
Abstract
bZIP transcription factors play key roles in plant growth, development, and stress signaling. A bZIP gene BnbZIP2 (GenBank accession number: KP642148) was cloned from ramie. BnbZIP2 has a 1416 base pair open reading frame, encoding a 471 amino acid protein containing a characteristic bZIP domain and a leucine zipper. BnbZIP2 shares high sequence similarity with bZIP factors from other plants. The BnbZIP2 protein is localized to both nuclei and cytoplasm. Transcripts of BnbZIP2 were found in various tissues in ramie, with significantly higher levels in female and male flowers. Its expression was induced by drought, high salinity, and abscisic acid treatments. Analysis of the cis-elements in promoters of BnbZIP2 identified cis-acting elements involved in growth, developmental processes, and a variety of stress responses. Transgenic Arabidopsis plants’ overexpression of BnbZIP2 exhibited more sensitivity to drought and heavy metal Cd stress during seed germination, whereas more tolerance to high-salinity stress than the wild type during both seed germination and plant development. Thus, BnbZIP2 may act as a positive regulator in plants’ response to high-salinity stress and be an important candidate gene for molecular breeding of salt-tolerant plants.
Introduction
P
The induction of ABA/stress-inducible genes can be directly correlated with the presence of stress-related cis-acting elements in the promoter region. As specific motifs, cis-elements in the promoter region mediate the expression of gene in response to various biotic and abiotic stresses. These cis-acting regulatory elements include ABA-responsive elements ABRE and MYB element (Tak and Mhatre, 2013), dehydration-responsive elements (DRE) (van den Dries et al., 2011), salt-responsive elements GT-1 cis-element (Tak and Mhatre, 2013), ethylene-responsive element ERE (Castrillón-Arbeláez et al., 2012), and so on, which are involved in the regulation of dehydration, drought, salt, and signaling responsive gene expression (Liu et al., 2012; Tak and Mhatre, 2013).
Although bZIP genes are strongly induced by biotic and abiotic stress conditions, they are differentially regulated by various stresses and have been suggested to play diverse roles in stress response. Expression of SlAREB1 is induced by drought and salinity. The SlAREB1 overexpressing transgenic tomato plants show increased tolerance to salt and water stress (Orellana et al., 2010). Expression of Poncirus trifoliate PtrABF gene is induced by dehydration, low temperature, and ABA. Overexpression of PtrABF gene in transgenic tobacco plants enhances tolerance to dehydration shock, long-term water stress, and low temperature treatment (Huang et al., 2010). OsABF1 expression in rice seedling shoots and roots is found to be induced by anoxia, salinity, drought, oxidative stress, cold, ABA, and D-allose treatment (Hossain et al., 2010; Fukumoto et al., 2013). OsABF1 mutants Osabf1-1 and Osabf1-2 are more susceptible to high salinity and dehydration than wild-type rice plants, and the upregulated expression of some ABA/stress-regulated genes in response to ABA treatment is suppressed in the Osabf1 mutants (Hossain et al., 2010). Overexpression of OsABF1 shows sensitivity to D-allose in rice and inhibits the shoot growth of rice (Fukumoto et al., 2013). Expression of ZmbZIP60 is strongly induced by a wide spectrum of stresses, including dehydration, high salinity, ABA, and tunicamycin treatments (Wang et al., 2012). Overexpression of maize ZmbZIP60 in wild-type Arabidopsis displays enhanced bolting trends when subjected to dithiothreitol stress, and affects the expression levels of several key genes in the floral transition control, but no morphological phenotypes in atbzip60 plants are observed under various stresses (Wang et al., 2012). Expression of Arabidopsis AtbZIP24 is specifically induced by salt and hyperosmotic stress, not by cold stress. Overexpression of AtbZIP24 is suppressed in the salt-tolerant close relative Lobularia maritima, indicating that the protein functions as a negative regulator in salt stress acclimation (Yang et al., 2009). Nonetheless, under nonstress growth conditions, transgenic Arabidopsis lines with decreased AtbZIP24 expression activate the expression of stress-inducible genes involved in cytoplasmic ion homeostasis and osmotic adjustment (Yang et al., 2009).
Despite a number of bZIP members have been identified and comprehensively studied in plants, such as Arabidopsis (Jakoby et al., 2002; Yang et al., 2009; Lee et al., 2010), rice (Chen et al., 2012; Fukumoto et al., 2013), maize (Wang et al., 2012), potato (Muñiz García et al., 2012), soybean (Liao et al., 2008a; 2008b), tomato (Hsieh et al., 2010; Orellana et al., 2010), Vitis vinifera (Tak and Mhatre, 2013), P. trifoliate (Huang et al., 2010), information concerning perennial herbaceous plants is limited. Ramie (Boehmeria nivea L.) is a perennial herbaceous plant of the Urticaceae family. It is widely cultivated in China, India, other Southeast Asian, and Pacific Rim countries. Ramie fiber is mainly used for textile materials due to its fine characteristics. In addition, ramie plants have been found to possess a certain degree of As, Cd, Pb, and Zn tolerance (Yang et al., 2010), as well as a certain extent of drought and salt tolerance (Huang et al., 2013, 2014). Thus, it can be a good candidate for both fiber production and phytoremediation of sites contaminated by multimetals (Yang et al., 2010; Zhou et al., 2010). bZIP genes are important regulators of many stress conditions, there is a need to further characterize such bZIP genes in other plant systems to gain a better understanding of stress resistance mechanisms. The present study reports the identification, expression profiles, subcellular localization, promoter analysis, and overexpression of a bZIP transcription factor BnbZIP2 from ramie.
Materials and Methods
Plant materials, growth conditions, and stress treatments
The commercial ramie cultivar Xiangzhu-3 was planted in previous autumn as rhizomes in the Yunyuan experimental field, Hunan Agricultural University, Changsha, China. After a dormancy of 4 months, these rhizomes started regrowth in early March of 2011. Different tissues used in the study were collected from 30-day-old plants. Samples were harvested from the following tissues: leaves, roots, stems, and shoot tips, then mixed as a material pool, immediately frozen in liquid N2, and stored at −80°C for RNA extraction.
For tissue-specific assays, leaves, roots, stems, and shoot tips were individually collected, immediately frozen in liquid N2, and kept at −80°C until use for RNA isolation. Male and female flowers (1–3 mm) were individually sampled in autumn when ramie plants blossom, immediately frozen in liquid N2, and stored at −80°C until use for RNA extraction.
