
Abstract
Interferon gamma (IFN-γ) is one of the key cytokines that plays a major role against viral and intracellular bacterial infection. In addition to the IFN-γ gene, teleost fish possess a second copy known as IFN-γ-related (IFN-γrel) gene. This report describes structural and functional properties of IFN-γrel gene in the Indian major carp, rohu (Labeo rohita), a commercially important freshwater fish species in the Indian subcontinent. The rohu IFN-γrel gene consisted of four exons with three intervening introns and phylogenetically closely related to grass carp. The full-length IFN-γrel cDNA comprised 927 bp nucleotides with a single open reading frame of 504 bp, encoding 167 amino acids (aa) polypeptide with a signal peptide of 24 aa. The mature rohu IFN-γrel protein was 143 aa with a predicted molecular weight of 16.85 kDa. Basal expression analysis of IFN-γrel showed its wide range of expression in all examined tissues: The highest was in the skin and the lowest was in the liver. In response to LPS, poly I:C, iE-DAP, muramyl dipeptide stimulations, and bacterial infections, IFN-γrel gene expression was significantly (p<0.05) induced in treated fish tissues as compared with their control. The IFN-γrel was expressed as recombinant protein (rIFN-γrel) and confirmed through western blot. Stimulation of peripheral blood leukocytes with rIFN-γrel protein resulted in the activation of IFN-γ receptor and marked induction of inducible nitric oxide synthase gene expression. These results together may suggest the important role of IFN-γrel as an antimicrobial cytokine in fish.
Introduction
I
Fish IFN-γ was first reported in fugu (Zou et al., 2004) followed by Atlantic salmon (Robertsen, 2006), catfish (Milev-Milovanovic et al., 2006), goldfish (Grayfer and Belosevic, 2009), zebrafish (Igawa et al., 2006), pufferfish (Igawa et al., 2006), common carp (Stolte et al., 2008), Atlantic cod (Furnes et al., 2009), grass carp (Chen et al., 2010), and rohu (Parhi et al., 2014). The fish IFN-γ showed a low sequence similarity (17–31%) with that of birds and mammals and formed a separate cluster in the phylogenetic tree.
The biological effects of IFN-γ are mediated by formation of receptor complex of IFN-γ receptor 1 and 2, resulting in activation of downstream signaling molecule JAK1 and JAK2, and subsequent phosphorylation of STAT1. The activated STAT1 homodimer translocates to the nucleus, where it activates the expression of many IFN-γ-stimulated genes (Walter et al., 1995). IFN-γ can activate the nitric oxide generation, involved in antimicrobial and cytotoxic defense. This chemical mediator is produced by nitric oxide synthase (NOS), which converts
Rohu (Labeo rohita), an economically important freshwater fish species in the Indian subcontinent, succumbed to various diseases during its commercial farming (Karunasagar et al., 1989). Therefore, development of various preventive measures and treatments against diseases were essential for its profitable farming. Hypothesizing the important function of IFN-γrel as innate immune cytokine in rohu, this gene was cloned and sequenced and its expression in healthy and bacteria-infected fish tissues was investigated. The function of rohu IFN-γrel was also investigated by expressing it as recombinant IFN-γrel protein. Stimulation of peripheral blood leukocytes (PBLs) with recombinant IFN-γrel resulted in activation of IFN-γ receptor and iNOS gene expression, suggesting IFN-γrel as an anti-microbial cytokine.
Materials and Methods
Fish
Rohu (L. rohita) fingerlings (average weight ∼50 g) were obtained from the Central Institute of Freshwater Aquaculture (CIFA), and they were stocked in 500 L aerated tanks (water temperature 25–28°C; pH 7.4–7.6) with each tank containing 50 fish. Before the start of the experiment, acclimatization was carried out for 3 weeks and the fish were fed twice a day with a commercial carp diet with daily two-third water exchange. Before sacrifice, fishes were anesthetized with tricane methane sulphonate (Himedia).
Bacteria
Aeromonas hydrophila (ATCC-35654), Shigella flexneri (ATCC-9199), and Edwardsiella tarda (ATCC-15947) were cultured separately in LB broth (USB) at 37°C for 16 h with constant shaking. Viable count was determined as a colony-forming unit (CFU) after 10-fold serial dilutions and plating on nutrient agar.
Cloning of rohu IFN-γrel
To clone rohu IFN-γrel gene, total RNA was extracted from rohu gill, and cDNA was prepared following the protocol as described later. To amplify IFN-γrel gene through polymerase chain reaction (PCR), degenerated primers IFN-γrel (A) FW and IFN-γrel (A) RV (Table 1A) were designed based on the nucleotide sequences of the conserved regions of common carp (AM261214.1), grass carp (FJ695519.1), and zebrafish (AB194272.1) IFN-γrel gene. PCR was carried out in a 50 μL reaction volume, utilizing 1 μL cDNA as template under the following conditions: one cycle of initial denaturation at 94°C for 2 min, 45 cycles of 94°C for 30 s, 58°C for 30 s, 72°C for 1 min, and a final extension at 72°C for 10 min. In a 2% agarose gel, 15 μL of the PCR product was analyzed, and the single specific band was purified with agarose gel purification kit (Roche). The eluted DNA was cloned in pGEM-T Easy vector (Promega), and both strand DNA sequencing was carried out with T7 and SP6 primer following Sanger sequencing protocol in a 96-capillary high-throughput sequencer, ABI 3730 XL (Xcelris Labs Ltd.). The obtained sequence was confirmed by BLAST search (Altschul et al., 1990). To obtain full-length IFN-γrel cDNA sequence, 5′ and 3′ RACE (rapid amplification of cDNA ends) was carried out using the SMARTer RACE cDNA amplification kit (Clontech). The rohu IFN-γrel gene specific forward (IFN-γrel-GSP-FW) and reverse (IFN-γrel-GSP-RV) primers were designed from the obtained partial IFN-γrel cDNA sequence. Touchdown-PCR was carried out for 3′ and 5′ RACE using IFN-γrel-GSP-FW/UPM and IFN-γrel-GSP-RV/UPM primer sets (Table 1A), respectively, under the following conditions: 1 cycle of initial denaturation at 94°C for 2 min, followed by 5 cycles of 94°C/30 s, 72°C/2 min; next 5 cycles 94°C/30 s, 70°C/30 s, 72°C/2 min; next, 27 cycles of 94°C/30 s, 60°C/30 s, 72°C/2 min; and 1 cycle at 72°C/5 min. Nested PCR for 3′and 5′ RACE were performed using IFN-γrel-GSP-FW/NUP for 3′ and IFN-γrel GSP-RV/NUP for 5′ nested primer sets (Table 1A). For nested PCR, 1 μL of primary RACE-PCR product was used as a template with the following conditions: initial 1 cycle of 94°C/2 min, then 30 cycles of 94°C/30 s, 60°C/30 s and 72°C/2 min followed by 1 cycle of 72°C/10 min. The PCR products were cloned in pGEM-T Easy vector, sequenced, and validated through BLAST search and overlapping sequences from the fragments were combined together to get the full-length cDNA. The obtained cDNA sequence was submitted to the GenBank.
Restriction sites are marked as bold and underlined.
To identify the introns and exons, PCR primers IFN-γrel (B) FW and IFN-γrel (B) RV (Table 1A) were designed from the obtained IFN-γrel cDNA sequence. PCR was carried out using rohu genomic DNA as a template with following PCR conditions: initial 1 cycle of 94°C/2 min, 40 cycles of 94°C/30 s, 52°C/30 s, and 72°C/3 min followed by 1 cycle of 72°C/10 min. Amplified product was gel purified, cloned in pGEM-T Easy vector and DNA sequencing was carried out with both forward and reverse primers. The obtained IFN-γrel genomic DNA sequence was analyzed by aligning with the full-length cDNA sequence and was submitted to the GenBank.
