
Abstract
We investigated the suppressive effects of crocin alone and in combination with hyperthermia (HT) on proliferation of breast cancer cells. Cell viability, colony formation ability, and apoptosis were assessed by 3-(4,5-dimetylthiazol-2-Yl)-2,5–diphenyltetrazolium bromide (MTT), soft agar, Hoechst 33258 staining, and percentage of lactate dehydrogenase (LDH) release methods, respectively. The mRNA levels Hsp27, Hsp70, Hsp90, Bax, and Bcl-2 were measured by quantitative real-time polymerase chain reaction (qRT-PCR). Hsp70 and Hsp90 proteins were determined using enzyme-linked immunosorbent assay (ELISA) technique. Crocin in combination with HT significantly inhibited the proliferation of cancer cells in a dose- and time-dependent manner. There was a degree of synergism in the combined treatment. However, crocin did not show the high cytotoxic effect on normal cells. This treatment decreased colony formation of cancer cells up to 94%. Changed nuclear morphology and increased LDH indicated that crocin combined with HT has a more apoptotic effect than crocin alone. Furthermore, in treated cells Bax/Bcl-2 ratio markedly increased, whereas expression of heat-induced genes decreased. Also, the Hsp70 and Hsp90 proteins decreased in the treated cells. Our study indicated that combination of crocin and HT has strong antiproliferative and apoptotic activities against breast cancer cells. Hence, it is suggested that more studies are warranted to apply crocin as a possible, safe, and promising anticancer agent in cancer.
Introduction
C
It has been reported that a temperature between 41°C and 44°C does not affect normal cells while significantly inhibiting cancer cell growth (Ahmed and Zaidi, 2013). Heat therapy induces various physiological changes in cells, such as alteration in membrane permeability, modification of cytoskeletal system, changes in macromolecule synthesis, intracellular signal transduction, and inhibition of DNA repair (Ahmed and Zaidi, 2013). Interestingly, many clinical trials have shown that combining HT with other cancer therapies can yield durable complete responses (Jones et al., 2005). However, HT is inevitably associated with heat shock proteins (HSPs). Importantly, increased expression of HSPs such as Hsp70 and Hsp90 in cancer cells has been reported (Gress et al., 1994). Reducing HSP levels in tumor cells induces cell death and/or sensitizes them to cytotoxic agents without deleterious effects on normal cells (Powers et al., 2008).
On the other hand, medicinal plants play an important role in cancer prevention and treatment (Bhandari, 2015; Hoshyar et al., 2015a). Among various herbs, the anticancer effects of saffron (Crocus sativus L.) and its secondary metabolites were shown in both in vitro and in vivo models (Bathaie et al., 2007, 2013; Hoshyar et al., 2013; Amin et al., 2016). Crocin is the main water-soluble carotenoid of saffron stigma that effectively suppresses the proliferation of cancer cells through apoptosis induction without affecting normal cells (Hoshyar et al., 2013). Recently, crocin has been used in combination with lower doses of other modern or conventional anticancer agents to inhibit cell proliferation (Li et al., 2013).
Apoptosis or programmed cell death is one of the cellular responses to anticancer agents, induced by HT or herbs (Ahmed and Zaidi, 2013). So, targeted inhibition of the antiapoptotic pathway is an attractive approach for combination cancer therapies (Ahmed and Zaidi, 2013). The aim of the present study is to investigate the cytotoxic and apoptotic effects of crocin alone or its combination with HT on breast adenocarcinoma (MDA-MB-468) cells.
Materials and Methods
Crocin preparation
Crocin is isolated and purified from the Iranian saffron stigma according to the registered method in the previous report (Patent No. 020577; Bolhasani et al., 2005).
Cell culture
Human breast cancer (MDA-MB-468) and normal breast epithelial (MCF10-A) cell lines were provided from Iranian Biological Resource Center (IBRC). Dulbecco's modified Eagle's medium (DMEM), fetal bovine serum (FBS), phosphate buffered saline (PBS), trypsin/EDTA solution, 3-(4,5-dimetylthiazol-2-Yl)–2,5–diphenyltetrazolium bromide (MTT), and dimethyl sulfoxide (DMSO) were purchased from Gibco BRL and Sigma, respectively. The cells were cultured in DMEM supplemented with 10% FBS, 100 units/mL penicillin, and 100 mg/mL streptomycin. Both cell lines were grown at 37°C in humidified atmosphere containing 5% CO2. For combined treatment, first MDA-MB-468 cells were incubated at 43°C for 2 h in a CO2 incubator with a humid atmosphere; second, cells were exposed to various concentrations of crocin (0–5 mg/mL) at different time intervals (0–72 h). Cells treated with crocin (3 and 4 mg/mL after 24 and 48 h) were washed with PBS. Then, morphological changes were examined at indicated times using a regular phase-contrast microscope.
Cell viability assay
The cytotoxic effects of crocin alone and combined with HT on normal and cancer cells were determined with MTT assay. The absorbance was recorded at 570 nm with a plate reader (BioTek, Epoch). The IC50 values (its minimum concentration to reduce cell viability to 50%) of crocin alone and combined with HT, were calculated using the dose- and time-dependent curves by linear interpolation (Hoshyar et al., 2015b).
