
Abstract
Large intergenic long noncoding RNA p21 (lincRNA-p21) has recently shown to play an important role in biological functions. However, the biological role of lincRNA-p21 in vascular endothelial cells remains unclear. In the present study, we investigated the role of lincRNA-p21 in vascular endothelial cells through gain- and loss-of-function studies and found that lincRNA-p21 promoted cell apoptosis and induced cell cycle progression. Furthermore, lincRNA-p21 acted as an endogenous sponge by directly binding to miR-130b and decreased miR-130b expression. In addition, miR-130b reversed the inhibitory effect of lincRNA-p21 on the growth of vascular endothelial cells. Taken together, our data highlight the pivotal role of lincRNA-p21 in the growth of vascular endothelial cells.
Introduction
L
In recent studies, large intergenic long noncoding RNA p21 (lincRNA-p21) has been shown to regulate apoptosis in response to p53 signaling (Huarte et al., 2010). LincRNA-p21 acts in concert with hnRNP-K as a coactivator for p53-dependent p21 transcription and affects global gene expression (Bao et al., 2015). LincRNA-p21 deficiency deregulated expression by altering the chromatin state of some polycomb target genes, increased proliferation rates, and enhanced reprogramming efficiency (Dimitrova et al., 2014). In addition, lincRNA-p21 is a hypoxia-responsive lncRNA and is essential for hypoxia-enhanced glycolysis (Yang et al., 2014a). Nevertheless, the role of lincRNA-p21 in vascular endothelial cells is not well established.
Recently, many RNA transcripts have been reported to function as competing endogenous RNAs (ceRNAs) by competitively binding common microRNAs (Sen et al., 2014; Tay et al., 2014; Poliseno and Pandolfi, 2015). These ceRNAs or natural miRNA sponges have an active role in regulating miRNA availability and form mutual regulatory networks. Previous reports have implicated diverse RNA species, including mRNAs and ncRNAs, as ceRNAs in human development and diseases (Xia et al., 2014; Yang et al., 2014b; Yuan et al., 2014). However, whether lincRNA-p21 functions as a ceRNA is still unknown.
In the present study, we investigated the role of lincRNA-p21 in vascular endothelial cells through gain- and loss-of-function studies, luciferase reporter assays, and RNA immunoprecipitation (RIP) assays. The results suggested that the interaction between miR-130b and lincRNA-p21 is associated with cell proliferation and apoptosis.
Materials and Methods
Cell cultures
SVEC4 cell line, the mouse vascular endothelial cell, was obtained from the American Type Culture Collection (ATCC). Cells were cultured at 37°C in an atmosphere containing 5% CO2 and in Dulbecco's modified Eagle's medium (Invitrogen) supplemented with 10% fetal bovine serum.
RNA extraction and quantitative real-time PCR
Total RNA was isolated using the TRIzol reagent (Invitrogen). First-strand cDNA was generated using the M-MLV Reverse Transcriptase (Invitrogen). Real-time polymerase chain reaction (PCR) was performed in the StepOne Real-Time PCR System (Applied Biosystems) using SYBR Green (Takara). GAPDH mRNA was employed as an endogenous control for mRNA and lncRNA. For microRNA analysis, real-time PCR was performed as above, using TaqMan MicroRNA assays according to the manufacturer's instructions (Applied Biosystems). The relative expression of RNAs was calculated using the comparative Ct method. Primers used for quantitative real-time PCR (qRT-PCR) are listed as follows: lincRNA-p21-F: 5′-CTCCTTCTTGTGGTTGTGACT-3′, lincRNA-p21-R: 5′-CCGGCCTACACCAGTTATTT-23′.
Cell viability assay
SVEC4 cells were seeded at a density of 1 × 103 cells per well in 96-well plates. The viability of the cells was assessed from three replicates by the MTT (3-2,5-diphenyl tetrazolium bromide) assay (Sigma) for 7 days. The resulting colored solution was quantified using an ELx800 Absorbance Microplate Reader (BioTek) at 570 nm with a reference wavelength of 630 nm.
Cell cycle assay
The cell cycle was analyzed using an In Situ Cell Proliferation Kit FLUOS (Roche) according to the manufacturer's instruction. Briefly, cells were labeled with BrdU for 40 min before trypsinization. Cells were fixed by adding 70% ice-cold ethanol. Fixed cells were incubated with 4 M HCl at room temperature for 30 min and then washed with phosphate buffered saline. The cells were then incubated with anti-BrdU-FLUOS antibody at room temperature for 45 min. Then cell suspensions were incubated with 7-AAD for 5 min and immediately analyzed by flow cytometry. The data were analyzed by the FlowJo software (Tree Star).