Sequence analysis
Amino acids in rohu IFN-γrel were predicted from full-length cDNA sequence by Lasergene DNA star software and were aligned with other species IFNs by CLUSTALW. The alignment file generated by CLUSTALW was submitted in ESPript2.2 program (
Tissue-specific expression of IFN-γrel gene
To examine basal expression of IFN-γrel gene, various organs, namely, skin, blood, muscle, gill, heart, spleen, intestine, brain, eye, kidney, and liver, were collected separately from three rohu fingerlings in TRIzol reagent. Total RNA was isolated from each sample and cDNA was prepared following the protocol as described later. The quantitative real-time PCR (qRT-PCR) was carried out to analyze IFN-γrel gene expression, keeping β-actin as an internal control.
Treatment of fish with microbial products and infection with bacteria
Healthy rohu fingerlings were divided into control and treated groups while keeping three fish in each group. For ligand exposure, purified LPS of Escherichia coli (serotype O55:B5) (Sigma) was reconstituted in endotoxin-free water at 5 mg/mL, and 100 μL of endotoxin-free water containing 20 μg of diluted LPS was injected into the fish (treated group) through intra-venous (i.v.) route. The control fish group was i.v. injected with 100 μL of endotoxin-free water. For poly I:C-treated fish group, purified poly I:C (Sigma) was diluted in DEPC-treated water at 10 mg/mL, 100 μL of DEPC-treated water containing 300 μg of diluted poly I:C was i.v. injected, and control fish received only 100 μL of DEPC-treated water through i.v. route. Muramyl dipeptide (MDP) (Sigma) was diluted in endotoxin-free water at 1 mg/mL, 100 μL of endotoxin-free water containing 50 μg MDP was i.v. injected, and control fish were injected with 100 μL of endotoxin-free water.
Expression of recombinant rohu IFN-γrel and its confirmation in western blotting
The rohu IFN-γrel cDNA fragment encoding the full-length matured peptide was amplified by using IFN-γrel (C) FW and IFN-γrel (C) RV primer pair (Table 1A). The forward and reverse primers were tagged with “BamHI” and “PstI” restriction sites, (marked as bold and underlined in Table 1A) respectively. The PCR product was analyzed on a 1.5% agarose gel, and the single specific band was purified with agarose gel purification kit (Roche). The purified PCR product was digested with restriction enzymes BamHI and PstI, ligated into the BamHI and PstI restriction enzyme sites of bacterial expression vector pQE30Xa (Qiagen) that consisted of six Histidine tag at the N-terminus for purification of the recombinant protein. The resultant plasmid pQE30Xa-rIFN-γrel was transformed into E. coli competent cells M15, and positive clones were selected by colony PCR. Plasmid DNA was purified by using High Pure Plasmid Isolation Kit (Roche) and sequenced for confirmation.
Single positive colony was cultured in LB medium containing 100 μg/mL ampicillin and 25 μg/mL kanamycin at 37°C. The expression of IFN-γrel was induced with various concentrations of IPTG for 4 h, and among them, 1 mM IPTG was selected as a standard inducer. After 4 h of induction with 1 mM IPTG, bacteria were harvested by centrifugation at 5000×g for 10 min and lysed by sonication (40 amp 10 s pulse). The recombinant IFN-γrel protein was expressed as inclusion bodies and remained in the insoluble fraction. Therefore, to confirm the expression of recombinant IFN-γrel, it was purified from the bacterial extracts under denaturing condition using the Ni-NTA resin column following the user manual of QIAexpress Ni-NTA Fast Start Kit (Qiagen). Purified recombinant IFN-γrel (rIFN-γrel) was analyzed in 15% SDS-PAGE followed by western blotting with penta-his antibody (mouse monoclonal IgG1) (Qiagen) as the primary antibody and Amersham ECL Plus detection system (GE Healthcare). To renature the column-purified rIFN-γrel, it was first diluted in 9×higher volume of refolding buffer (50 mM Tris–HCl, pH 8.0; 2 mM oxidized glutathione; 0.22 mM reduced glutathione; 0.5 M
Isolation of PBLs and stimulation with recombinant rohu IFN-γrel
Leukocytes were isolated from the peripheral blood of rohu fingerlings by the density gradient centrifugation using histopaque-1077 (Sigma-Aldrich). Briefly, 1.5 mL of caudal venous blood was collected in an EDTA containing tube and mixed with PBS at a ratio of 1:1. The suspension was layered over an equal volume of histopaque-1077 (Sigma-Aldrich) gradient, and it was then centrifuged at 2500 g for 30 min at 18°C. After centrifugation, the interface and band were collected in a new tube, washed twice in PBS, and re-suspended in RPMI-1640 medium (Sigma-Aldrich). Cells were confirmed as PBLs under microscopic observation, and the cell viability was determined by trypan blue staining.
In a 12-well cell culture plate (TPP), purified PBLs in RPMI-1640 medium were distributed at 1 mL per well (∼106 cells). Cells in three wells were kept untreated and marked as control. In the treated group, cells in three wells were stimulated with 500 ng of refolded and purified rIFN-γrel protein. After 4 h, PBLs were separately collected from each well, and total RNA was extracted with TRIzol reagent. The cDNA was prepared and qRT-PCR was carried out to analyze the IFN-γ receptor and iNOS gene expression in control and treated cells after normalizing with β-actin gene expression.
RNA isolation and first-strand cDNA synthesis
Total RNA was extracted from the TRIzol-treated samples, namely, skin, blood, muscle, gill, heart, spleen, intestine, brain, eye, kidney, and liver, following the standard protocol (Invitrogen). The concentration of total RNA was measured by a UV-spectrophotometer (Eppendorf), and the integrity of the RNA was assessed by observing the band intensity of 28 and 18S ribosomal RNA in 1% agarose gel. To prepare first-strand cDNA, 1 μg of total RNA was treated with 1 U of DNase I (MBI) and reverse transcription was carried out using oligo-dT primer and RevertAid first-strand cDNA synthesis kit (MBI). The confirmation of cDNA synthesis was carried out with PCR amplification of β-actin gene, and the synthesized cDNA was kept at −80°C till further analysis.
Real-time PCR analysis
Quantitative real-time PCR (qRT-PCR) of the target genes (IFN-γrel, IFN-γ receptor, iNOS) and the reference gene (β-actin) was carried out in a 10 μL reaction volume, containing 1.0 μL of cDNA, 0.25 μL of FW and RV primers (2.5 μM each; Table 1B), 5 μL of 2×LightCycler®480 SYBR Green I master mix (Roche), and 3.5 μL of PCR grade H2O in LightCycler480 II real-time PCR detection system (Roche). The qRT-PCR was performed in triplicate wells under following conditions: initial denaturation at 95°C for 10 min followed by 45 cycles of 94°C/10 s, 53°C (for IFN-γrel, IFN-γ receptor, iNOS), or 55°C (for β-actin) for 10 s and 72°C/10 s. Negative control reaction was carried out without the template (cDNA). The PCR efficiencies were determined by analyzing the serial dilutions of cDNA, and efficiencies were almost 100%, allowing the use of 2−ΔΔCT method (Livak and Schmittgen, 2001) for calculation of relative gene expression of the target genes with that of reference gene, β-actin. The correct size and single band amplification was determined by analyzing 8 μL of the real-time PCR products in ethidium bromide-stained 2% agarose gel. The relative expression ratios were obtained by normalizing expression of the target gene, as determined by mean crossing point (cp) deviation by that of a nonregulated reference gene, β-actin following 2−ΔΔCT method. The data obtained from qRT-PCR analysis were expressed as mean of three individual experiments±standard error (s.e.), and the significant difference between the control and treated groups at each time point was determined by the Student's t-test using Microsoft Excel 2010 with p<0.05 as significance level.
