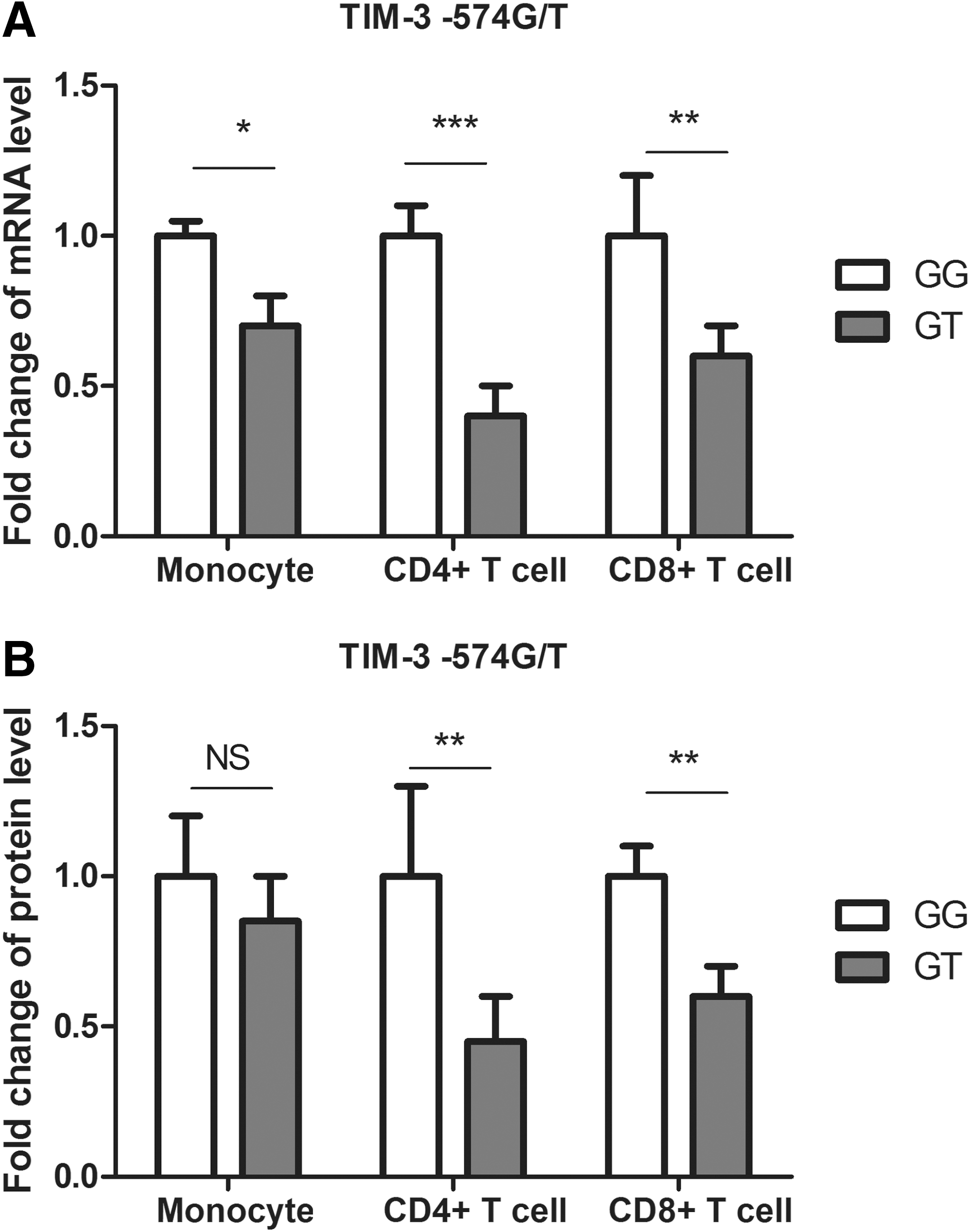
TIM-3 expression in monocytes, CD4+ T cells, and CD8+ T cells from healthy controls with different −574G/T genotypes.