The synergy index (q) was measured according to the following formula:
E(A + B) represented the inhibitory rate of crocin combined with HT, EA and EB were inhibitory rates of crocin and HT alone, respectively (Li et al., 2013).
Soft agar colony formation assay
Anchorage-independent growth was evaluated through the soft agar growth assay. In 6-cm Petri dishes, MDA-MB-468 cells (1800 cell/dish) were seeded in the top layer containing 0.35% agar, DMEM, and 10% FBS in the absence and presence of different concentrations of crocin (1.5 and 3 mg/mL). The bottom layer contained 0.5% agar, DMEM, and 10% FBS. The Petri dishes were covered with 1.5 mL of covering medium (DMEM, 10% FBS with or without crocin) and were maintained overnight at 37°C. Then, these Petri dishes were incubated at 43°C for 2 h. This was continued for additional 16 days at 37°C. The colonies were stained with 1 mL of 0.005% Crystal Violet and counted under phase contrast microscopy with low magnification.
Lactate dehydrogenase leakage measurement
To evaluate cell death levels, supernatants were measured for lactate dehydrogenase (LDH) leakage. After HT, cells were treated with 1.5 and 3 mg/mL crocin for 24–48 h. The culture medium was aspirated and centrifuged at 2000 g for 10 min to obtain a cell-free supernatant. Percentage of LDH release was monitored with an LDH Leakage Detection Kit (Sigma).
Apoptosis analysis
Hoechst staining was used to determine the proapoptotic effect of crocin alone and in combination with HT. After 24 h, cells were spotted onto slides and examined by a fluorescence microscopy with DAPI filter (Abedini et al., 2004).
RNA isolation and real-time RT-PCR analysis