Thirty-day-old (about 10 cm tall) seedlings were treated with abiotic stress. Chemical treatments consisted of 20% polyethylene glycol (PEG) 6000, 250 mM NaCl, and 200 μM ABA. Seedlings in the fields were transferred to beakers containing fresh distilled water and kept for 10 days until fine roots were formed. These plants were divided into three groups. Each group had a similar distribution of plant types. For each group, 60 shoots were used. For drought, salt, and ABA treatment, 20% PEG 6000, 250 mM NaCl, and 200 μM ABA were added to the beakers and kept for 0, 2, 4, 6, and 8 h, then roots, leaves, stems, and shoot tips were individually sampled at different time points, immediately frozen in liquid N2, and stored at −80°C until RNA extraction.
Identification and bioinformatics analysis of BnbZIP2
Total RNA was extracted from mixed tissues of ramie using pBIOZOL Plant Total RNA Extraction Reagent (pBIOZOL reagent; Bio Flux Company) following the manufacturer's instruction, then was digested with DNase I (Fermentas). The quality of the isolated RNA was checked on 0.8% agarose gel and visualized under UV transillumination while the quantity of RNA was measured using a spectrophotometer. The DNase-treated RNA was reverse transcribed into cDNA using the First-Strand cDNA Synthesis Kit (Fermentas) according to the manufacturer's protocol. The newly synthesized cDNA was used as template for PCR amplification, with specific forward primer (5′ACGCGTCGACATGCAATCGTCATCTTCCTCTTC3′) and reverse primer (5′GGGGTACCTCATTGGTCCTGCCTAGGCCT3′) designed to amplify full-length open reading frame (ORF) of BnbZIP2. Primers for full-length cDNA were designed manually based on Unigene21903 identified in ramie transcriptome, using Primer Premier 5 (
The cDNA and protein sequences were analyzed using the BLASTn and BLASTp algorithm at NCBI database, respectively. Multiple alignments of the deduced amino acid sequence were performed using the ClustalW program. Theoretical isoelectric point (pI) and molecular weight were identified by tools in an internet server, ExPASy (Expert Protein Analysis System;
Expression profiles of BnbZIP2 under stress and nonstress conditions using RT-PCR analysis
Total RNA was isolated from different tissues grown under normal growth conditions and under drought, salinity, or ABA stress conditions. cDNA synthesis was performed using the same way as mentioned above. RT-PCR was performed to examine the tissue-specific expression of BnbZIP2 using forward primer 5′ TAAGCCAAATAGAGCCATTGACGG 3′ and reverse primer 5′ GATTATCAGCCTGTCTAACG-AAACC 3′. The ramie actin gene was amplified as an internal control with forward primer 5′ GCTCCGTTGAACCCTAAG 3′ and reverse primer 5′ GCTCCGATTGTGATGATTT 3′. RT-PCR was performed in an ABI 7300 Real-Time System (PE Applied Biosystems) according to the following parameters: 50°C for 2 min, 95°C for 10 min, 95°C for 30 s, 60°C for 60 s, and by 40 cycles. Each sample was amplified in three independent replicates. The PCR products were confirmed by sequencing of plasmid DNA constructs as described above. The relative expression levels were determined using the method of 2−ΔCT (Tak and Mhatre, 2013).
Isolation of BnbZIP2 promoter
Genomic DNA was isolated from collected leaves using TransGen reagent (TransGen Biotech Company) following the instruction provided by the manufacturer. DNA quality was examined on 0.8% agarose gel for purity. Thermal asymmetric interlaced (Tail) PCR was performed according to the methods of Thanh et al. (2012), using the genomic DNA as template and specific degenerate primers in an ABI 7300 Real-Time System (PE Applied Biosystems). Tail-PCR primers were designed manually using Primer Premier 5 (
Subcellular localization of EGFP fusion proteins
For subcellular localization studies, the BnbZIP2 full-length cDNA clone was isolated by RT-PCR using total RNA extracts from drought-treated roots and shoot tips of ramie seedlings with the following primers (The underlined are restriction site): forward primer containing a sal I site 5′ acgc
The sequence-confirmed fusion construct (pRI 101-BnbZIP2-EGFP) and control vector (pRI 101-EGFP) were individually introduced into Agrobacterium tumefaciens strain EHA105 following the method of Li et al. (2009). Transformation of onion epidermal cells was done according to the method of Sun et al. (2007). The transformed onion epidermal cells were cultured on sterile Murashige and Skoog (MS) medium (Phyto Technology Laboratories Company) for 16 h with a 16-h light–8-h dark photoperiod at 25°C, then EGFP was detected using a universal fluorescence microscope (Olympus).
Transformation of Arabidopsis and overexpression of transgenic plants
Generation of transgenic Arabidopsis plants
The construction of the overexpression vector was the same as that of subcellular localization studies, except for the use of the pRI 101-AN vector (TaKaRa). After verification by sequencing, the overexpression vector (pRI 101-BnbZIP2) and control vector (pRI 101-AN vector) were individually ligated into A. tumefaciens strain EHA105 following the method of Li et al. (2009).
Seeds of Arabidopsis (provided by Dr. Ying Lu, Hunan Agricultural University, Changsha, China) were sterilized for 20 s in 70% (v/v) ethanol, followed by rinse with sterile distilled water four times. These seeds were planted in multiple containers filled with a 5:5:1 mixture of vermiculite, black soil, and perlite, and grown in a growth chamber with a 16-h light at 23°C–8-h dark photoperiod at 16°C and 75% relative humidity, watered with nutrition solution. Transgenic Arabidopsis plants were generated according to the A. tumefaciens floral dip method (Clough and Bent, 1998). T1 seeds of treated plants and seeds of control plants were individually harvested.
Selection for transgenic plants using kanamycin
T1 seeds of transgenic Arabidopsis and wild type were sterilized for 5 min in 75% (v/v) ethanol, and incubated in 0.1% (v/v) HgCl2 for 5 min, followed by rinse with sterile distilled water five times. Afterward, the seeds were sown on MS agar plates. Kanamycin was used to screen for positive transgenic plants. Transgenic seeds were selected on selective MS medium (PhytoTechnology Laboratories Company) added with 30 μg/L kanamycin in a growth chamber with a 16-h light at 23°C–8-h dark photoperiod at 16°C. After selection for 14 days, the kanamycin-resistant shoots with green were transferred into the multiple containers, further screened for the transgene using PCR and specific primers. The seeds of these plants were collected and germinated to produce T2 seedlings.