Apoptosis assay
Cells were stained with fluorescein isothiocyanate-conjugated Annexin V and 7-AAD (Apoptosis Detection Kit; KeyGEN), as suggested by the manufacturer. Cells were then analyzed with a flow cytometer, and the data were studied using FlowJo software (Tree Star).
Isolation of cytoplasmic and nuclear
Cytoplasmic and nuclear RNA were isolated and purified using the Cytoplasmic & Nuclear RNA Purification Kit (Norgen) according to the manufacturer's instructions.
Lentiviral construction and cell transfection
To generate clones stably overexpressing lincRNA-p21, SVEC4 cells were first infected at a multiplicity of infection of 20 with a lentiviral vector encoding a full-length human lincRNA-p21 gene sequence or an empty lentiviral vector control. Stable clones were selected for 2 weeks using puromycin, and the expression level of lincRNA-p21 was determined by qRT-PCR.
The pLKO.1 empty vector was purchased from Open Biosystems. We constructed two independent, lentivirus-based shRNA expression constructs to knockdown the lincRNA-p21 expression. The negative control lentiviral vector containing nonsilencing shRNA was used. SVEC4 cells were infected with either of the lentiviral vectors encoding specific shRNA sequences or the negative control vector. The efficiencies of RNA interference were determined by qRT-PCR. The target sequences were listed as follows: sh1: 5′-GTGGTTGTGACTTCCATAA-3′, sh2: 5′-GACTCACTGAATCCTCAAA-3′.
Luciferase reporter assay
To construct plasmids used in dual-luciferase reporter assays, wild-type lincRNA-p21 cDNA (containing miR-130b binding site) and a mutant lincRNA-p21 sequence (mutant in miR-130b binding site) were cloned into pmirGLO vectors, and the resulting vectors were designated respectively as pmirGLO-lincRNA-p21 and pmirGLO-lincRNA-p21-mut. The pmirGLO, pmirGLO-lincRNA-p21, or pmirGLO-lincRNA-p21-mut was cotransfected with miR-130b mimics or miR NC into SVEC4 cells by lipofectamine-mediated gene transfer. The relative luciferase activity was normalized to Renilla luciferase activity 48 h after transfection.
RNA immunoprecipitation
SVEC4 cells were cotransfected with pcDNA3.1-MS2, pcDNA3.1-lincRNA-p21-MS2, or pcDNA3.1-lincRNA-p21-mut-MS2 and pMS2-GFP (Addgene). After 48 h, cells were used to perform RIP assays using a green fluorescent protein (GFP) antibody (Abcam) and Magna RIP™ RNA-Binding Protein Immunoprecipitation Kit (Millipore) according to the manufacturer's instructions. Briefly, cells were lysed in a lysis buffer containing a protease inhibitor cocktail and RNase inhibitor. Magnetic beads were preincubated with an anti-GFP antibody or anti-rabbit IgG for 30 min at room temperature, and lysates were immunoprecipitated with beads at 4°C overnight. RNA was purified from RNA protein complexes bound to the beads and was then analyzed by qRT-PCR.
RNA pull-down
LincRNA-p21 and lincRNA-p21-mut were in vitro transcribed, biotin-labeled with the Biotin RNA Labeling Mix (Roche) and T7 RNA polymerase (Roche), and purified with an RNeasy Mini Kit (Qiagen), respectively. One milligram of whole-cell lysates from SVEC4 cells were incubated with 3 μg of purified biotinylated transcripts for 1 h at 25°C and complexes were isolated with streptavidin agarose beads (Thermo). The RNA present in the pull-down material was detected by qRT-PCR analysis.
Statistical analyses
All statistical analyses were performed using SPSS software. For comparisons, Student's t-test (two-tailed) was performed as indicated. A p-value < 0.05 was considered significant.
Results
LincRNA-p21 inhibits cell proliferation
To explore the biological function of lincRNA-p21, we next performed gain- and loss-of-function studies in SVEC4 cells. We developed SVEC4 cells with stably overexpressed and knockdown lincRNA-p21 expression (Fig. 1A, B). We found that lincRNA-p21 overexpression decreased the proliferation of SVEC4 cells compared to control cells containing the empty vector by performing MTT assays (Fig. 1C). In contrast, knockdown endogenous lincRNA-p21 expression significantly increased the proliferative capacity of SVEC4 cells (Fig. 1D).