The cells were treated with the most effective dose of crocin (1.5 mg/mL) for 12–48 h. Total RNA was isolated using the GenMark Kit and the first-strand cDNA was synthesized according to the manufacturer's procedure of Thermo Scientific RevertAid First-Strand cDNA Synthesis Kit (Thermo, EU). Gene expression levels were determined by real-time RT-PCR by step one real-time PCR (Applied Biosystems) using SYBR Green Master Mix (Parstous). The forward and reverse PCR primers are listed in Table 1. To normalize relative levels of gene expression, β-actin was utilized as an internal reference control and the comparative threshold cycle (2−ΔΔCT) method was applied to quantify the mRNA levels of interested genes (Hoshyar et al., 2015a).
qRT-PCR, quantitative real-time polymerase chain reaction.
ELISA analysis
Cells were treated with crocin (1.5 mg/mL) for 24–72 h after HT. The cell supernatants were collected after 24, 48, and 72 h incubation and centrifuged at 3000 rpm for 20 min. Finally, the levels of Hsp70 and Hsp90 proteins in the cell culture supernatants were determined using the Sandwich Enzyme-Linked Immunosorbent Assay (ELISA) Kit (Chongqing Biospes Co., Ltd.).
Statistical analysis
Results are expressed as the mean ± SD of at least three independent experiments. Data were analyzed using one-way ANOVA and followed by Tukey's post hoc test to assess differences between experimental groups (PRISM 5.0; GraphPad Software, Inc.). Statistical significance was inferred at p < 0.05.
Results
Viability of MDA-MB-468 cells
HT significantly decreased proliferation of cancer cells in a time-dependent manner (Supplementary Fig. S1; Supplementary Data are available online at
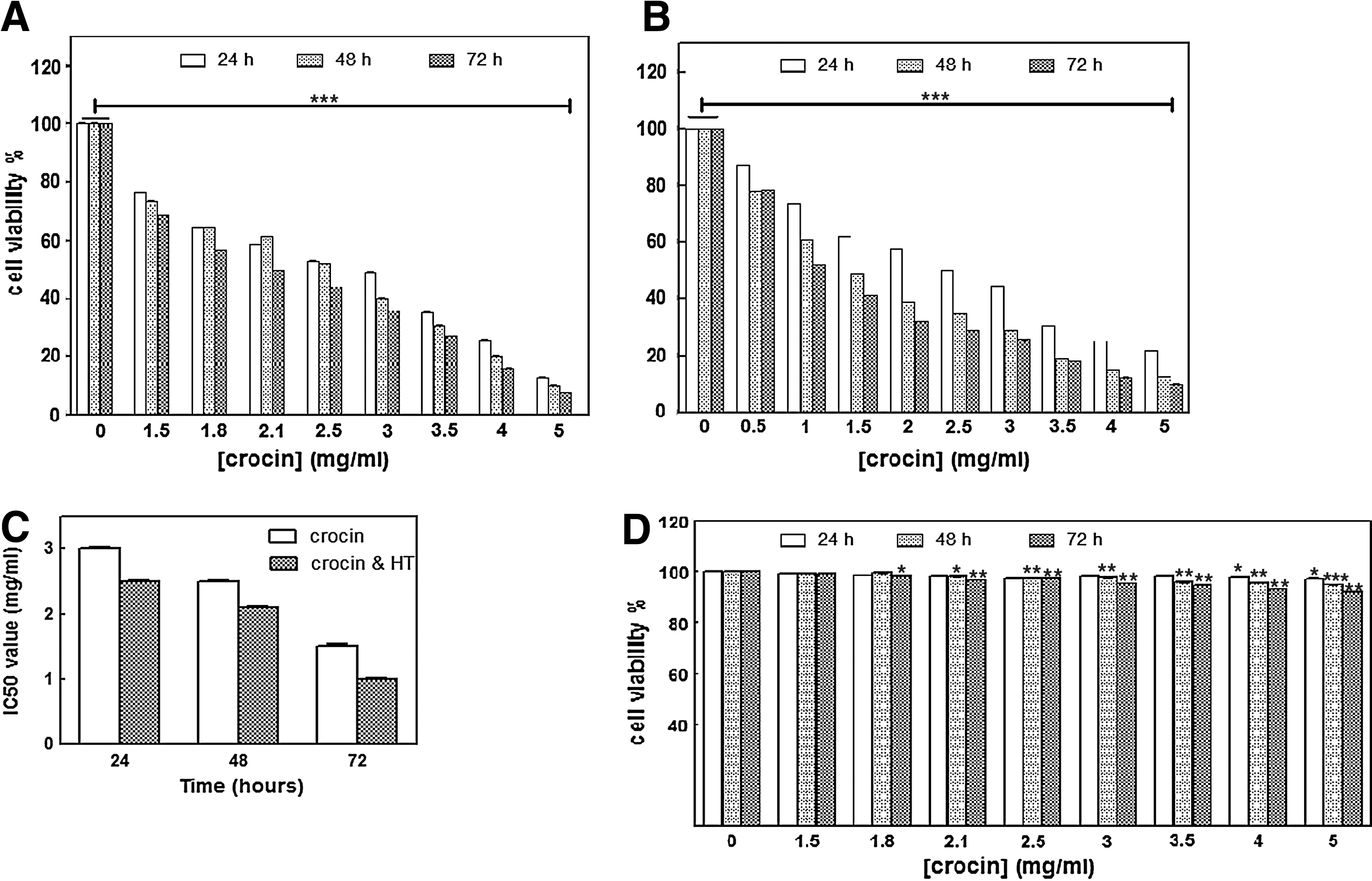
The effect of
Colony formation of MDA-MB-468 cells
Treatment of MDA-MB-468 cells with crocin (1.5 and 3 mg/mL) combined with HT resulted in significant decreases (94% and 95%; p < 0.01) in the number of colonies when compared with the control nonheat cells, respectively (Table 2; Fig. 2).
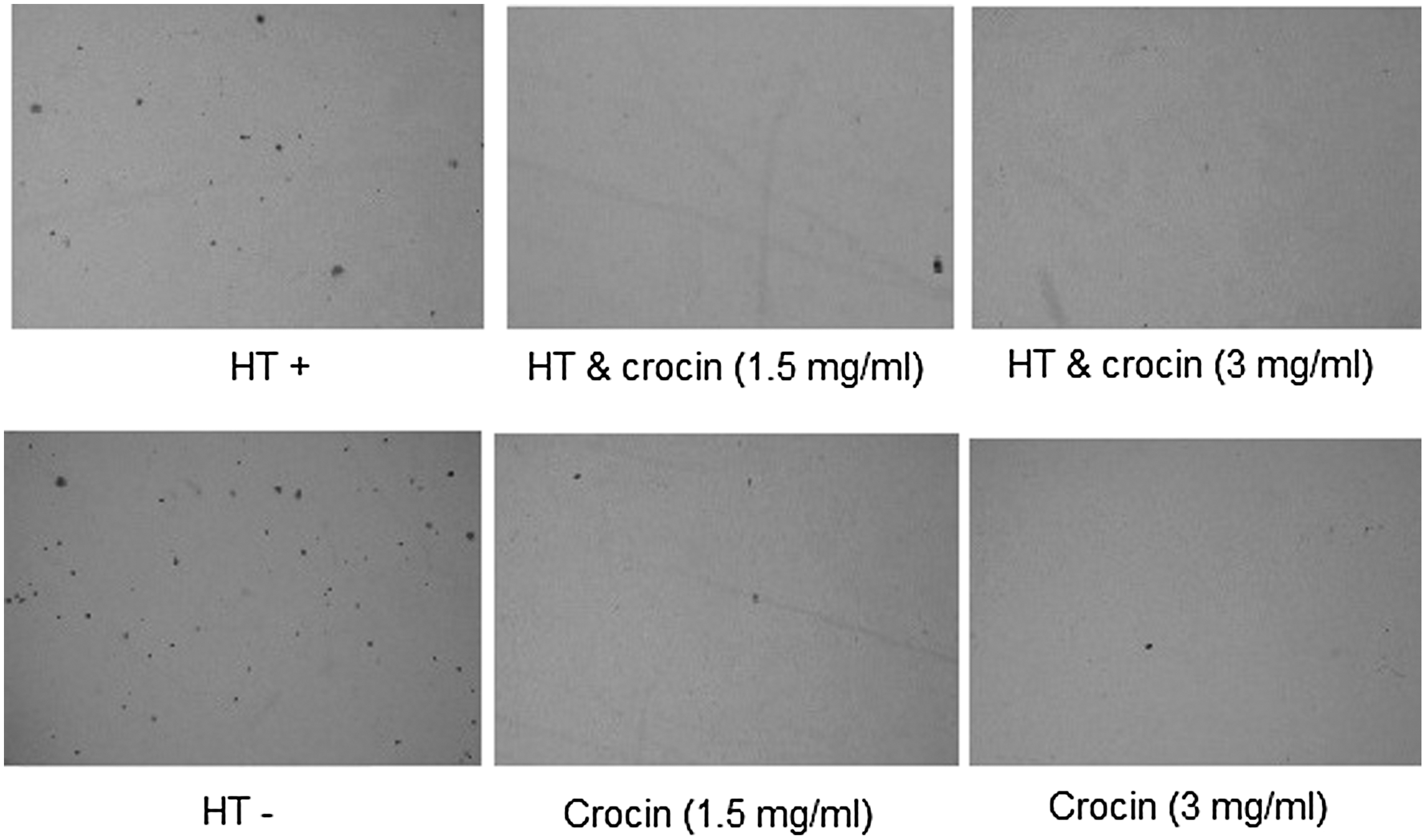
Combination effects of crocin (0–3 mg/mL) and HT (43°C for 2 h) on colony formation of MDA-MB-468 cells in soft agar after 16 days.