Molecular analysis of the transgenic Arabidopsis plants
Genomic DNA was isolated from leaves of putative transgenic Arabidopsis lines and untransformed control Arabidopsis as described earlier. After examination on 0.8% agarose gel for purity, genomic DNA was used as template in the PCR reaction in an ABI 7300 Real-Time System (PE Applied Biosystems). Specific primers used for PCR amplification to CaMV35S, NOS, and BnbZIP2 were as follows: Arabidopsis-BnbZIP2 (forward primer: 5′ TAAGCCAAATAGAGCCATTGACGG 3′, reverse primer: 5′ AGCTTGTTGGCCATCAAGGTATAG 3′) and CaMV35S-BnbZIP2 (35S forward primer: 5′ GAAGGTGGCTCCTACAAATGC 3′, NOS reverse primer: 5′ CATCGCAAGACCGGCAAC 3′). The PCR conditions used were 94°C for 4 min, 94°C for 30 s, 60°C for 30 s, 72°C for 40 s by 32 cycles, and a final 7 min extension at 72°C. Each sample was amplified in three independent replicates. PCR products were analyzed on 1.2% (w/v) agarose gel and visualized under UV transillumination.
Germination test of the transgenic plants
For germination tests, T2 seeds of transgenic Arabidopsis and wild type were sterilized as mentioned above and germinated on plates containing MS medium (PhytoTechnology Laboratories Company). For drought stress evaluation, 0, 50, 100, 150, 200, or 250 mM Mannitol was added to the MS medium; for salt tolerance analysis, 0, 50, 100, 150, 200, or 250 mM NaCl was added to the MS medium; for heavy metal tolerance assay, 0, 50, 100, 150, 200, or 250 μM CdCl2 was added to the MS medium. Next, the same number of the transgenic seeds and wild-type control were individually placed on the MS medium plates, kept in a plant growth chamber with a 16-h light at 23°C–8-h dark photoperiod at 16°C for 16 days. The number of germinated seeds was counted every 12 h until most of the seeds had germinated. When seedlings were grown for 16 days, the length of shoot and root was recorded. The drought, salt, and heavy metal tolerance experiments were repeated three times. The rate of seed germination was calculated as: germination rate = number of germinated seeds/total seeds × 100. The seedling length was calculated as: seedling length = shoot length + root length.
Statistical analysis
Drought, salt, and heavy metal treatment of the transgenic plants and wild type (WT) were repeated three times. The results are shown as mean ± SE. Data were processed by SPSS 16.0. Comparisons were tested after analysis of variance (one-way ANOVA) using Newman–Keuls test. Different letters indicate statistical difference at p < 0.05.
Results
BnbZIP2 contains a characteristic bZIP domain
The full-length cDNA of BnbZIP2 gene was isolated from ramie tissues by RT-PCR using the gene-specific primers. The GenBank accession number of BnbZIP2 gene is KP642148. Sequencing and bioinformatics analysis showed that the cDNA is 1587 bp in length, contains a 1416 bp ORF, encodes a predicted polypeptide of 471 amino acids with a predicted molecular weight of 52.19 kDa and a theoretical pI of 7.73. The putative BnbZIP2 showed 73% similarity with Nicotiana tabacum (GenBank: AAY15214), 60% similarity with Arabidopsis thaliana (GenBank: CAC40649), 74% similarity with V. vinifera (GenBank: XP_02275147), and 77% similarity with Ricinus communis (GenBank: XP_002532393) (Fig. 1). The BnbZIP2 protein contains a typical bZIP basic region domain (from position 183–200) and a leucine zipper containing three leucine repeats (at positions 206, 213, and 220) that were spaced from each other by seven amino acids.

Multiple alignments of the deduced amino acid sequence of BnbZIP2 and Ricinus communis (XP_002532393), Nicotiana tabacum (AAY15214), Vitis vinifera (XP_02275147), and Arabidopsis thaliana (CAC40649). Black shaded backgrounds indicated that the amino acids were identical or similar to BnbZIP2, respectively. The basic region and the three heptad leucine repeats are shown by double line and arrows, respectively.
To evaluate the divergence of BnbZIP2 from other bZIPs, a phylogenetic tree was constructed using DNAstar 5.0 with the known classified Arabidopsis bZIP transcription factors (Corrêa et al., 2008; Nijhawan et al., 2008). This analysis indicated that BnbZIP2 was most closely related to AtbZIP20 and AtbZIP45 that belong to Group D of the bZIP family in Arabidopsis (Fig. 2).