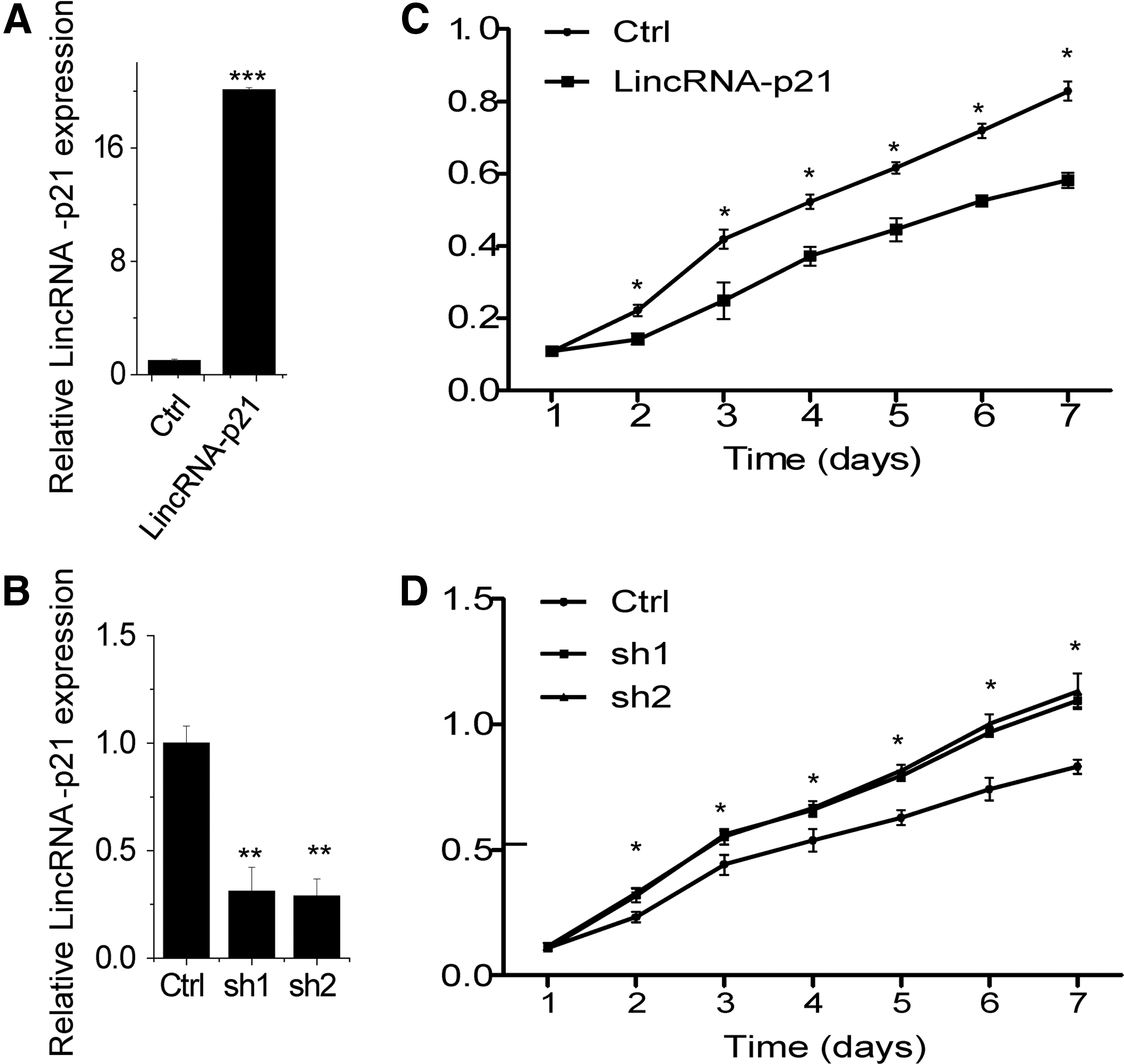
LincRNA-p21 inhibits SVEC4 cell proliferation.
LincRNA-p21 promotes cell apoptosis and induces cell cycle progression
To gain insight into the mechanism by which lincRNA-p21 inhibits cell proliferation, we performed fluorescence-activated cell sorting (FACS) to analyze differences in cell cycle distributions following lincRNA-p21 overexpression or knockdown. We found that lincRNA-p21 knockdown drove progression beyond the G1/S transition, whereas significant G1/S arrest was observed in lincRNA-p21-overexpression SVEC4 cells (Fig. 2A, B). These results demonstrated that lincRNA-p21 inhibits cell proliferation by facilitating cell cycle progression.