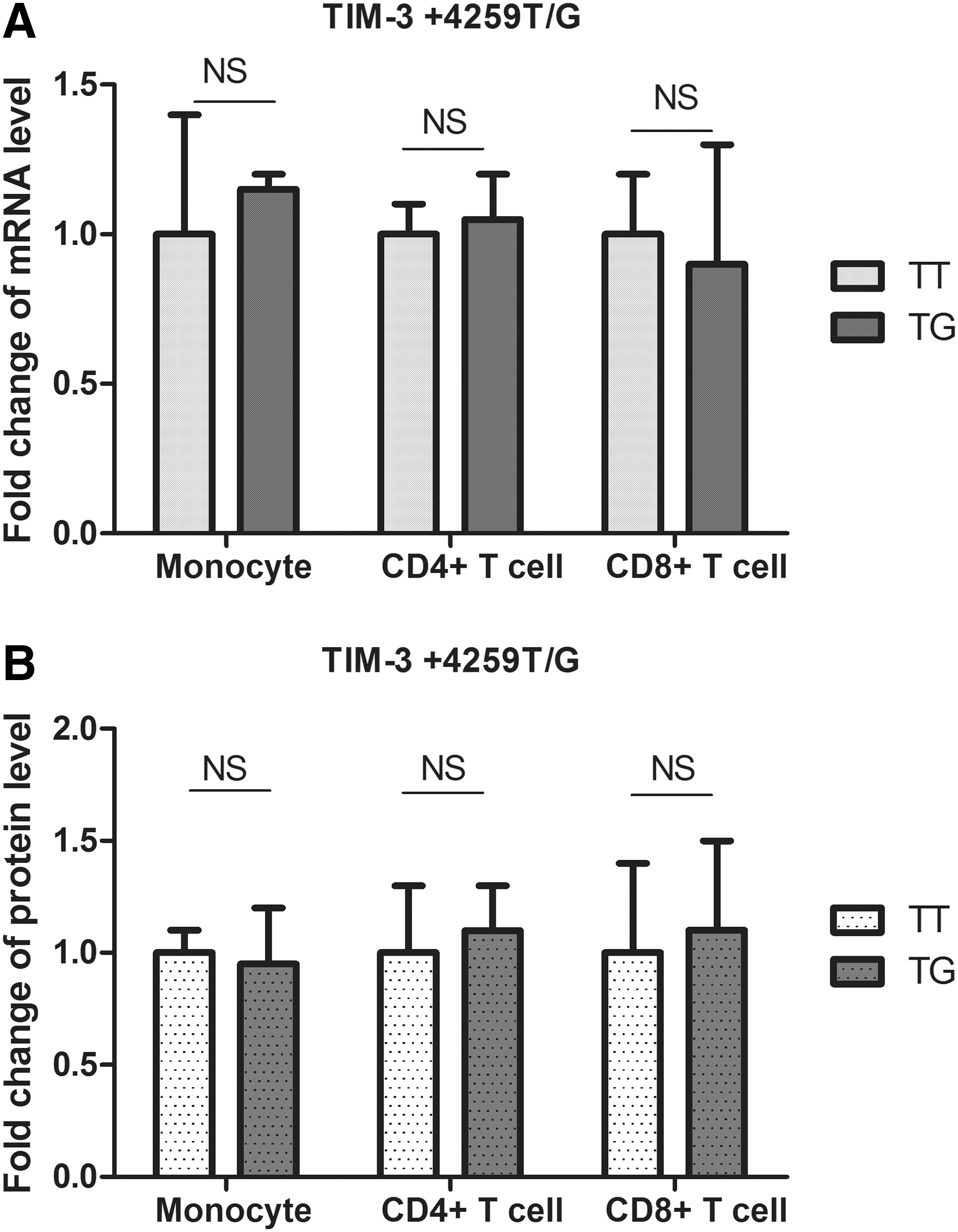
TIM-3 expression in monocytes, CD4+ T cells, and CD8+ T cells from healthy controls with different +4259T/G genotypes.
Discussion
In the current study, we identified that −574G/T SNP was associated with increased risk of AS. Furthermore, subjects carrying this polymorphism showed lower TIM-3 mRNA and protein levels in CD4+ T cells, CD8+ T cells, and monocytes, in which TIM-3 expression in CD4+ T cells revealed the largest decrease. These results suggested that the −574G/T SNP may affect the susceptibility to AS by interfering TIM-3 gene expression.
Polymorphism studies have suggested that TIM-3 −574G/T and +4259T/G SNPs are associated with increased risk of non-Hodgkin lymphomas (NHL), especially HIV-related NHL; +4259T/G SNP is associated with increased risk of nonsmall-cell lung cancer (NSCLC) and can be used as a prognostic marker for the disease; +4259TG genotype and +4259G allele are significantly increased in the pancreatic cancer cases than in controls; and −574G/T and +4259T/G polymorphisms were significantly increased in the renal cell carcinoma cases (Cao et al., 2010; Xu et al., 2011, 2012; Cai et al., 2012; Tong et al., 2012; Bai et al., 2013; Song et al., 2013). All the results indicate an important role of TIM-3 −574G/T and +4259T/G SNPs in the development of various diseases. Our study identified the correlation between −574G/T and AS. The −574TT and +4259GG genotypes were not found in our study population, which was consistent with other literature. In addition, the −574T allele was 3.0% and +4295G allele was 3.2% in the healthy controls, which were consistent with previous studies about the Han Chinese population.
The mechanism behind why −574G/T and +4259T/G polymorphisms are critical to different diseases remains unclear. The −574G/T SNP is located in the promoter region of TIM-3 gene. It is possible that the polymorphism may affect the gene expression by interfering TIM-3 promoter activity. Our data revealed that subjects carrying −574GT genotype had significantly lower TIM-3 mRNA and protein levels, suggesting that the polymorphism may affect TIM-3 expression from transcription level, probably by downregulating the promoter activity. In addition, we observed a different effects of this SNP on different immune cells (Fig. 1), suggesting that other factors may also be involved in the function of the polymorphism. Since TIM-3 plays inhibitory roles in T-cell responses, it is possible that −574G/T SNP downregulates TIM-3 expression, causes sustained activation of T-cell responses, and results in the increased risk of developing AS. The +4259T/G polymorphism is located in exon 3 of TIM-3 gene. The T to G switch causes an amino-acid substitution from arginine to leucine. Effects of this amino-acid change are yet unknown. According to our data, this SNP seemed not to affect TIM-3 mRNA or protein levels (Fig. 2). Since +4259T/G polymorphism is associated with various diseases, it should have some functional significance. It is possible that the amino-acid substitution caused by the SNP results in changes of TIM-3 structure and, therefore, affects the immune functions of TIM-3-expressing cells.
This case-control study demonstrates for the first time that the TIM-3 SNP is associated with increased risk of AS and downregulates TIM-3 mRNA and protein levels in monocytes, CD4+ T cells, and CD8+ T cells. Our results may show a new direction for further understanding the functions of TIM-3 gene.
Footnotes
Disclosure Statement
No competing financial interests exist.