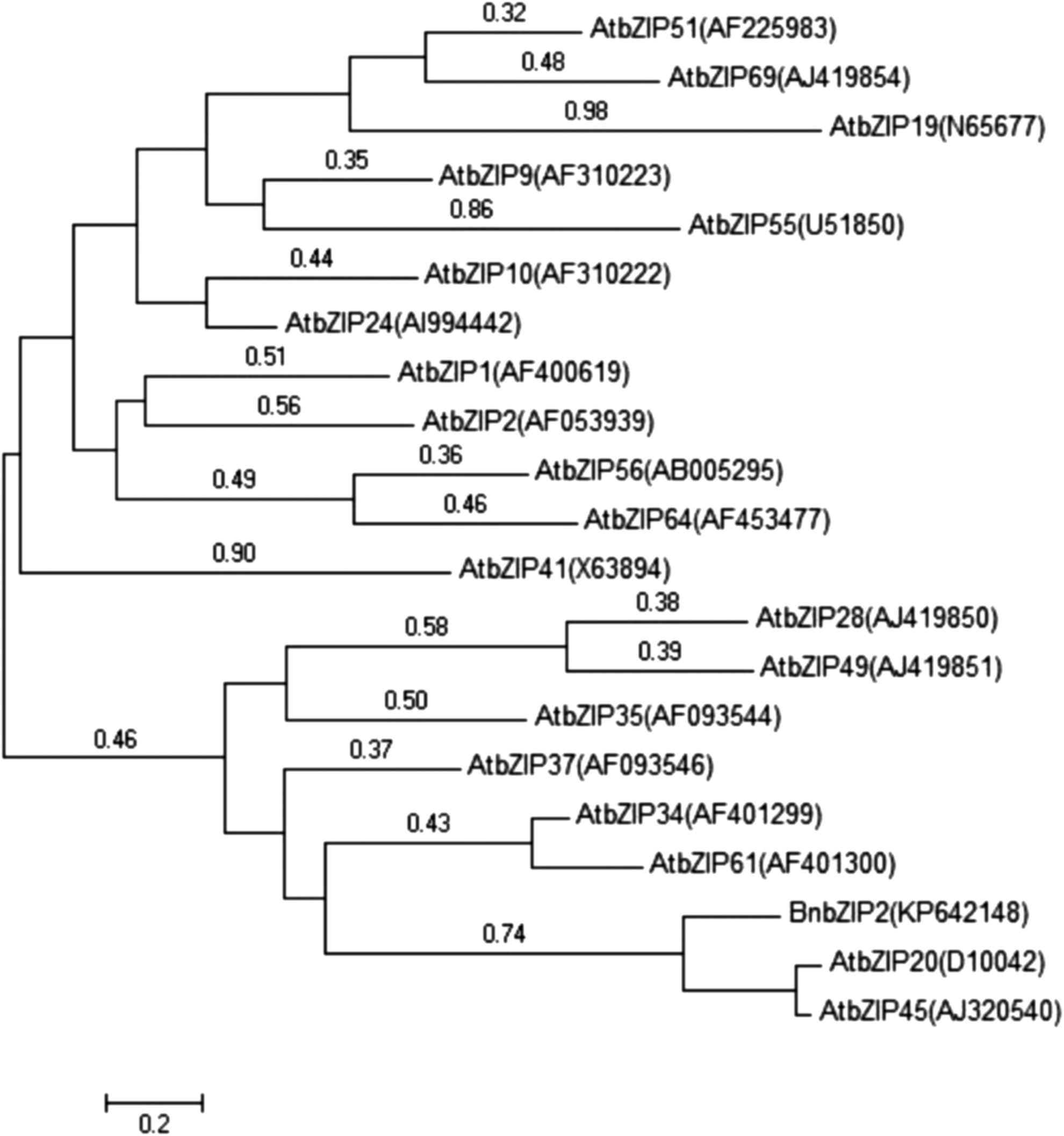
Phylogenetic tree of the bZIP subfamily members from Arabidopsis and ramie.
Subcellular localization of BnbZIP2
To determine the subcellular localization of the BnbZIP2 protein, the BnbZIP2 ORF was fused to EGFP in the pRI 101-EGFP vector. The fusion construct (pRI 101-BnbZIP2-EGFP) or the control (EGFP) was transiently expressed in onion epidermal cells through Agrobacterium infection. The EGFP fluorescence was observed in both nuclei and cytosol of pRI 101-BnbZIP2-EGFP-transformed onion epidermal cells (Fig. 3A, B), whereas in pRI 101-EGFP-transformed onion epidermal cells, the EGFP fluorescence was also seen throughout the whole cell (Fig. 3C, D), suggesting the protein is localized to nucleus and cytoplasm.
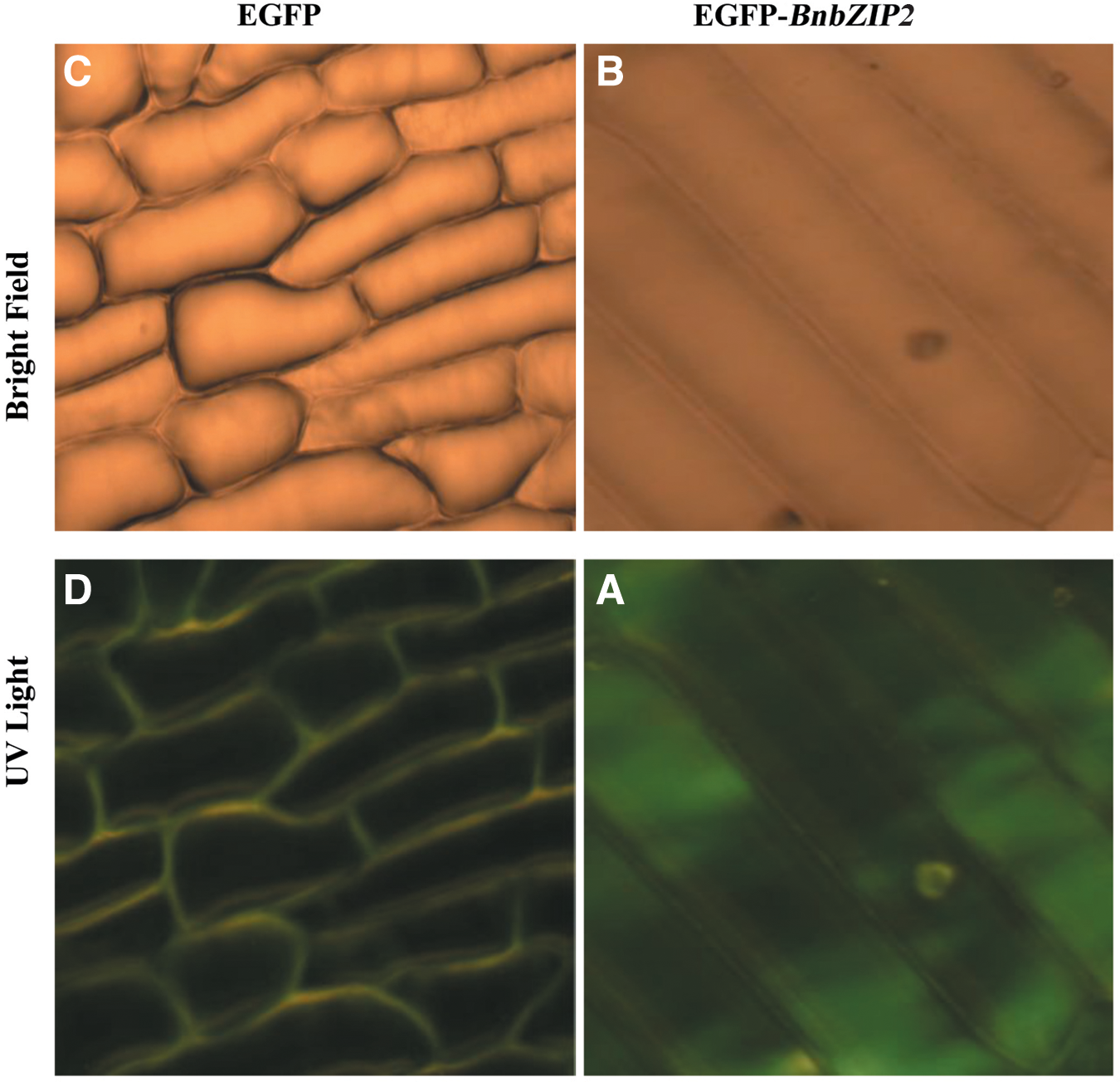
Nuclear localization of BnbZIP2 protein by EGFP fusion expression in onion epidermal cells through Agrobacterium infection. Fluorescence images
Expression of BnbZIP2 was detected under normal growth conditions and upregulated by drought, high salinity, and ABA treatment
Under normal growth conditions, the expression of BnbZIP2 was widely detected in different tissues like the root, leaf, stem, shoots tip, and female and male flower. The expression levels of BnbZIP2 greatly varied among tissues. The highest expression was observed in female flowers, which was more than 21 folds compared to that in root; followed by male flowers, which was more than 3 folds compared to that in root (Fig. 4A).