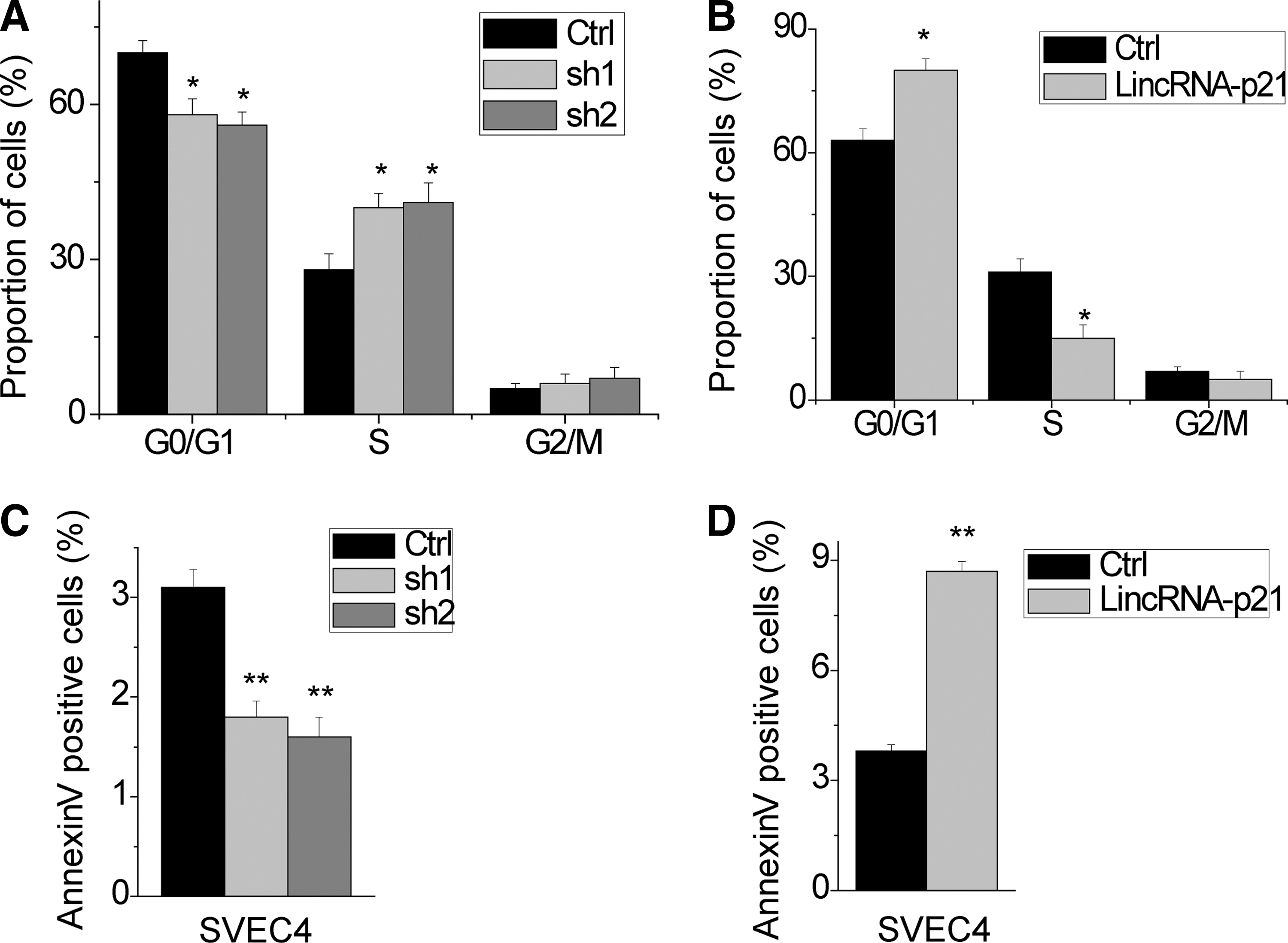
LincRNA-p21 reduces cell cycle progression and promotes cellular apoptosis in SVEC4 cells.
We speculated that lincRNA-p21 may be critical for cell apoptosis in SVEC4 cells. To test this hypothesis, we performed FACS analysis in the SVEC4 cell stained for Annexin V and 7-AAD to detect apoptosis. The results showed that overexpression of lincRNA-p21 had a significantly higher percentage of Annexin V-positive cells than control cells (Fig. 2C). In contrast, we observed that knockdown of lncRNA-p21 had protective effects to apoptosis (Fig. 2D). Taken together, these data indicate that lincRNA-p21 affects vascular endothelial cell proliferation.
LinRNA-p21 locates in both of the cytoplasm and nucleus
The cellular location of lncRNA determined the way by which lncRNA exerts its function. We first performed cellular fractionation to analyze the subcellular localization of lincRNA-p21. LincRNA-p21 located in both the cytoplasm and nuclear (Fig. 3), consistent with a potential role in epigenetic and posttranscriptional regulation.
