
Abstract
Gastric cancer is one of the most common malignancy in the world. microRNAs (miRNAs) are naturally occurring noncoding RNA that control gene expression by targeting messenger RNA (mRNA) for post-transcriptional repression or cleavage. This study focused on a specific miRNA, miR-21, which was overexpressed in gastric cancer and examined the effects of miR-21 inhibitor on biological functions of gastric cancer cells and its possible mechanism. Gastric cancer cells MKN74 were treated with miR-21 inhibitor, negative control, and blank control. Cell proliferation, colony formation, migration, and invasion were assessed. Real-time PCR and western blot were applied to examine the expression of phosphatase and tens in homolog deleted on chromosome ten (PTEN)/PI3K/mTOR pathway molecules. miR-21 inhibitor markedly suppressed proliferation, migration, invasion, and colony formation of gastric cancer cells. Anti-miR-21 treatment also reduced the expression ratio of B cell lymphoma 2 (Bcl-2)/Bax. Furthermore, miR-21 inhibition was associated with increased expression of PTEN, which in turn decreased the ratios of S235/236, S240/244, and p-AK/AKT in gastric cancer cells. Inhibiting miR-21 modulates biological functions of gastric cancer cells via PTEN/PI3K/mTOR pathway and miR-21 inhibitor may provide a novel therapeutic strategy for gastric cancer.
Introduction
G
The discovery of microRNA (miRNAs) opened a new generation for understanding cancer (Zhang et al., 2012). miRNAs are small, highly conserved, noncoding RNAs (21–23 nucleotides). These small molecules mainly bind imperfectly to the 3′ untranslated region of target messenger RNAs (mRNAs) and negatively regulate gene expression post-transcriptionally by inhibiting translation and causing degradation of target mRNA (Lewis et al., 2005). As each single miRNA can potentially target hundreds of different genes, it has been demonstrated that miRNAs play an important role in many physiological processes such as differentiation, proliferation, apoptosis, and stress response (Ambros, 2003). Additionally, they are key regulators in many diseases including cardiovascular disease, cancer, and genetic disease (Mendell and Olson, 2012).
Among those well studied miRNAs, miR-21 overexpression is commonly observed in tumors arising from diverse tissue types, including gastric cancer (Volinia et al., 2006). Several significant miR-21 targets associated with malignancy have been identified by different groups: Phosphatase and tens in homolog deleted on chromosome ten (PTEN) (Meng et al., 2007), programmed cell death 4 protein (PDCD4) (Asangani et al., 2008), reversion-inducing-cysteine-rich protein with kazal motifs (RECK) (Gabriely et al., 2008), RECK, and B cell lymphoma 2 (Bcl-2) (Papagiannakopoulos et al., 2008).
Despite the massive ongoing research on miRNA in cancer, the possible roles of miR-21 in gastric cancer pathogenesis remain so far unknown. The relevant pathway of miR-21 in the development of gastric cancer and whether its antagonism could be used as a potential treatment for gastric cancer are considerably less clear. In this study, we aim to investigate the role of miR-21 and its target gene PTEN in gastric cancer cells proliferation, migration, and invasion and to determine the underlying signaling pathway.
Materials and Methods
MKN74 cell culture and transfection
The human gastric adenocarcinoma cell line MKN74 was purchased from American Type Culture Collection (ATCC, USA). Cells were grown in DMEM supplemented with 10% fetal bovine serum (FBS) and penicillin/streptomycin (Gibco, USA) in a 5% CO2 humidified incubator at 37°C. 2 × 105 cells per well were seeded onto 6-well plates (Corning, USA). After cell attachment for 6 h, cells were transfected with 12.5 μl of miR-21 inhibitor (Ji Ma Pharmaceutical Co., Ltd, Shanghai, China), negative control (RiboBio) or PBS using 8 μl Lipofectamine™ 2000 (Invitrogen) diluted in 250 μl DMEM without FBS, and the process was according to the manufacturer's instructions.