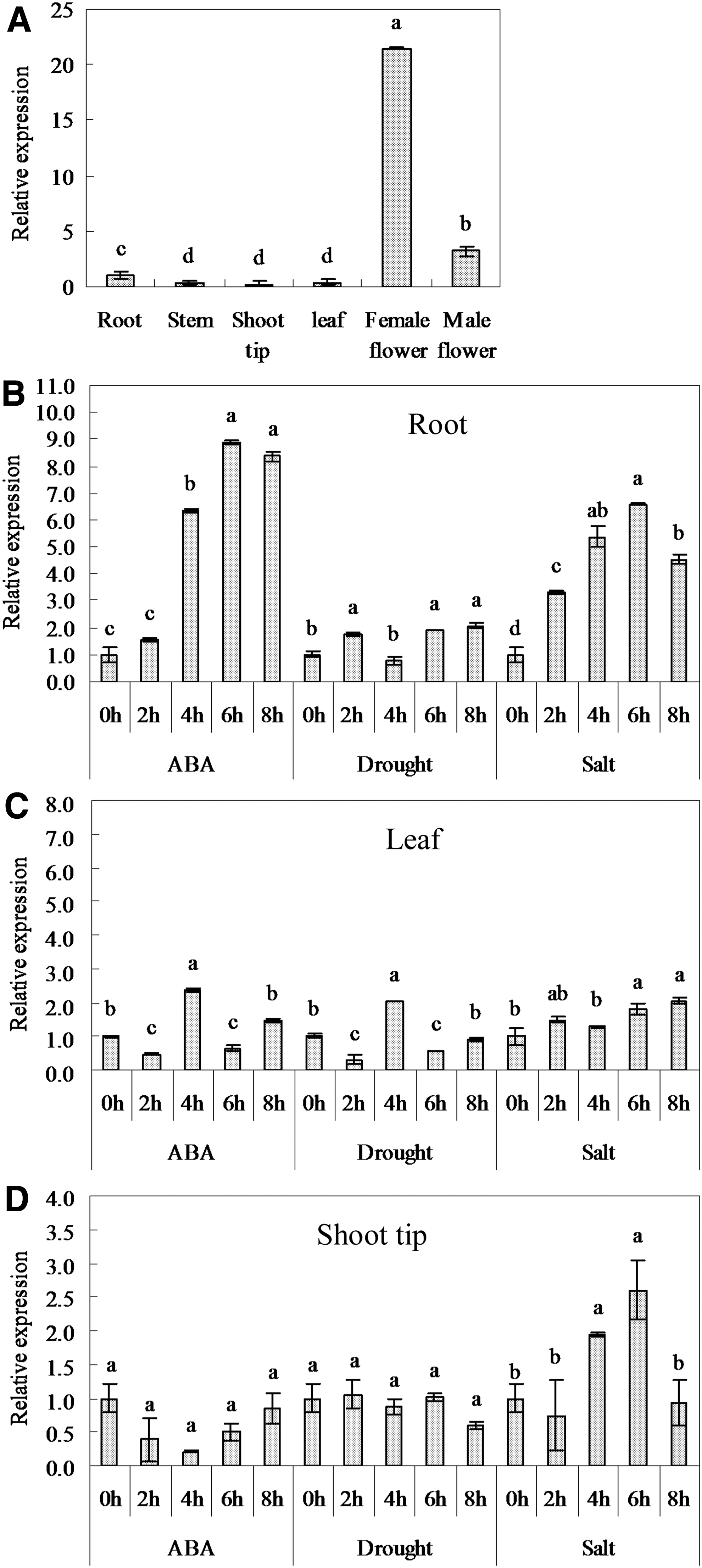
Expression profiles of the BnbZIP2 gene among ramie tissues under normal growth conditions
The expression pattern of the BnbZIP2 gene under various abiotic stresses was analyzed in root, shoot tip, and leaf tissues through RT-PCR (Fig. 4B–D). The transcription levels of BnbZIP2 in the roots was rapidly and strongly upregulated by ABA, high-salinity, and drought treatment (Fig. 4B). The expression of BnbZIP2 in the leaves was also noticeably enhanced in response to ABA, drought, and high-salinity treatment (Fig. 4C). The transcript levels of BnbZIP2 in the shoot tips significantly increased under high-salinity stress, whereas slightly changed under ABA and drought stress (Fig. 4D).
Putative cis-acting elements were found in the BnbZIP2 promoter
A 796 bp fragment upstream region of BnbZIP2 coding sequences was isolated using Tail-PCR and gene-specific primers from ramie genome sequence present in the public database NCBI. Computational analysis using PlantCare and PLACE databases revealed a number of putative cis-acting elements within the promoter region of BnbZIP2 (Fig. 5). Among them were three ATAT boxes at positions −499, −733, and −784 bp, four TATA boxes at positions −21, −535, −776, and −780 bp, whereas three CAAT boxes were located at positions −93, −109, and −607 bp. Other putative cis-acting elements were found within the promoter sequence, such as P-box at positions −366, −522, and −623, 5′-UTR Py-rich stretch at positions −276 and −469 bp, and TCA-element at position −391 bp (Fig. 5). The detailed function of putative cis-acting elements is in Table 1.