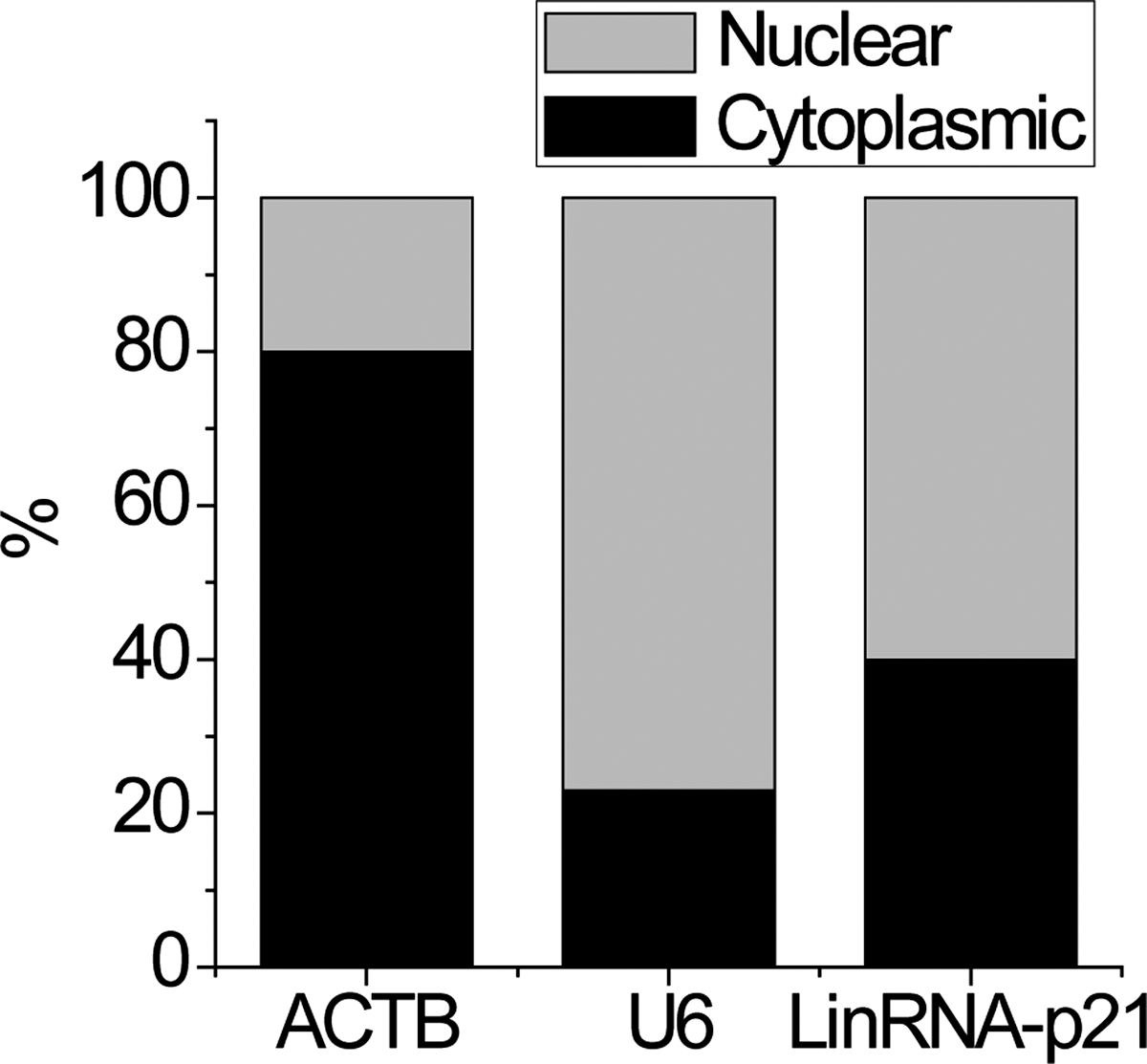
The subcellular location of lincRNA-p21. Representative analysis of lincRNA-p21 distribution by cellular fractionation of SVEC4 cells. ACTB mRNA and U6 RNA served as controls for cytoplasmic and nuclear RNAs, respectively.