To better study the effects of miR-21 inhibitor on biological functions of gastric cancer cells and its possible mechanism. The miR-21 inhibitor, the negative control, and PBS group are referred to as inhibitor group (IN), negative control group (NC) and blank control group (BC), respectively. We used the approach which has been successfully used to inhibit miR-21 (Chan JA, 2005). The anti-miR-21 inhibitor is a sequence-specific and chemically modified oligonucleotide to specifically target and knockdown miR-21.
Cell proliferation
Cell Counting Kit-8 (CCK-8) assay was performed to detect cell proliferation. The cells at logarithmic growth phase were collected and used to prepare single cell suspensions. About 1 × 105/mL exponential-phase cells were digested with trypsin (Invitrogen) and seeded in 96-well tissue culture plates at a density of 1 × 104 cells per well. All groups were performed in quadruplicates. At 24, 48, 72, and 96 h after incubation respectively, 10 μL of CCK-8 (Dojindo) was added to each well, and plates were incubated at 37°C for another 1–4 h. The optical density (OD) was measured at 450 nm using a scanning multi-well spectrophotometer. Cell viability rate (%) was calculated as (OD/ODBC at 24h) × 100%.
Clonegenic assay
Clonogenic assays were performed as described previously (Franken et al., 2006) with slight modifications. Briefly, MKN74 cells treated under three conditions were plated in triplicates at 50 cells/well in a 24-well plate and cultured at 37°C until visible colonies formed. Culture media were removed and cells were washed twice with PBS. Cells were then fixed with 4% paraformaldehyde for 15 min at room temperature and stained with Crystal Violet solution for 10 min. The stain was decanted, and plates were carefully rinsed with distilled water until background staining of the wells was minimized. Finally, plates were air-dried and colonies with more than 50 cells were counted manually using an inverted microscope. Experiment was repeated three times.
Cell migration
Cell migration was measured using modified Boyden chambers consisting of Transwell (Corning) membrane filter inserts in 24-well tissue culture plates as previously described (Sieg et al., 1998). Exponentially growing cells were trypsinized and resuspended to the density of 5 × 104/mL in FBS-free DMEM. Two hundred microliters cell suspension was added into the upper compartment of the chamber while 600 mL of 10% FBS-DMEM culture media were added to the lower chamber. After 24 h of incubation at 37°C the nonmigrated cells on the upper surface of the membrane were removed by a cotton swab. Migrated cells were fixed in 4% paraformaldehyde for 15 min and stained with 0.1% Crystal Violet for 15 min. Cells migration values were determined by counting stained cells using a light microscope with × 100 magnification.
Each determination represents the average of five individual fields and error bars represent standard deviation (SD). Experiment was repeated three times.
Cell invasion
Cell migration experiments were performed using Corning™ BioCoat™ Matrigel™ Invasion Chamber (Corning). MKN74 cells at the concentration of 25 × 104/mL were added into the upper compartment of the chamber. The rest of the experimental procedure was as described for the cell migration assay.
RNA extraction and RT-qPCR
Total RNA were extracted by TRIzol Reagent (Invitrogen) according to the manufacturer's instruction. The quality of extracted RNAs was examined by measuring the absorbance at 260/280 nm on spectrophotometer. Quantification of miRNA and mRNA was performed using SYBR Green PCR kit (Qiagen) with 7500 Fast Real-Time PCR system (Applied Bioystems). The relative change between miR-21 and U6 small RNA was quantified. For the quantification of PTEN, Bcl-2, and Bax mRNA, b-actin was used as a housekeeping gene.