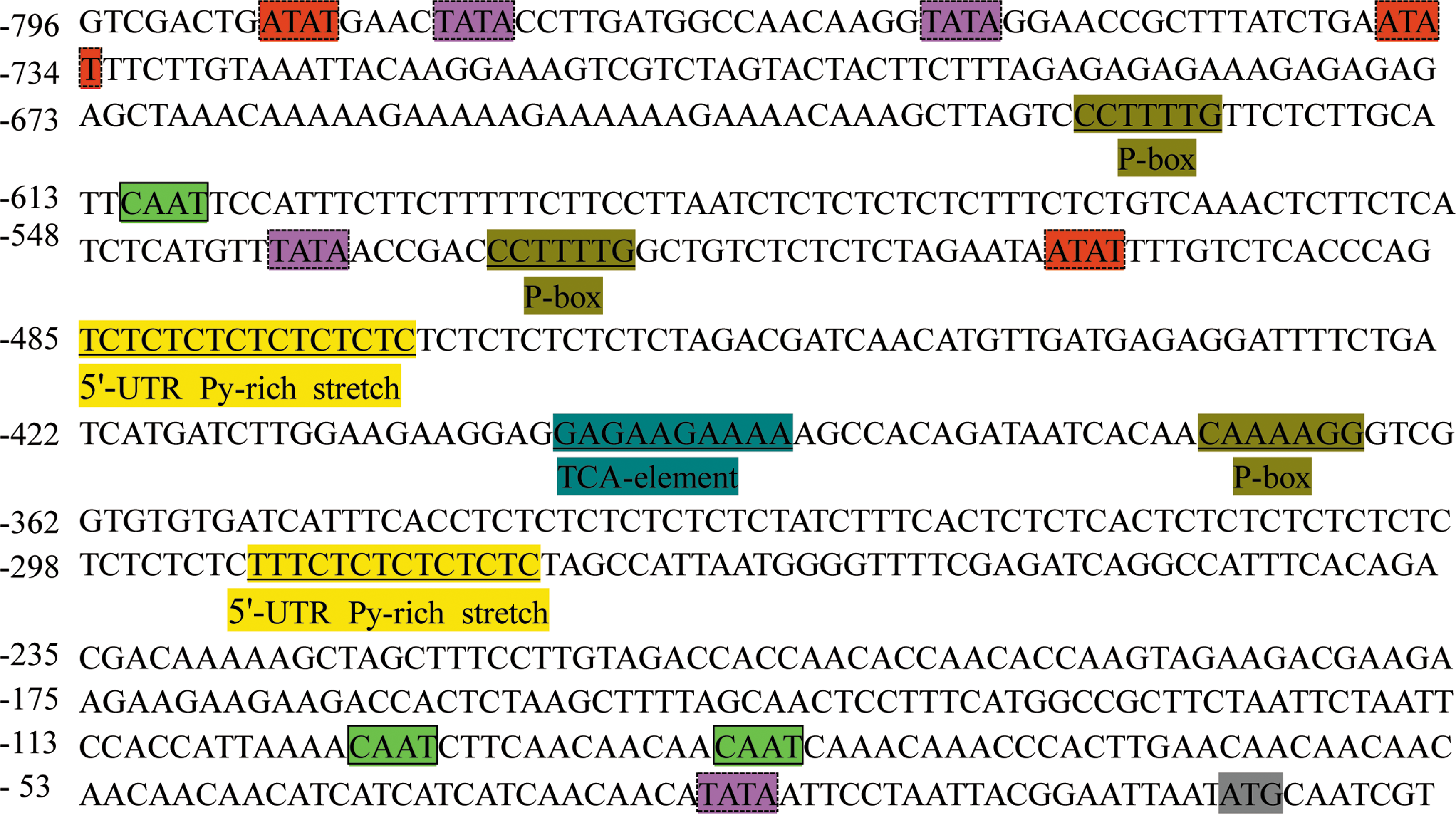
Nucleotide sequence of the BnbZIP2 promoter. Nucleotides were numbered on the left. The putative translation start sites, TATA box, CAAT box, and other important cis-regulatory elements are boxed and labeled. Color images available online at
The 5′ regulatory regions were analyzed using PlantCare and PLACE databases.
GA, gibberellins; SA, salicylic acid.
BnbZIP2 exhibits sensitivity to drought and metal stress, and tolerance to high-salinity stress
To examine the function of BnbZIP2 in plants, Agrobacterium-mediated transformation of transgenic Arabidopsis plants were generated using a binary vector containing BnbZIP2 under the control of CaMV 35S promoter. The kanamycin plate selection method was used to detect transformed progeny seedlings. Obvious differences in the phenotypes of the transgenic and WT seedlings were observed. The kanamycin-resistant transgenic plants were deep green and grew normally, whereas the kanamycin-sensitive wild seedlings exhibited a lack of greening, and the growth was inhibited (Supplementary Fig. S1; Supplementary Data are available online on
To investigate the effects of the overexpression of BnbZIP2 on drought, salt, and heavy metal tolerance, seeds of the BnbZIP2-overexpressed plants (T2 progeny) and WT were subjected to germination tests. When exposed to 0 (CK), 50, 100, 150, 200, or 250 mM Mannitol, a continuous decline in seed germination in both the transgenic Arabidopsis and WT was observed with the increasing of Mannitol concentration (Fig. 6A). Although seed germination showed no difference between the WT and transgenic Arabidopsis under control (nonmannitol), it was significantly lower in the transgenic lines than in WT when subjected to the remaining five Mannitol levels (Fig. 6A). Similar trend was also observed when seeds of the BnbZIP2-overexpressed Arabidopsis (T2 progeny) and WT were grown on MS agar plate with 0 (CK), 50, 100, 150, 200, or 250 μM CdCl2 (Fig. 6B). However, when subjected to 50, 100, 150, 200, or 250 mM NaCl, a considerable reduction in seed germination in the WT was observed at 50, 100, or 150 mM NaCl (Fig. 6C). When subjected to 200 mM NaCl, the WT did not germinate, whereas transgenic seed germination was 9.2%, respectively (Fig. 6C). When grown under 250 mM NaCl, neither the WT nor the transgenic seed germinated. With the increasing of NaCl levels, both the germination of the WT and transgenic seeds noticeably decreased, but the germination was always higher in the transgenic plants than in the WT (Fig. 6C).