LinRNA-p21 may function as a ceRNA of miR-130b
Interactions between lncRNAs and miRNAs provide an additional regulation of gene expression (Yuan et al., 2014; Cao et al., 2015). Using the TargetScan prediction algorithm, we found a set of miRNAs that potentially bind to lincRNA-p21 (Fig. 4A). Among these candidate miRNAs, the miR-130b arouses our interest. The miR-130b has been reported to promote proliferation through interacting with the 3′-untranslated region of PTEN mRNA and indirectly regulated the level of phosphorylated Akt (Yu et al., 2015). To validate the direct binding between miR-130b and lincRNA-p21 at endogenous levels, we perform an RIP assay to pull down endogenous miRNA associated with lincRNA-p21. The result showed that the lincRNA-p21 RIP in SVEC4 cells was significantly enriched for miR-130b compared to the empty vector (pcDNA3.1-MS2), IgG, and lincRNA-p21 with the mutation of miR-130b targeting site (Fig. 4B–D). The association between lincRNA-p21 and miR-200s was further validated by affinity pull-down of endogenous miR-130b using in vitro transcribed biotin-labeled lincRNA-p21 (Fig. 4E).
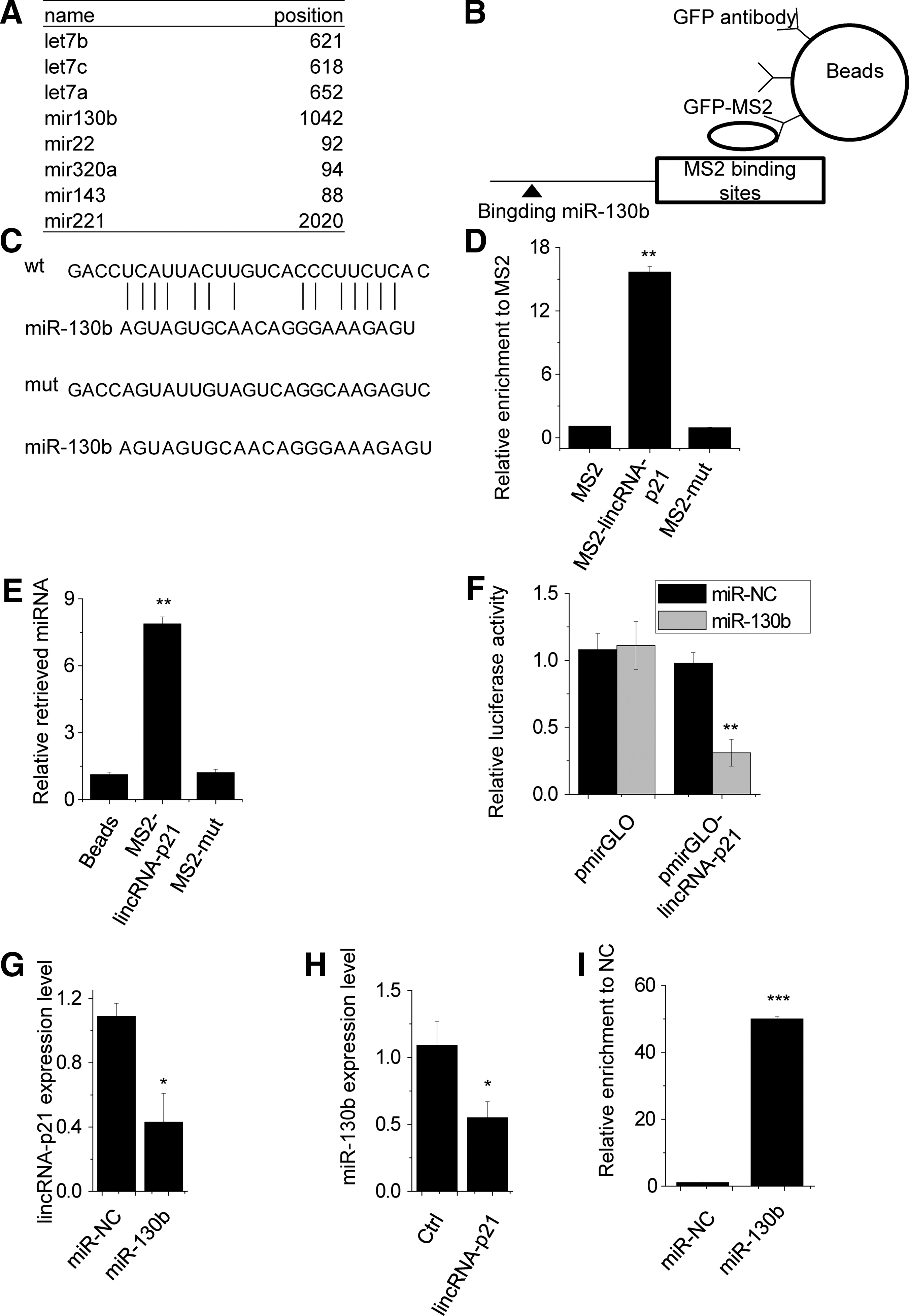
LincRNA-p21 interacts with miR-130b.