Results
Cloning and characterization of rohu IFN-γrel gene
The rohu IFN-γrel full-length cDNA comprised 87 bp 5′ un-translated region (UTR), a 504 bp open reading frame (ORF), and 336 bp of 3′ UTR (with poly A tail). The ORF encoded 167 amino acid (aa) protein with a 24-aa signal peptide was expected to be cleaved between Ala24 and Phe25. The estimated molecular mass of the mature rohu IFN-γrel protein was predicted to be 16.85 kDa. The 3′ UTR consisted of three mRNA instability motifs (ATTTA) and was expected to be responsible for rapid mRNA degradation. A putative poly-adenylation signal (AATAAA) was identified at the 13 bp upstream of the poly (A) tail. Using full-length IFN-γrel ORF primers, a 950 bp PCR product was obtained from rohu genomic DNA. The alignment of IFN-γrel cDNA sequence with the genomic DNA sequence revealed four exons and three introns in rohu IFN-γrel gene (Fig. 1A, B). The first exon was 147 bp nucleotides in length; exons 2, 3, and 4 were 63, 210, and 84 bp, respectively. Three introns were 252, 89, and 105 bp, respectively. A typical intron splice motif (5′ GT-intron-AG 3′) was present at the 5′ end and the 3′ end of all the three introns (Fig. 1A). Fish IFN-γ and IFN-γrel sequences were aligned by ClustalW program (Fig. 2), and result revealed that there were four conserved amino acids, leucine, tyrosine, and two alanine, across the fish IFN-γ and IFN-γrel sequence. Furthermore two conserved cysteine residues were identified in fish IFN-γrel but not in IFN-γ molecules. The IFN-γ signature motif ([I/V]-QX-[KQ]-A-X2-E-[L/F]-X2-[I/V]) was present toward the C-terminal end of the sequence (Fig. 2). The predicted rohu IFN-γrel secondary structure consisted of four helixes. A putative glycosylation site was identified between helix D and E loop. The IFN-γ signature motif and two conserved alanine residues remained in the helix E region. Another two conserved amino acids leucine and tyrosine were present in helix A and D, respectively (Supplementary Fig. S1; Supplementary Data are available online at
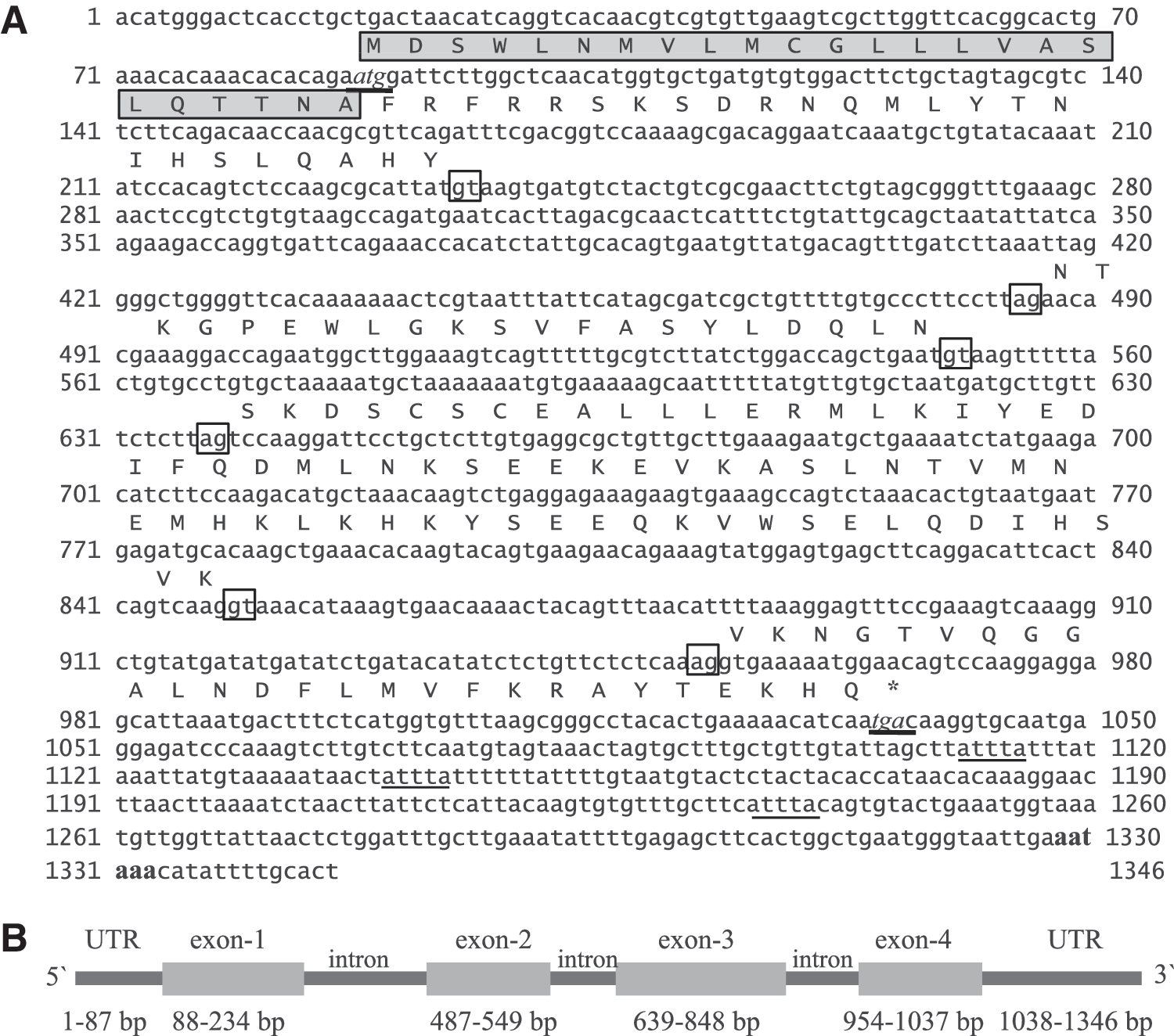
The nucleotide and deduced amino-acid sequence of rohu interferon gamma-related (IFN-γrel) gene.

Multiple alignment of fish IFN-γrel and IFN-γ amino-acid sequences. Amino-acid sequences were retrieved from the GenBank data base, aligned with CLUSTALW and block was prepared using ESPript2.2 program. The black background boxes represented strictly conserved regions, and other boxes represented well-conserved (>50%) regions. IFN-γ signature motif and two conserved cysteine residues in fish IFN-γrel genes were boxed, and putative glycosylation site was underlined.
Phylogenetic relationship of IFN-γrel
The percentages of similarity and identity of rohu IFN-γrel amino acids with other species were predicted in MatGAT, and they showed the highest similarity (83.8%) and identity (67.6%) with grass carp IFN-γrel. Within the mammalian IFN-γ, human IFN-γ showed the highest similarity (46.1%), and cow IFN-γ showed the highest identity (21.9%) with rohu IFN-γrel (Table 2).