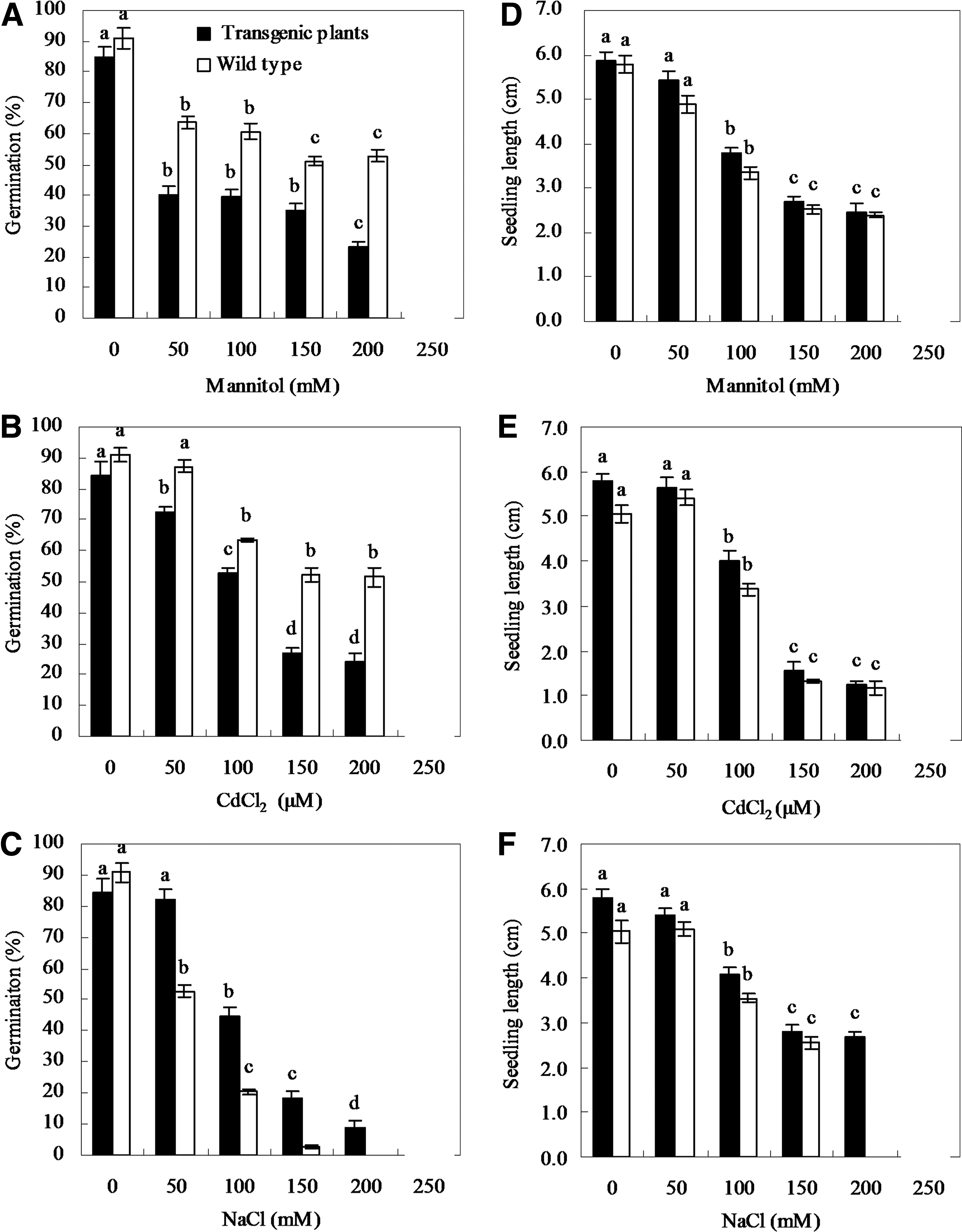
Seed germination and seedling length of WT and 35S-BnbZIP2 were grown on MS agar plates containing 0, 50, 100, 150, 200, and 250 mM Mannitol
With the increasing of Mannitol, CdCl2, and NaCl levels, a continuous decrease in the length of seedlings was detected in both transgenic plants and wild type (Fig. 6D–F). There was no significant difference in the length of seedlings between transgenic plants and wild type under dehydration and heavy metal stress. Similar trend was also found when exposed to 50, 100, 150 mM NaCl, but the transgenic seeds could germinate and grow at 150 mM NaCl (Fig. 6C, F). Interestingly, both transgenic seedlings and wild type became yellow under CdCl2 stress, whereas they were green under Mannitol or NaCl stress (Supplementary Fig. S2–S4). Shoots and roots appeared as same morphological phenotypes between transgenic plants and wild type under Mannitol, CdCl2, or low NaCl stress (Supplementary Fig. S2–S4).
Discussion
In this study, ramie BnbZIP2 was isolated and characterized. The BnbZIP2 gene contains a characteristic bZIP basic region domain in its N-terminal end, which is a typical protein structure for the group member of bZIP family (Corrêa et al., 2008). Amino acid sequence alignment demonstrated that BnbZIP2 is similar to other bZIP proteins, including N. tabacum, A. thaliana, V. vinifera, and R. communis (Fig. 1). The phylogenetic analysis indicated that BnbZIP2 is closely related to AtbZIP20 and AtbZIP45, which are classified as Arabidopsis bZIP group D subfamily (Jakoby et al., 2002). These findings suggest the possibility of functional conservation among these transcription factors. Subcellular localization analysis revealed that the protein was localized to both nuclei and cytoplasm, which is in agreement with that reported on bZIP transcription factor SmJLB1 (Voigt et al., 2013).