Furthermore, dual-luciferase assays showed a significant decrease in luciferase activities following cotransfection of miR-130b and the wild-type lincRNA-p21 expression vector, but not a mutant lincRNA-p21 (Fig. 4F). Next, we clarified the regulatory relationship between lincRNA-p21 and miR-130b. Overexpression of miR-130b significantly decreased lincRNA-p21 expression (Fig. 4G), and overexpression of lincRNA-p21 inhibits miR-130b expression (Fig. 4H).
The microRNAs are known to bind their targets and cause RNA degradation in an AGO2-dependent manner (Karginov and Hannon, 2013). To determine whether lincRNA-p21 was also regulated by miR-130b in such a manner, we performed anti-AGO2 RIP in SVEC4 cells transiently overexpressing miR-130b. Endogenous lincRNA-p21 pull-down by AGO2 was specifically enriched in miR-130b-transfected cells (Fig. 4I), supporting that miR-130b is a bona fide lincRNA-p21-targeting microRNA. These data demonstrated that the miR-130b bound to lincRNA-p21 induced the reduction of lincRNA-p21 and may function as a ceRNA.
LincRNA-p21 inhibits cell proliferation through binding miR-130b
To investigate whether lincRNA-p21 inhibits cell proliferation through degradation of miR130b, rescue experiments were performed. We overexpressed miR-130b in lincRNA-p21 overexpression cells (Fig. 5A). Overexpression of wild-type lincRNA-p21 inhibited proliferation and induced G1/S arrest and apoptosis (Fig. 5B–D). Ectopic expression of miR-130b in lincRNA-p21 overexpressed cells abrogated this effect (Fig. 5C, D). Together, these results demonstrate that linRNA-p21 exerts its function, at least in part, through regulating the miR-130b expression.
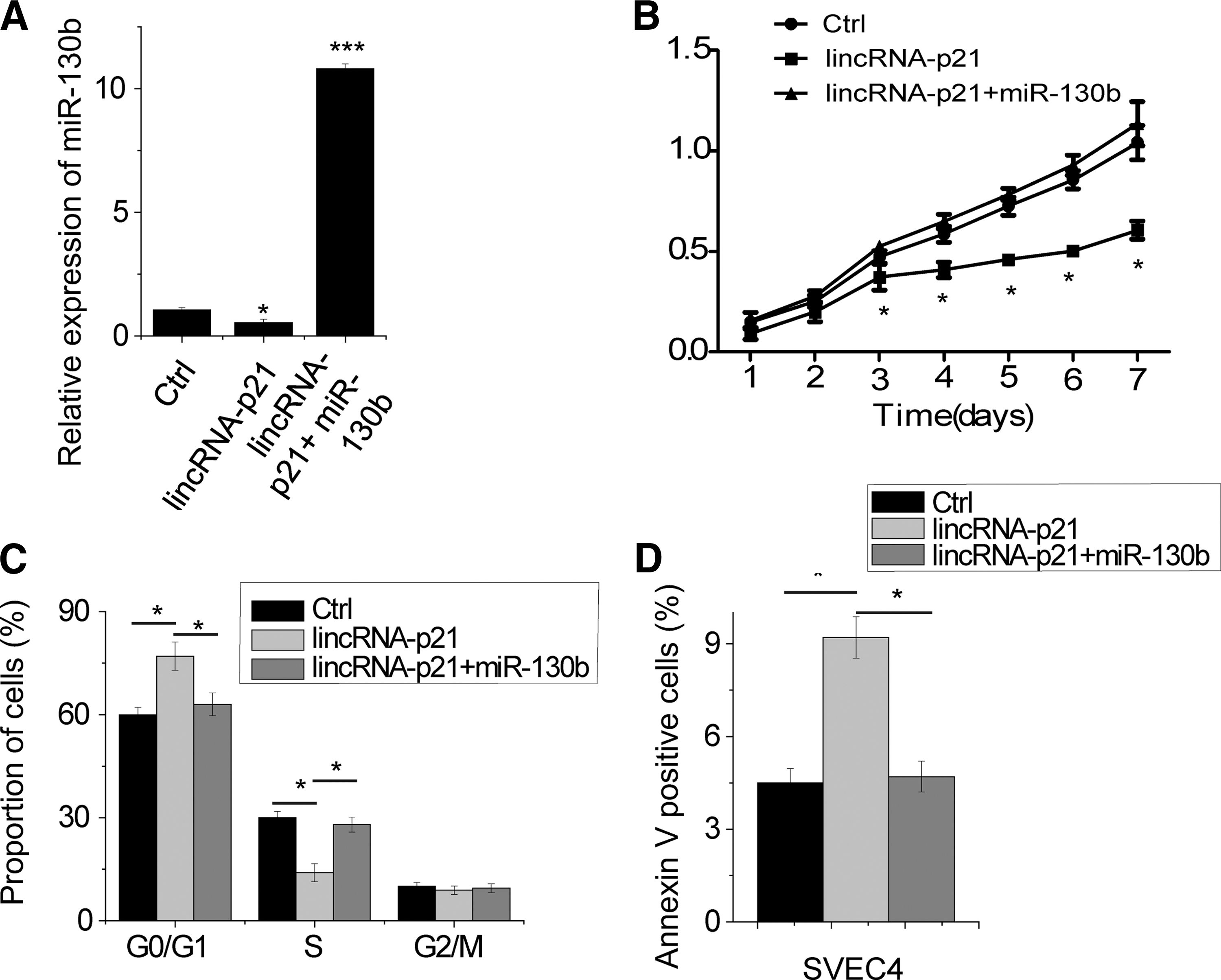
lincRNA-p21 inhibits cell proliferation through binding miR-130b.