To understand the evolutionary relationship, a phylogenetic tree was constructed with full-length IFN proteins of various species, and it revealed five major clusters, mammalian IFN-γ, avian-IFN-γ, fish IFN-γ, fish IFN-γrel, and out-group type I IFN supported by high bootstrapping values. Within the fish IFN-γrel cluster, rohu IFN-γrel was closer to the grass carp IFN-γrel (Fig. 3).
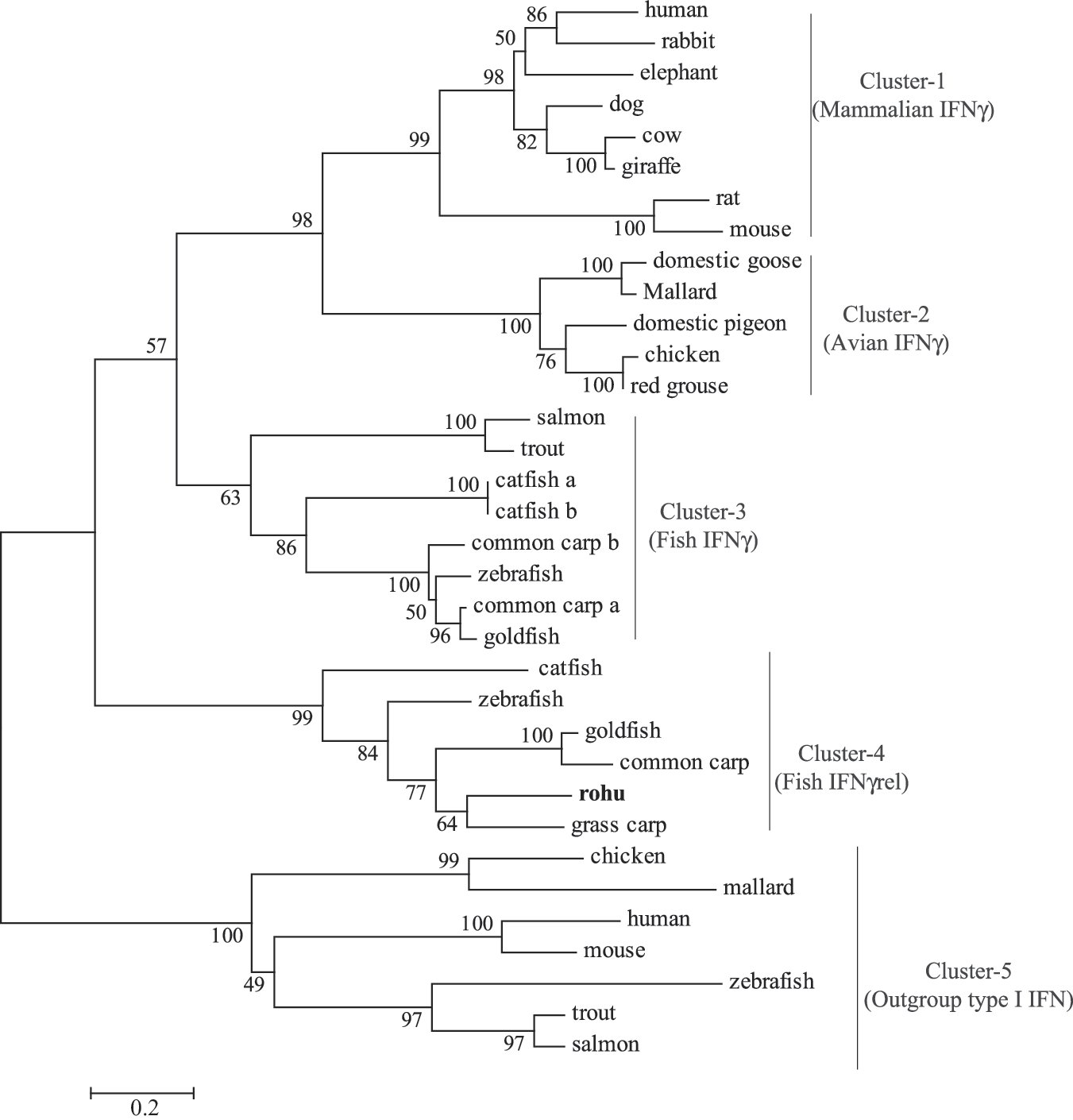
Phylogenetic relationship of rohu IFN-γrel with other species. IFN-γrel amino-acid sequences were aligned by CLUSTALW program within DNASTAR, and the unrooted phylogenetic tree was generated by the neighbor-joining method within the MEGA4 program. The tree was bootstrapped 1000 times, and the percentages were shown. The IFN amino-acid sequences used for tree construction were retrieved from the following GenBank database: human: P01579, NP_076918; rabbit: BAA24439; elephant: ABS28997; dog: P42161.2; cow: P07353; giraffe: ABS28996; rat: P01581; mouse: ABN80441, NP_034632; domestic goose: AAS57786; mallard: AAO13016, ABK55353; domestic pigeon: ABF18829; chicken: ABI83735, ABB05334; red grouse: ABF14465; salmon: ACN37863, AAP51035; trout: ACJ03571, AAV39394; catfish: AAZ40504, AAZ40505, AAZ40506; common carp: CAJ98867, CAJ51089, CAJ 51088; zebrafish: BAD06253, BAD72865, CAD67754; goldfish: ACG68885, ACV41807; and grass carp: ACN56578.
Tissue-specific expression of IFN-γrel
Expression of IFN-γrel in various organs/tissues of healthy rohu fingerlings was analyzed by qRT-PCR, and their mean values were presented graphically in Figure 4. Among the examined tissues, IFN-γrel gene expression was at detectable levels in all organs/tissues, but the level of expression varied among them. Lowest expression of IFN-γrel was observed in the liver (calibrator tissue; 1-fold), and the highest expression was detected in the skin (7.4-fold), followed by blood (∼5.7-fold), muscle (∼5.3-fold), gill (∼4-fold), heart (∼4-fold), spleen (∼3.5-fold), intestine (2.7-fold), and brain (2.2-fold). In the eye and kidney of rohu fingerlings, basal expression of IFN-γrel gene was ∼1.5-fold.