The results are represented as the mean ± SD (** p < 0.01).
HT, hyperthermia.
Apoptosis of MDA-MB-468 cells
Results of Hoechst staining indicated that HT increased the apoptotic impact of crocin in a dose-dependent manner (28.26% vs. 43.66% and 45.59% vs. 70.34% of apoptotic cells of total cells after 1.5 and 3 mg/mL crocin treatments, respectively). Percentage of LDH released in the treated cancer cells with crocin and HT significantly increased (67.5% at 48 h) over time compared with control nonheat cells (9% after 24 h incubation, Fig. 3).
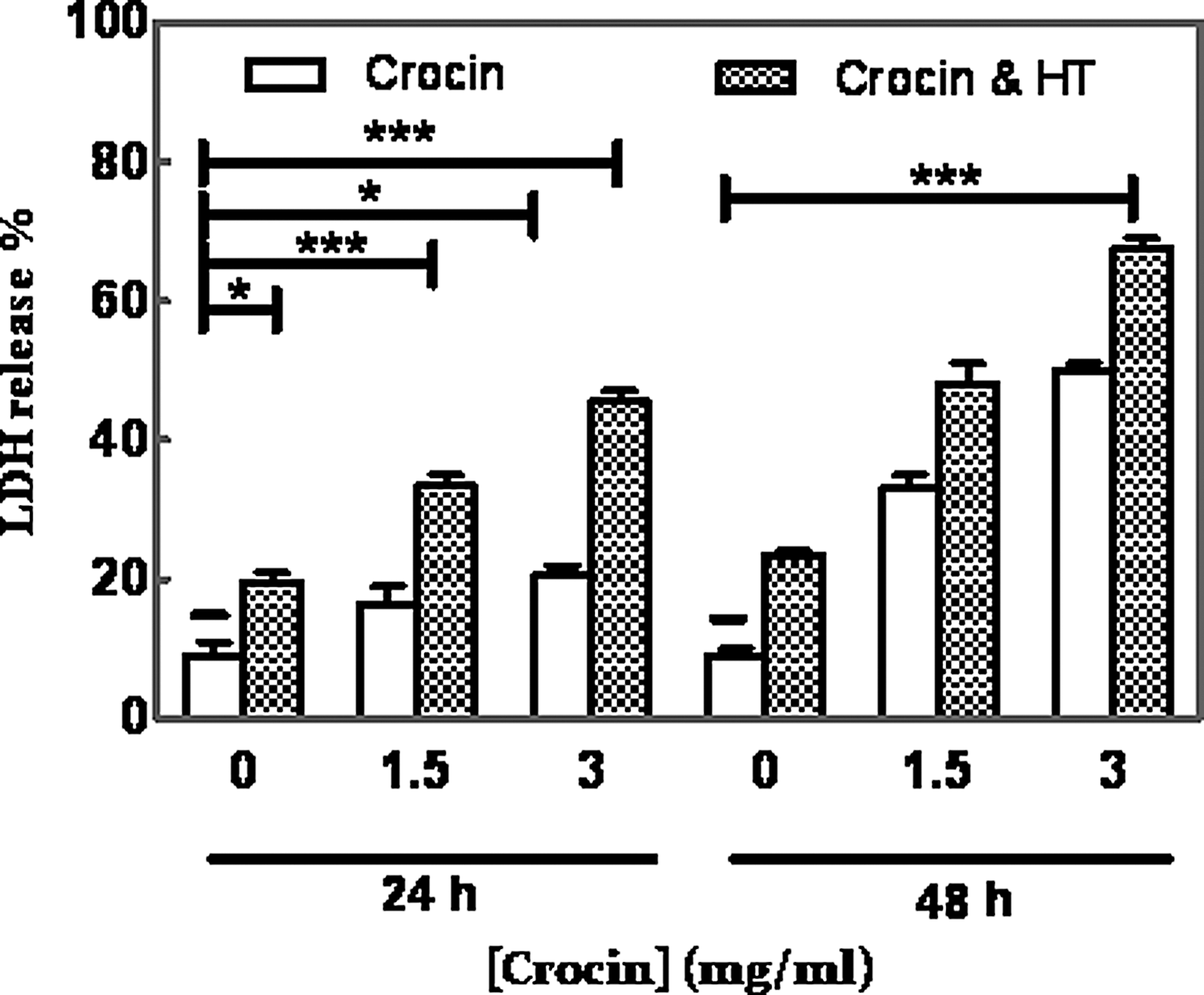
Combination effects of crocin (0–3 mg/mL) and HT (43°C for 2 h) on percentage of LDH release from MDA-MB-468 cells after 24 and 48 h. The results are represented as the mean ± SD (*p < 0.05; ***p < 0.001). LDH, lactate dehydrogenase.