The primers for miR-21 are (F) 5′-CAAAGATCACTATCCCAATCATC 3′ and (R) 5′-GCG GTCTTTCTCAATCTAAGTC-3′. The primers for U6 are (F) 5′-CTCGCTTCGGCAGCACA-3′ and (R) 5′-CTGGTC CTGGTATGAAGAAT-3′. The primers for PTEN are (F) 5′-TCCACAAACAGAACAAGATG-3′ and (R) 5′-CTGGT CCTGGTATGAAGAAT-3′. The primers for Bax are (F) 5′-CAGGATGCGTCCACCAAGAA-3′ and (R) 5′-CGTGTC CACGTCAGCAATCA-3′. The primers for Bcl-2 are (F) 5′- AGCGTCAACAGGGAGATGTC-3′ and (R) 5′-TTCCAC AAAGGCATCCCAGC-3′. The primers for b-actin are (F) 5′-CTAAGTCATAGTCCGCCTAGAAGCA-3′ and (R) 5′-TGGCACCCAGCACAATGAA-3′. Experiment was repeated three times.
Western blot
Cell lysates were prepared by resuspending the cells in cell lysis buffer (Cell Signaling) with protease and phosphatase inhibitors (Cell Signaling). Cells were then subjected to sonication for 20 s on ice. Lysates were centrifuged at 10,000 g for 10 min to sediment the particulate material. The protein concentration of the supernatant was measured by BCA protein assay (Pierce). Forty micrograms of protein were subjected to electrophoresis with a 10% sodium dodecyl sulfate-polyacrylamide gel (Bio-Rad) and then transferred to a polyvinylidene fluoride membrane. The membranes were incubated with antibodies against PTEN, Bcl-2, Bax, cleaved poly ADP-ribose polymerase (PARP), caspase 3, p-AKT, AKT, S235/5, S240/4, and b-actin overnight at 4C after the addition of blocking buffer. Secondary antibody to IgG conjugated to horseradish peroxidase (Santa Cruz) was used.
The blots were then incubated with the ECL western blot detection system according to the manufacturer's instructions. Data presented are representative of three separate experiments.
Statistical analysis
Data were expressed as the mean value ± SD. One-way ANOVA was performed by using statistical software SPSS19. Differences were considered statistically significant at p < 0.05.
Results
Inhibition of miR-21 decreases gastric cancer cells proliferation