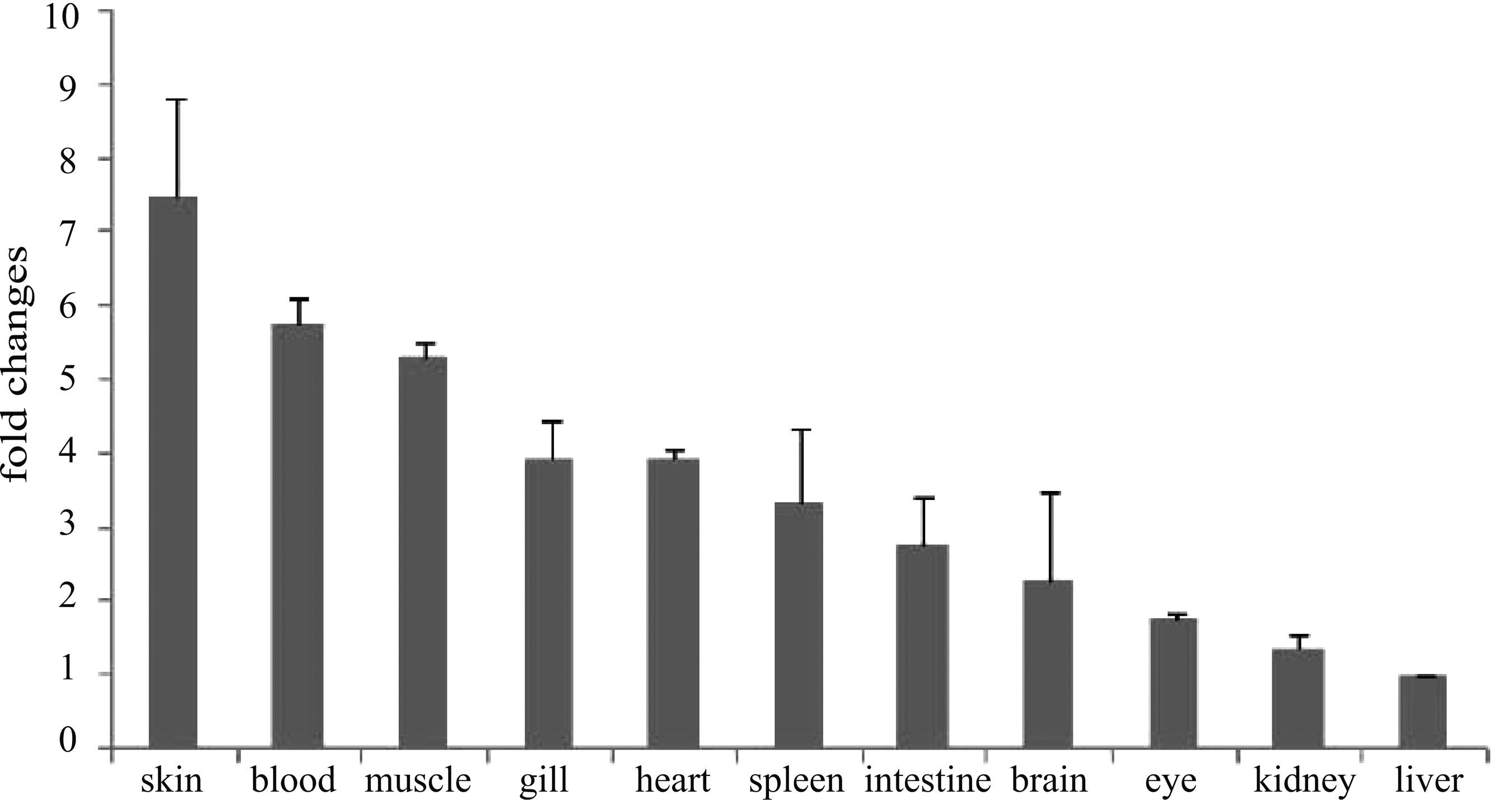
Basal expression of IFN-γrel gene in various tissues. Total RNA was extracted from skin, blood, muscle, gill, heart, spleen, intestine, brain, eye, kidney and liver, and it was analyzed by quantitative real-time polymerase chain reaction (qRT-PCR) assay to examine the expression of IFN-γrel gene among these tissues. Expression of IFN-γrel transcripts was represented as a ratio relative to β-actin (internal control) levels in the same samples. Liver was chosen as a calibrator (1), and the relative expression of IFN-γrel in other tissues was represented as fold changes from the calibrator. The results were expressed as mean±standard error (bars in the graph) from three fish (n=3).
Expression of IFN-γrel gene in response to pathogen-derived products
The LPS, iE-DAP, and MDP are the major constituents of various bacteria. To investigate the response of IFN-γrel gene, rohu fingerlings were i.v. injected with these pathogen-derived products, and expression of IFN-γrel gene was analyzed by qRT-PCR assay (Fig. 5). In response to LPS treatment, IFN-γrel was significantly (p<0.05) induced in gill (∼3-fold), liver (∼20-fold), kidney (∼37.5-fold), intestine (∼27.5-fold), and blood (∼40-fold) as compared with control. In response to iE-DAP, in all tested tissues, except gill and liver, significant (p<0.05) induction of IFN-γrel gene expression was observed as compared with their control. Among the tissues, the highest induction was observed in blood (5.1-fold), followed by kidney (2.7-fold) and intestine (2.6-fold). Similar pattern of IFN-γrel gene expression was also observed in MDP treatment. Among the tested tissues, the highest induction of IFN-γrel was observed in blood (∼11.7-fold), followed by kidney (∼6.5-fold), intestine (6.4-fold), and liver (1.6-fold).
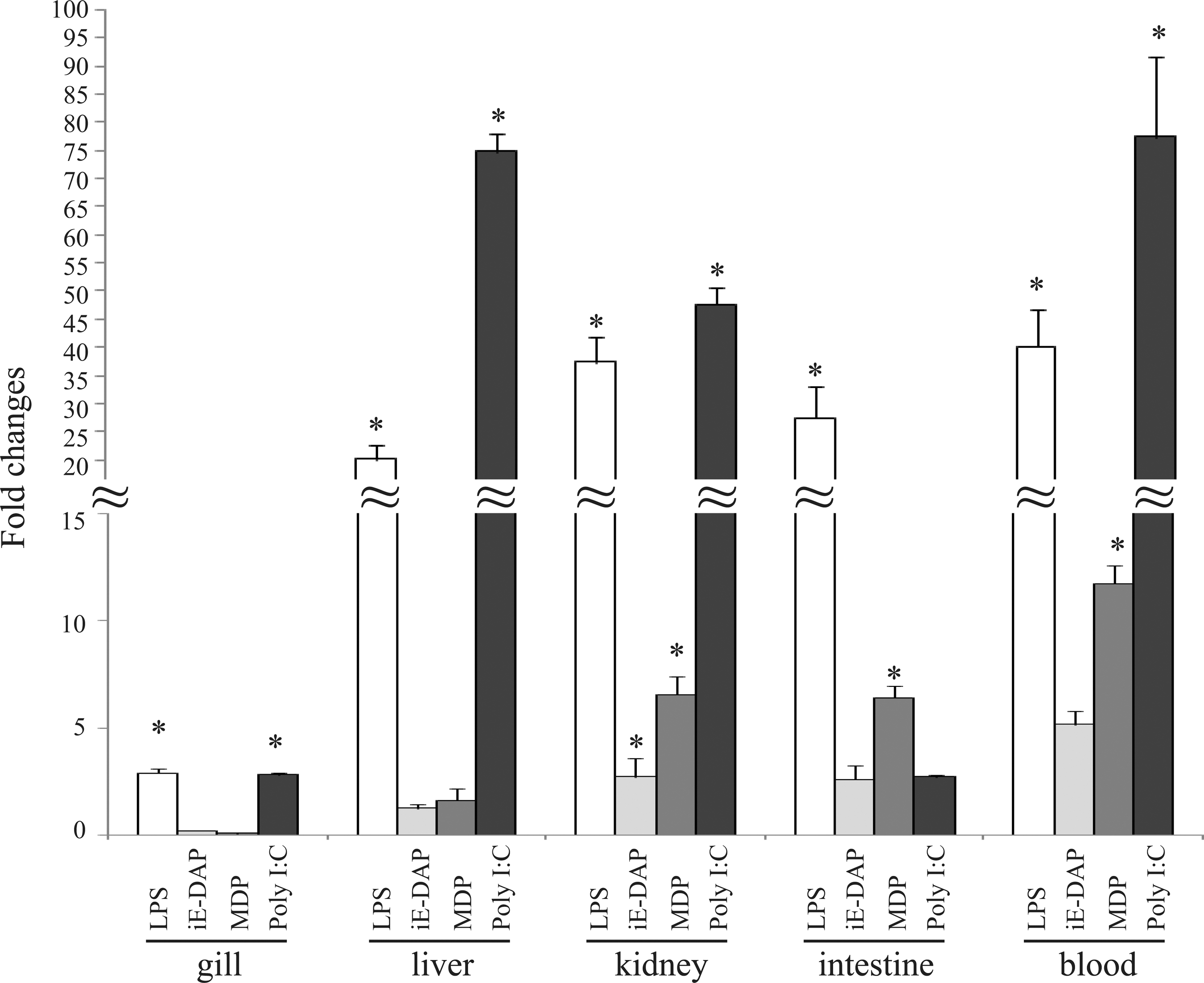
Modulation of IFN-γrel expressions by LPS, iE-DAP, muramyl dipeptide (MDP), and poly I:C treatment. In rohu fingerlings, LPS (20 μg/fish), iE-DAP (200 μg/fish), MDP (50 μg/fish), and poly I:C (300 μg/fish) were i.v. injected. After 4 h, total RNA of each tissue was analyzed with real-time PCR. The relative expression of IFN-γrel gene was normalized to the expression of β-actin (internal control) and expressed as fold changes relative to the untreated control group. The mean value of three fish (n=3) was shown, and the bars indicated standard error. Significant difference (p<0.05) of IFN-γrel gene expression between control and treated fish group was indicated with asterisks (*).