Expression of apoptosis-related genes
Quantitative real-time PCR results showed that HT significantly increased mRNA levels of Hsp27, Hsp70, and Hsp90 (1.8-, 2.5-, and 3.7-fold) than the levels in control nonheat cells, respectively. The most effective dose of crocin (1.5 mg/mL) reduced HT-induced HSPs expression in a time-dependent manner compared with control nonheat cells (p < 0.001; Fig. 4A–C). Additionally, data showed that combination of crocin and HT significantly increased (76.8%) the Bax/Bcl-2 ratio in comparison with crocin (Fig. 5).
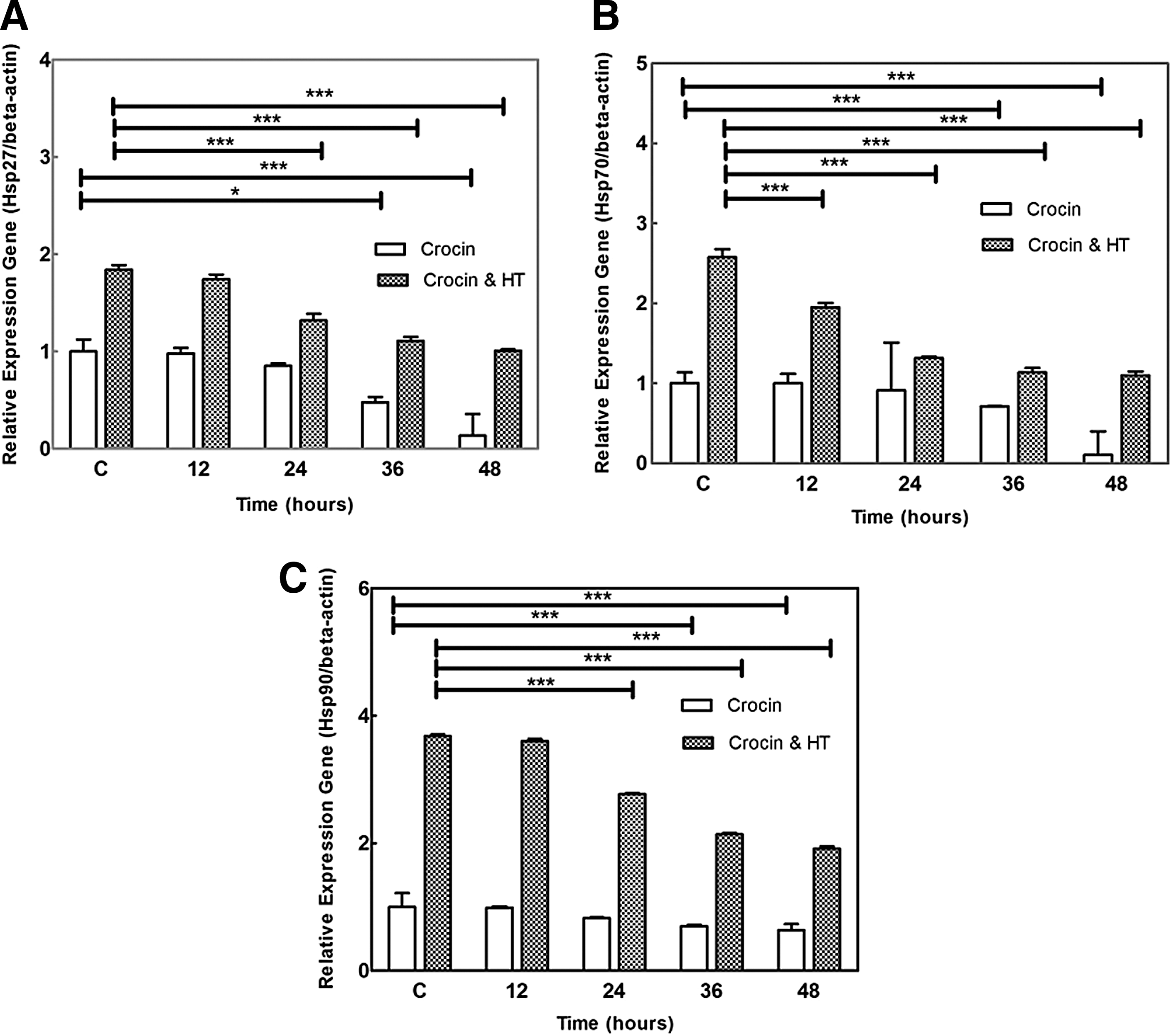
Combination effects of crocin (1.5 mg/mL) and HT (43°C for 2 h) on mRNA expression of