The expression of BnbZIP2 was widely detected in ramie tissues, with remarkably higher levels in flowers than in other tissues. The expression was more than sevenfold in female flowers in comparison with that in male flowers, suggesting that BnbZIP2 gene may participate in plant growth, flower development, or sexual development. Consistent with this, four TATA boxes that regulate many developmental processes, including flower, fruit, and root development (Castrillón-Arbeláez et al., 2012) were predicted in the promoter sequence of BnbZIP2 gene (Fig. 5). This is also supported by the existence of three P-boxes for gibberellin (GA)-responsive elements (Li et al., 2012). It is well known that the plant hormone GA regulates major aspects of plant growth and development. In addition, BnbZIP2 is strongly induced by drought and ABA, which is recognized by the presence of MYB cis-element (ATAT) for drought and ABA responsiveness (Fig. 5) (Abe et al., 2003; Castrillón-Arbeláez et al., 2012; Tak and Mhatre, 2013). ATAT element also acts as Opaque-2-like transcriptional activators that are involved in controlling the expression of distinct classes of endosperm genes, and SUC box that is required for sugar responsive gene. ATAT box is essential for anthocyanin and flavonoid production, and trichrome differentiation. It also regulates genes that are responsive to development and seed germination (Castrillón-Arbeláez et al., 2012).
Moreover, a typical salicylic acid (SA)-responsive element (TCA-element) (Pandey et al., 2012) was found in the promoter sequence of BnbZIP2 gene. SA-mediated signaling pathways are in response to various biotropic agents and leads to the establishment of systemic acquired resistance (Shah, 2003). When suffered from pathogen attack, SA induces the expression of pathogenesis-related genes throughout the plant (Jakoby et al., 2002). Furthermore, the promoter fragment harbors three CAAT boxes that generally exert a positive effect on transcription, and two 5′-UTR Py-rich stretch motifs that confer high transcription levels (Pandey et al., 2012). Whereas, analysis of the cis-elements in the promoter region of BnbZIP2 identified no salt stress cis-element, which showed that it needed other elements to respond to salt inducement. All of the above showed that BnbZIP2 gene was involved in plant development and biotic and abiotic stress responses. The Arabidopsis bZIP Group D, consisting of 10 bZIP members, plays important roles in plant defense against pathogens and development. Among them, two members are involved in developmental processes, and one of them, AtbZIP46/Perianthia, controls floral organ number in Arabidopsis (Chuang et al., 1999). Thus, BnbZIP2 may function in the same way as members of Arabidopsis bZIP Group D.
bZIP transcription factors play important regulatory roles in plant growth, development, and responses to abiotic and biotic stresses. Some of them, such as ABF2, AREB1, VvbZIP23, and PtrABF, etc., act as a positive regulator (Kim et al., 2004; Fujita et al., 2005; Huang et al., 2010; Tak and Mhatre, 2013), whereas others, including OsbZIP52/RISBZ5, OsbZIP79, and Foatf1, etc. function as a negative regulator (Liu et al., 2012; Qi et al., 2013; Miyamoto et al., 2015). In this study, overexpression of BnbZIP2 suppressed seed germination of transgenic Arabidopsis plants, whereas there was no evident morphological difference between the transgenic and WT lines in terms of their seedling length and plant development at various levels of Mannitol or CdCl2, indicated that overexpression of BnbZIP2 just affected seed germination, but did not affected the growing of plants under Mannitol or CdCl2 stress condition (Supplementary Figs. S2 and S3).
On the other hand, overexpression of BnbZIP2 improved seed germination and development of transgenic Arabidopsis plants under high-salinity stress (Supplementary Fig. S4). The BnbZIP2 overexpression lines displayed altered stress response. They exhibited increased sensitivity to dehydration and Cd stress, whereas enhanced tolerance to high salinity. These results suggested that BnbZIP2 plays a positive regulatory role in plants response to high-salinity stress, whereas a negative regulatory role in plants response to drought and heavy metal Cd stress. Such results are also found in other bZIP transcription factors. The Arabidopsis ABF3 and ABF4 transgenic plants exhibit salt hypersensitivity, but drought tolerance (Kang et al., 2002). The Arabidopsis DREB2C overexpression plants are dehydration sensitive, whereas freezing and heat tolerant (Lee et al., 2010). Transgenic rice plants overexpressing OsbZIP16 has stronger resistance to drought, but more sensitive to ABA than wild type plants (Chen et al., 2012). Transgenic Arabidopsis plants overexpressing GmbZIP132 reduce ABA sensitivity, increase water loss rate, whereas enhance tolerance to salt treatment during germination (Liao et al., 2008). These findings suggest that members of the bZIP family regulate complex transcriptional networks in mediating plant abiotic stresses.
In fact, negative regulators of abiotic stress are relatively rare. As a negative regulator of drought and heavy metal Cd stress, BnbZIP2 might have a value for research in the abiotic stress pathways. On the other hand, most bZIP genes are induced by salt stress, but only a few bZIP genes such as Arabidopsis ABF2, rice OsbZIP23, soybean GmbZIP44, GmbZIP62, GmbZIP78, GmbZIP132, and tomato SlAREB have been reported to positively regulate salt stress tolerance (Kim et al., 2004; Liao et al., 2008a, 2008b; Xiang et al., 2008; Hsieh et al., 2010; Orellana et al., 2010). Soil salinity severely affects plant growth and agricultural productivity. As a positive regulator of high-salinity stress, BnbZIP2 may be involved in the physiological pathways that aid ramie in adapting to salt-affected soil. This meets the essential condition for utilization in salinity-prone environments. Thus, BnbZIP2 has high potential usefulness in genetic improvement of salt stress tolerance.
Conclusion
This study showed that ramie BnbZIP2 gene belongs to a group of bZIP transcription factors. Expression of BnbZIP2 is induced by drought, high salinity, and ABA. Analysis of the cis-elements in promoters of BnbZIP2 identified cis-acting elements involved in growth, developmental processes, and a variety of stress responses. Overexpression of BnbZIP2 in Arabidopsis displayed complex expression patterns involved in plant abiotic stresses. Nonetheless, BnbZIP2 acts as a positive regulator in plants response to high-salinity stress and may be an important candidate gene for molecular breeding of salt-tolerant plants.
Footnotes
Acknowledgments
This research was supported by the project of National Natural Science Fund (31 371 704), the National “Twelfth 5-Year” Scientific and Technological Support project (2012BAD20B05-04), and Scientific and Technological project of Hunan (2010TP4004-1, 2010FJ2013, and 2012FJ4063).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.