Discussion
The mammalian genome encodes thousands of lncRNAs, many of which participate in a series of biological processes ranging from normal embryonic development to disease (Angrand et al., 2015; Philippen et al., 2015; Yarmishyn and Kurochkin, 2015). Understanding the exact mechanisms of regulation pattern of lncRNAs could help to shed light on the role of the same lncRNAs in other contexts, particularly in diseases. In this study, applying loss- and gain-of-function approaches, we found that lincRNA-p21 plays a key role in cell cycle and apoptosis regulation. Furthermore, it was shown that lincRNA-p21 was associated with miR-130b and lincRNA-p21 exerted its function at least in part through binding and suppressing of miR-130b. Previous studies have suggested that a number of lncRNAs were associated with miRNA. For example, lncRNA-ATB and CCAT1, two well-characterized lncRNAs, both interacted with miRNAs to take part in posttranscriptional gene regulation (Yuan et al., 2014; Ma et al., 2015). LncRNA-ATB, which was known for its role in promoting tumor metastasis, binds to miR-200s family to regulate ZEB1 and ZEB2 expression (Yuan et al., 2014). CCAT1 interacted with let-7 for derepression of its endogenous targets HMGA and c-Myc (Ma et al., 2015). In combination with these findings, the functional interaction between lincRNA-p21 and miR-130b further emphasizes the central role of lncRNA in the regulation of biological function and suggests that interaction with miRNAs may be an important mechanism by which lncRNAs exert their functions.
Recent studies have reported that posttranscriptional gene regulation involves coregulatory crosstalk between RNA transcripts, which compete for common pools of miRNA molecules (Karreth and Pandolfi, 2013; Shi et al., 2013; Tay et al., 2014). For example, the processed pseudogene PTENP1, which was highly homologous to the tumor suppressor gene PTEN, was shown to posttranscriptionally regulate PTEN expression in a miRNA-dependent manner. PTENP1 exerted its tumor suppressive properties through sequestration of miR-21, providing additional support for the functional relevance of PTENP1 as a tumor suppressive ceRNA for PTEN (Chen et al., 2015; Guo et al., 2015). In addition to their roles in various cancers, lncRNA ceRNAs have been implicated in human development. A muscle-specific lncRNA, lincMD1, which was activated on myoblast differentiation and controls muscle differentiation in human myoblasts through its ceRNA activity, has been identified (Cesana et al., 2011). LincMD1 sequestered miR-133 and miR-135 and effectively regulated the expression of MAML1 and MEF2C mRNAs, which encoded transcription factors known to activate muscle-specific gene expression. These reports suggested that ceRNA regulation is of crucial importance in diseases and development. In the present study, for the first time, we found that lincRNA-p21 exerts its function as a ceRNA of miR-130b to regulate cell cycle and apoptosis. These findings highlight the pivotal role of lincRNA-p21 in the growth of vascular endothelial cells. Whether lincRNA-p21 can serve as ceRNAs for other miRNAs needs further studies, which will be helpful for exploring the function of lincRNA-p21 in diseases.
Conclusion
Altogether, our study constitutes a resource that may guide others to investigate the function of lincRNAs in mRNA posttranscriptional regulation. Future work will further delineate the networks controlled by lncRNAs, which will be helpful to develop a therapeutic target for patients with disease.
Footnotes
Acknowledgment
This study was supported by grants from the National Natural Science Foundation of China (No. 81470387).
Disclosure Statement
No competing financial interests exist.