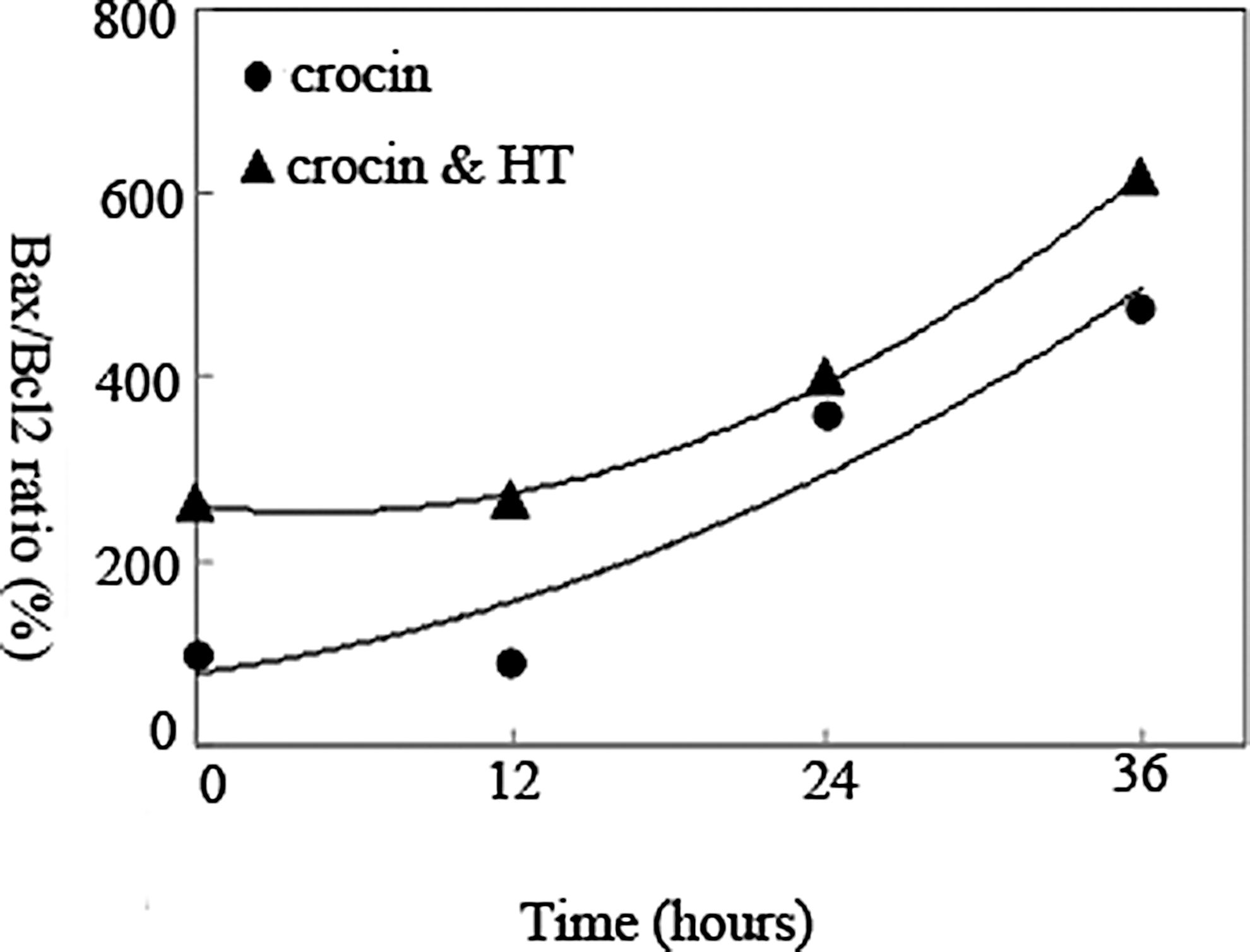
Combination effects of crocin (1.5 mg/mL) and HT (43°C for 2 h) on Bax/Bcl-2 ratio in MDA-MB-468 cells. Data represented relative gene expression (Target/β-actin).
ELISA analysis
Cell culture supernatants assayed with sandwich ELISA for Hsp70 and Hsp90 proteins. Results were consistent with our quantitative real-time polymerase chain reaction (qRT-PCR) data. Hsp70 and Hsp90 proteins in supernatants of HT-treated cells increased more than 1.8-fold compared with control nonheat. Crocin (1.5 mg/mL) significantly decreased the level of these HSPs in a time-dependent manner in both heat and nonheat groups (p < 0.001; Fig. 6A, B).