Poly I:C is a synthetic double-stranded RNA (dsRNA) that mimics viral dsRNA signature. To investigate the response of IFN-γrel gene during viral infection, poly I:C was i.v. injected and expression of IFN-γrel was assessed by real-time PCR. As shown in Figure 5, IFN-γrel was significantly induced in gill (∼2.8-fold), liver (∼74.8-fold), kidney (∼47.7-fold), and blood (∼77.5) as compared with their respective control.
Differential expression of IFN-γrel after bacterial infection
To investigate whether IFN-γrel expression could be modulated in bacterial infection, rohu fingerlings were infected with A. hydrophila, E. tarda, and S. flexneri, and IFN-γrel gene expression in gill, liver, kidney, intestine, and blood was examined by qRT-PCR assay at various time points (Fig. 6). As shown in Figure 6A, except in gill, IFN-γrel gene transcripts were significantly enhanced in all tested organs of A. hydrophila-infected fish as compared with control fish. The highest induction of IFN-γrel was noted in blood (∼25-fold) and intestine (∼15-fold) at 12 h, and in liver and kidney (∼17-fold) at 24 h postinfection.
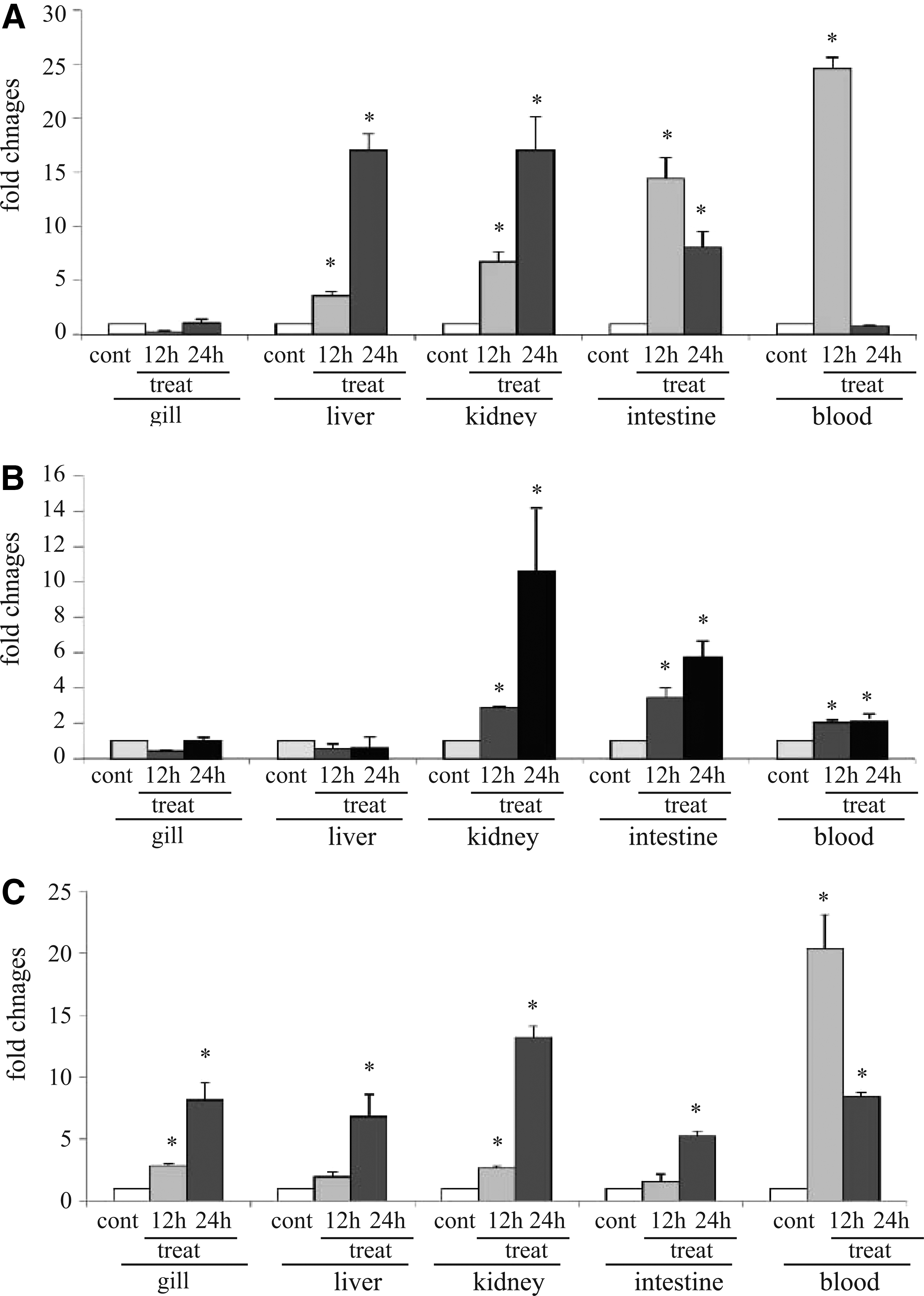
Modulation of IFN-γrel gene expression after bacterial infection. Aeromonas hydrophila (1×106 CFU/fish), Edwardsiella tarda (3×106 CFU/fish), or Shigella flexneri (5×106 CFU/fish) were injected separately into rohu fingerlings by intra-peritoneal (i.p.) route. After the designated time course, total RNA was extracted from gill, liver, kidney, intestine, and blood. Real-time PCR was conducted to analyze IFN-γrel and β-actin expression in various organs/tissues. The results were expressed as mean±standard errors (bars) from three fish (n=3) after normalizing the values with β-actin (internal control). Significant difference (p<0.05) between the control and treated group was indicated with asterisks (*).
In response to E. tarda infection, a steady induction of IFN-γrel was observed in the kidney, intestine, and blood of the infected fish. In the infected fish kidney, the IFN-γrel induction was ∼3 fold at 12 h, and at 24 h it reached ∼11 fold. In the intestine and blood, the induction was ∼4 and ∼2 fold at 12 h, and at 24 h, it reached ∼6 and 2.2 fold, respectively. In contrast to these tissues, there was not much variation of IFN-γrel expression in gill and liver between the infected and control fish (Fig. 6B). Among the organs, kidney was the most responsive followed by the intestine and blood.