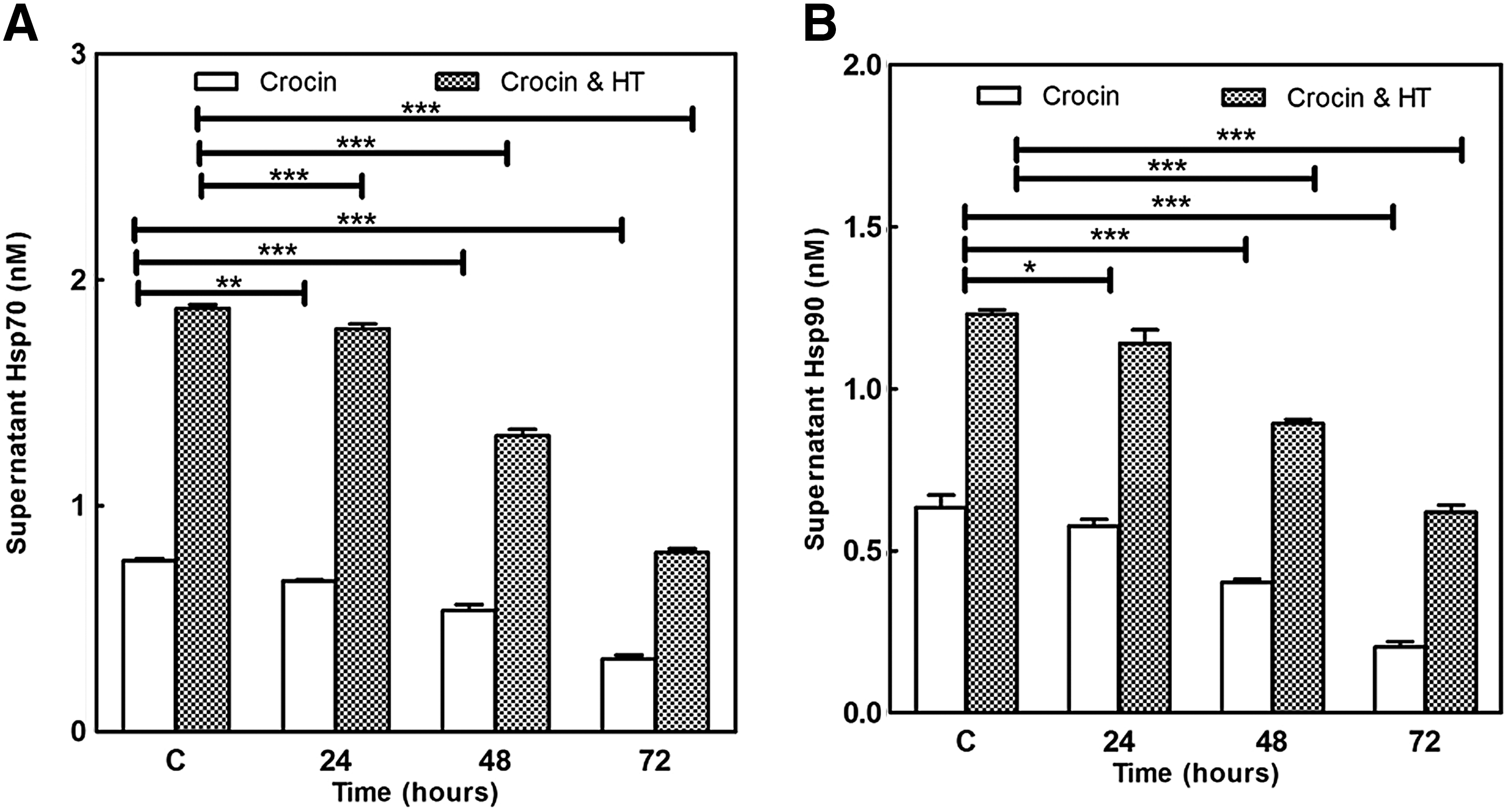
Combination effects of crocin (1.5 mg/mL) and HT (43°C for 2 h) on
Discussion
Breast cancer is the most common cancer among women with nearly 1.7 million new cases in 2012 worldwide (World Cancer Research Fund International, 2015). The prevalence of breast cancer is rising among Iranian women (Sepandi et al., 2014). Combined treatments have been examined in clinical trials to show benefits of adjuvant and neoadjuvant therapy for breast cancer. These strategies may greatly delay cancer growth and improve the patient's survival (Block et al., 2015; Vinay et al., 2015)
Crocin alone or combined with other botanical formulations exert an inhibitory effect on the proliferation of various cancer cells, including gastric, breast, prostate, etc. (Hoshyar et al., 2013; Festuccia et al., 2014; Bhandari, 2015). The main mechanisms underlying anticancer activities of crocin involve (1) interaction with DNA, histone H1, and H1-DNA complex (Ashrafi et al., 2005; Bathaie et al., 2007); (2) downregulation of catalytic subunit of human telomerase, hTERT (Noureini and Wink, 2012); and (3) cell cycle arrest and induced apoptosis (Hoshyar et al., 2013; Rezaee et al., 2013; Amin et al., 2015). Apoptosis is regulated by several mediator genes such as proapoptotic Bax and antiapoptotic Bcl2 and HSPs genes, which are important targets for cancer therapy (Sreedhar and Csermely, 2004).
On the other hand, several studies have demonstrated the antitumor property of HT on breast cancer (Iemwananonthachai et al., 2003; Oldenborg et al., 2016). HT is introduced as the fifth effective method of cancer therapy after surgery, chemo, radiation, and biological therapies (Liang et al., 2007).