Next, we analyzed the expression of IFN-γrel in S. flexneri-infected and control fish by qRT-PCR. The data of qRT-PCR revealed significant (p<0.05) upregulation of IFN-γ in all treated tissues as compared with control (Fig. 6C). Time course (12 and 24 h) analysis showed the progressive induction of IFN-γrel in gill, liver, kidney, and intestine and at 24 h post infection, the intensity of IFN-γrel induction in these tissues reached ∼8-, 7-, 13.2-, and 5.2-fold, respectively. In the blood, the peak induction (∼20-fold) of IFN-γrel was at 12 h p.i., and with the advancement of time it declined to ∼8.2 fold at 24 h p.i.
Recombinant rohu IFN-γrel activates IFN-γ receptors and induces iNOS gene expression
To investigate the efficacy of rohu IFN-γrel in inducing IFN-γ receptor and in nitric oxide generation, recombinant IFN-γrel was expressed in E. coli. The expression of rIFN-γrel in response to various concentrations of IPTG stimulation was confirmed by SDS-PAGE analysis (Fig. 7A). The rIFN-γrel protein was purified from the bacterial cell lysates through affinity chromatography, and it was confirmed by western blotting showing a single specific band of ∼23 kDa (Fig. 7B). Stimulation of PBLs with refolded, and purified rIFN-γrel protein significantly induced IFN-γ receptor (Fig. 8A) and iNOS (Fig. 8B) gene expression.
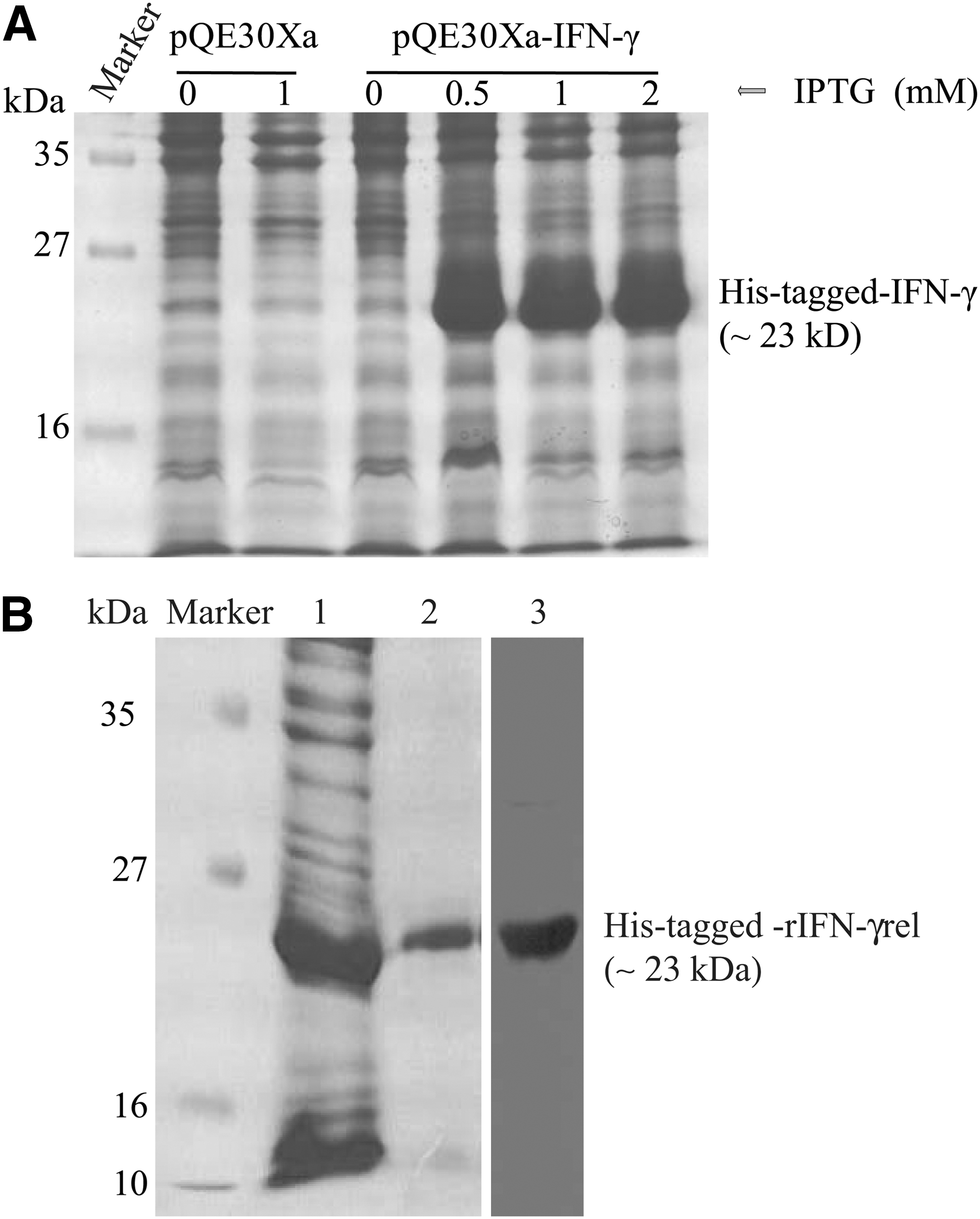
Expression and purification of recombinant rohu IFN-γrel protein.
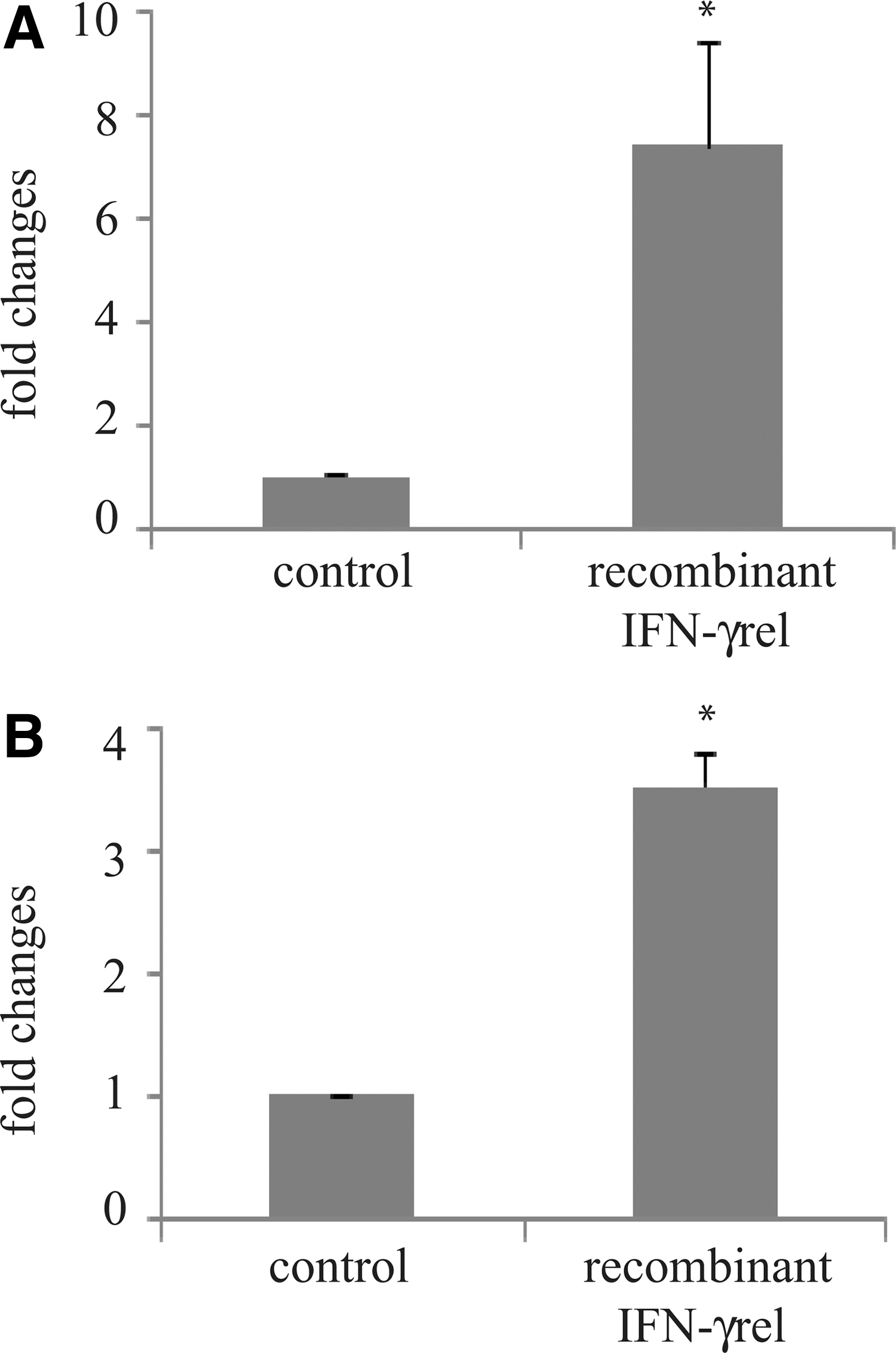
Recombinant rohu IFN-γrel induced IFN-γ receptors and iNOS gene expression. Peripheral blood leukocytes (PBLs) were isolated from blood and were distributed at 1 mL per well (∼106 cells) in RPMI-1640 medium. Cells in three wells were kept untreated and marked as control. In the treated group, cells in three wells were stimulated with 500 ng of refolded and purified rIFN-γrel protein for 4 h. After the designated time course, total RNA was extracted from the cells, and cDNA was analyzed with quantitative real-time PCR assay. The relative expression of IFNγ receptor and iNOS genes was normalized to the expression of β-actin (internal control) and expressed as fold changes relative to the untreated control cells. The mean of three wells (n=3) was shown, and the bars indicated error. Significant difference (p<0.05) between control and treated group is indicated with asterisk (*).
Discussion
IFN-γ is a pro-inflammatory cytokine, and it is known to be expressed after bacterial and viral infections (Dussurget et al., 2014). In addition to the IFN-γ gene, teleost fish possess a second copy known as IFN-γrel gene. Unlike IFN-γ, most of the functions of IFN-γrel gene remained unknown in various fish species (Aggad et al., 2010).
In India, rohu is one of the most commercially important freshwater fish species, and this report describes the molecular characterization of IFN-γrel gene in this fish species. The rohu IFN-γrel gene consisted of four exons and three introns, and it was structurally similar to other fish species. The presence of signal peptide in rohu IFN-γrel protein strongly suggested it as a secretory cytokine. Phylogenetically, rohu, and grass carp IFN-γrel formed a separate cluster and this relationship was also supported by the maximum similarity (83.8%) and identity (67.6%) in their amino-acid sequences. The similarity and identity of the amino acids also likely suggest the functional similarities of IFN-γrel protein in rohu and grass carp.
In rohu, IFN-γrel was constitutively expressed in all tested organs and the expression was the highest in the skin and the lowest in the liver, and markedly different from zebrafish (Igawa et al., 2006), catfish (Milev-Milovanovic et al., 2006), and common carp (Stolte et al., 2008).
LPS and iE-DAP of Gram-negative bacteria and MDP of Gram-positive bacteria are major structural constituents. In rohu, induction of IFN-γrel gene expression was significantly induced by LPS, iE-DAP, and MDP, hypothesizing the critical role of IFN-γrel during bacterial infections. To confirm this, rohu fingerlings were challenged with A. hydrophila, E. tarda, and S. flexneri infections, and as expected, significant upregulation of IFN-γrel gene expression was noted in the infected fish tissues. Among these bacterial infections, most significant expression of IFN-γrel gene was observed in A. hydrophila infection followed by S. flexneri and E. tarda. The variation of IFN-γrel gene expression pattern by these three different bacteria may be due to their different mode of infection.
The polyinosinic-polycytidylic acid (poly I:C) is a synthetic dsRNA that mimics the genome of viral hemorrhagic septicemia virus and grass carp reovirus (Su et al., 2009; Yang and Su, 2010). In response to poly I:C, IFN-γrel expression was also induced in rohu fingerlings. Induction of IFN-γrel expression after poly I:C and LPS treatment was previously reported in common carp (Milev-Milovanovic et al., 2006), grass carp (Chen et al., 2010), and zebrafish (Igawa et al., 2006). In addition to other fish species, these observations in rohu strongly suggested the critical role of IFN-γrel in fish during bacterial and viral infections.
The mechanism of inductive expression of IFN-γrel gene in rohu and other fish species still remained unknown. In the Indian major carps, TLR3 and TLR4 were previously reported to recognize poly I:C and LPS receptor, respectively, resulting in the induction of various cytokines (Basu et al., 2013; Samanta et al., 2013). In various animals, activation of TLRs signaling also resulted in the induction of pro-inflammatory gene expression (Vogel et al., 2003; Neill and Bowie, 2007). In addition to TLRs, NOD1 and NOD2 were also activated by poly I:C and LPS treatments in Indian major carp, grouper, and grass carp (Chen et al., 2010; Hou et al., 2012; Swain et al., 2012, 2013). The iE-DAP and MDP are known ligands of NOD1 and NOD2, respectively. They activate NOD signaling and induce both type-I and type-II IFN (Takahashi et al., 2006; Rosenzweig et al., 2009; Chang et al., 2011). These observations together suggest that IFN-γrel gene in rohu might be regulated through either TLR or NOD signaling or interplay between them. Further works are necessary to understand the mechanism of IFN-γrel gene expression.
In mammals, IFN-γ was reported to bind its receptor complex and activates the JAK-STAT signaling pathway (Schroder et al., 2004; Dai et al., 2008). In rohu, IFN-γrel stimulated IFN-γ receptor in PBLs, suggesting that both IFN-γ and IFN-γrel may bind to common receptors to activate JAK-STAT signaling pathway. In goldfish macrophages, IFN-γrel was also reported to induce iNOS gene expression and nitric oxide production, resulting in antimicrobial defense (Grayfer et al., 2010). In rohu PBLs, rIFN-γrel also induced iNOS gene expression. These data in rohu and goldfish together suggest the important function of IFN-γrel in antimicrobial defense in various fish species and warrant further studies to be explored.
This article describes about the IFN-γrel gene in rohu, a fish species of highest commercial importance in the Indian subcontinent. The rohu IFN-γrel gene consisted of four exons with three intervening introns and the full-length cDNA comprised 927 bp nucleotides. After LPS, poly I:C, iE-DAP, MDP, and bacterial infections, IFN-γrel gene was significantly upregulated in various tissues, suggesting its important role during diseases. The recombinant rohu IFN-γrel protein induced IFN-γ receptor and iNOS gene expression in PBLs, indicating its critical role as an anti-microbial cytokine. This study might be helpful to conduct further research in developing therapeutic/preventive measures against fish diseases.
Footnotes
Acknowledgments
This work was supported by the grant of National Agricultural Science Fund (NASF/BS-4003) of the Indian Council of Agricultural Research (ICAR), Govt. of India. The authors express their sincere gratitude to Dr. A. Bandyopadhay, former NC, and Dr. P.K. Agrawal, ADG, NASF, for suggestions and Dr. S.S. Mishra, Head of Division, Fish Health Management Division, CIFA, for the research facilities.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.