To our knowledge, the cytotoxic effects of crocin in combination with HT have not been shown yet. In this study, we demonstrated that HT enhanced the antitumor effect of crocin through growth inhibition and apoptosis induction in breast cancer cells (MDA-Mb-468), whereas these changes were not observed in normal human breast epithelial cells (MCF10-A). Crocin induced morphological changes related to apoptosis such as blebbing and pyknotic nuclei in MDA-MB-468 cells, which is in agreement with other studies (Sun et al., 2011). It has been previously demonstrated that crocin suppressed the proliferation of cancer cells, while it did not have such cytotoxic effect on normal cells (Hoshyar et al., 2013). Figure 1A indicated that crocin markedly reduced MDA-MB-468 cell viability in a dose- and time-dependent manner (IC50 = 3, 2.5, and 2.1 mg/mL at 24, 48, and 72 h, respectively; Fig. 1C; p < 0.001). In this context, our data indicated that various concentrations of crocin did not inhibit the growth of normal cells (Fig. 1D). Besides that, crocin combined with HT significantly decreased the number of alive cancer cells (Fig. 1B) (IC50 = 2.5, 1.5, and 1 mg/mL at 24, 48, and 72 h, respectively; Fig. 1C; p < 0.001).
Crocin and its combination with HT notably decreased anchorage-independent growth of MDA-MB-468 cells with an inhibition rate of about 91–94% versus 94–95% by crocin alone 1.5 and 3 mg/mL, respectively (Fig. 2 and Table 2; p < 0.01). Abdullaev et al. (2003) previously showed the inhibitory effect of crocin on colony formation of cancer cells.
Several studies have proposed crocin as a proapoptotic component that induced changes in nuclear morphology (Bakshi et al., 2010; Li et al., 2013). We confirmed both these previous findings and found that crocin and HT induced apoptosis as determined by Hoechst staining of cancerous cells. These data indicated that HT enhanced crocin-induced apoptosis in a dose-dependent manner. Generally, the release of cytosolic LDH enzyme from damaged cells was increased. Our results showed that the percentage of LDH leakage in cancer cells after combined treatment of crocin and HT was significantly more than crocin alone (Fig. 3; p < 0.001).
HSPs are known as molecular chaperones that contribute to the folding of misfolded proteins. HSPs have cytoprotective effects that lead to cell survival under various stresses. They are related with key stress signaling and apoptotic molecules so that they inhibit apoptosis and promote cell survival, proliferation, or differentiation (Schwock et al., 2008). HT induces different changes in the cells; however, it is believed that protein denaturation and cell membrane injury are main effects of HT toxicity. The destabilized membrane may allow more drug uptake and chemosensitization. However, HT increases the expression of HSPs in cancer cells (Ahmed and Zaidi, 2013), and HSPs can repair denatured proteins and decrease hyperthermic apoptosis, developing thermotolerance of cells. Sekihara et al. (2013) showed that pifithrin increased sensitivity of prostate cancer cells to HT through inhibited Hsp70. Also, He et al. (2012) demonstrated that quercetin containing liposomes increased HT-induced apoptosis through downregulation of Hsp70 in colon cancer cells. Based on cell growth inhibitory activity (as shown in MTT results) and loss of membrane integrity (as shown in LDH release into the medium) of crocin and HT against MDA-MB-468 cells, we evaluated the effect of crocin on the expression of HSPs, which was significantly increased by HT. Our data showed that an effective dose of crocin (1.5 mg/mL) resulted in decreased levels of Hsp27, Hsp70, and Hsp90 mRNA (Fig. 4; p < 0.001) and supernatant Hsp70 and Hsp90 proteins (Fig. 6; p < 0.001). The levels of supernatant Hsp70 and Hsp90 were 0.75 ± 0.009 and 0.63 ± 0.03 nM in control cells, respectively. Crocin decreased the amount of these HSPs to 0.5 ± 0.02 and 0.4 ± 0.009 nM after 48 h treatment, respectively. Interestingly, both crocin and HT reduced the supernatant Hsp70 and Hsp90 in treated cancerous cells (from 1.87 ± 0.01 and 1.23 ± 0.01 to 1.31 ± 0.02 and 0.89 ± 0.01 at 48 h, respectively (p < 0.001).
The relative levels of Bcl-2 (antiapoptotic protein) and Bax (proapoptotic protein) modulate apoptosis in the cells. Liang et al. (2007) demonstrated that the HT downregulated the expression of Bcl-2 and upregulated the expression of Bax in colon cancer cell line. In addition, Hoshyar et al. (2013) revealed that crocin induced apoptosis in gastric cancer cells through high Bax/Bcl-2 ratio. Our data were not only in agreement with their results, but also illustrated that the combination of crocin and HT enhanced Bax/Bcl-2 mRNA ratio more than crocin in treated cells (Fig. 5).
Conclusion
In conclusion, we investigated the antitumor effects of crocin on HT-treated human breast cancer cells. Our findings suggested that HT may increase the uptake of crocin by breast cancer cells through altering membrane permeability. In addition, crocin increased the Bax/Bcl2 ratio and reduced the expression of antiapoptotic HSPs, which were induced by HT. Therefore, we found that crocin in combination with HT treatment effectively inhibited proliferation of human breast cancer. Future studies are needed to evaluate the molecular mechanisms of the anticancer effects of crocin alone or in combination with chemotherapy in both in vitro and in vivo mammary tumor models.
Footnotes
Acknowledgments
The authors would like to thank the personnel of Central Laboratory of Research and Technology at Birjand University of Medical Sciences. They would also like to thank Dr. Mohammad Reza Abedini (Birjand University of Medical Sciences, Birjand, Iran) for performing the Hoechst staining.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.