Gastric adenocarcinoma cell line MKN74 was used to test whether miR-21 expression could modulate cell proliferation. Cells were transfected with miR-21 inhibitor (IN), negative control (NC), or PBS (blank control, BC). As shown in Figure 1, viability of MKN74 cells in IN group was significantly lower than blank control group (BC) at 48 h time point (*p < 0.05 compared with BC group, # p < 0.05 compared with NC group) while no difference was observed in NC group compared to BC. Time course analysis revealed further inhibiting effect of anti-miR-21 at later time points, 72 h and 96 h (*p < 0.05 compared with BC group, # p < 0.05 compared with NC group).
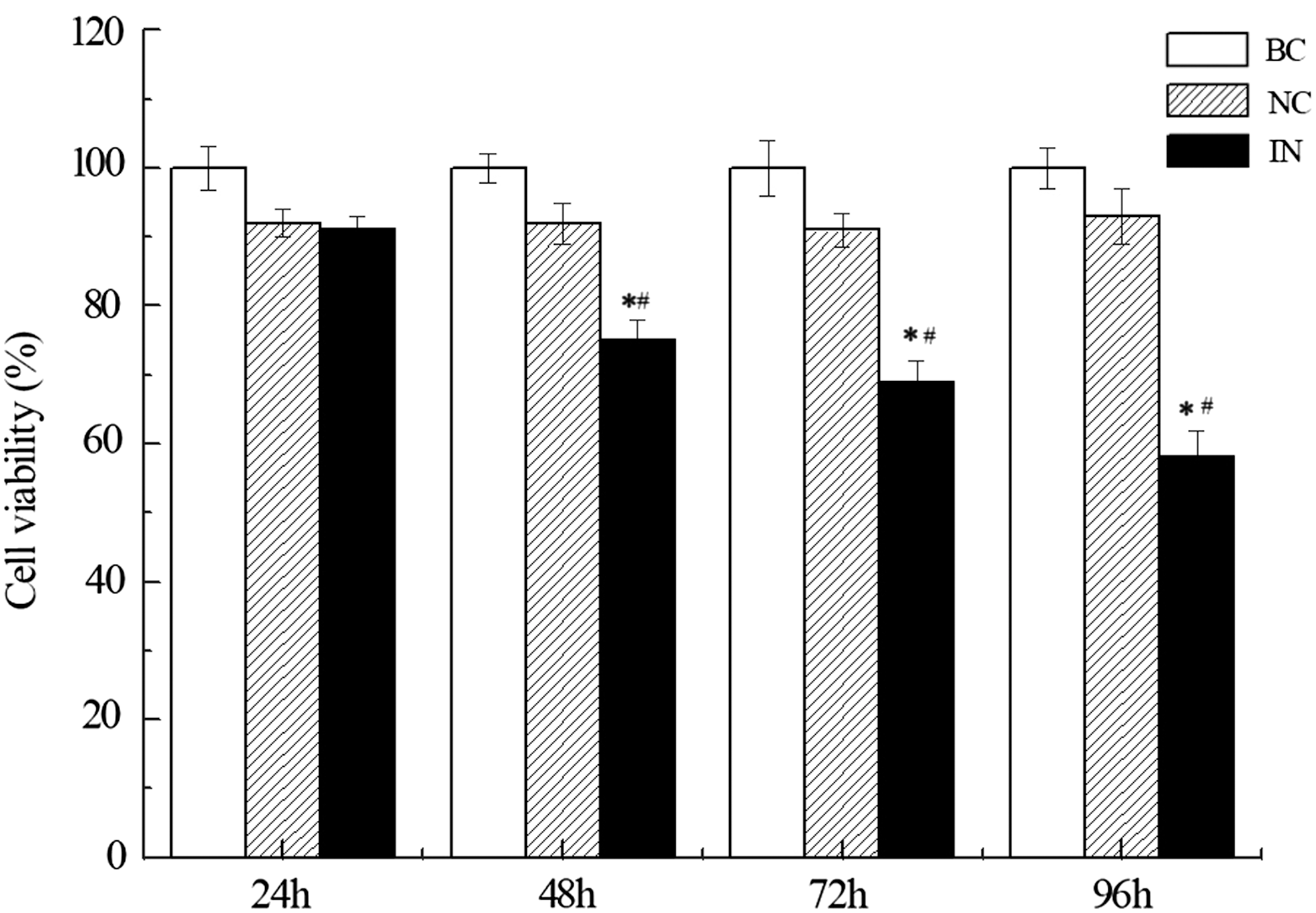
miR-21 inhibitors regulate gastric cancer cell proliferation. Cell proliferation was shown after transfection with miR-21 inhibitor (IN), negative control (NC), or PBS (blank control, BC) in MKN74 cells by using Cell Counting Kit-8. Cell viability rate (%) was calculated as (OD/ODBC at 24h) × 100% compared to blank control group at 24 h and it was assessed at 24, 48, 72, and 96 h. n = 4, *p < 0.05 compared with BC group, # p < 0.05 compared with NC group. OD, optical density.
miR-21 is essential for clonogenic ability of gastric cancer cells
Clonogenic assay or colony formation assay is an in vitro cell survival assay based on the ability of a single cell to grow into a colony, which is defined to consist of at least 50 cells. The assay essentially tests every cell in the population for its ability to undergo “unlimited” division. Anti-miR-21 was found to suppress colony formation of hepatocellular carcinoma cells. We applied this assay to investigate the role of miR-21 in clonogenic ability of gastric cancer cells. Consistent with findings in hepatocellular carcinoma cells, miR-21 inhibitor profoundly suppressed the colony formation ability of gastric cancer cells MKN74 compared to negative control and blank control groups (Fig. 2, *p < 0.05 compared with BC group, # p < 0.05 compared with NC group). It indicates that miR-21 is essential for clonogenic ability of gastric cancer cells.
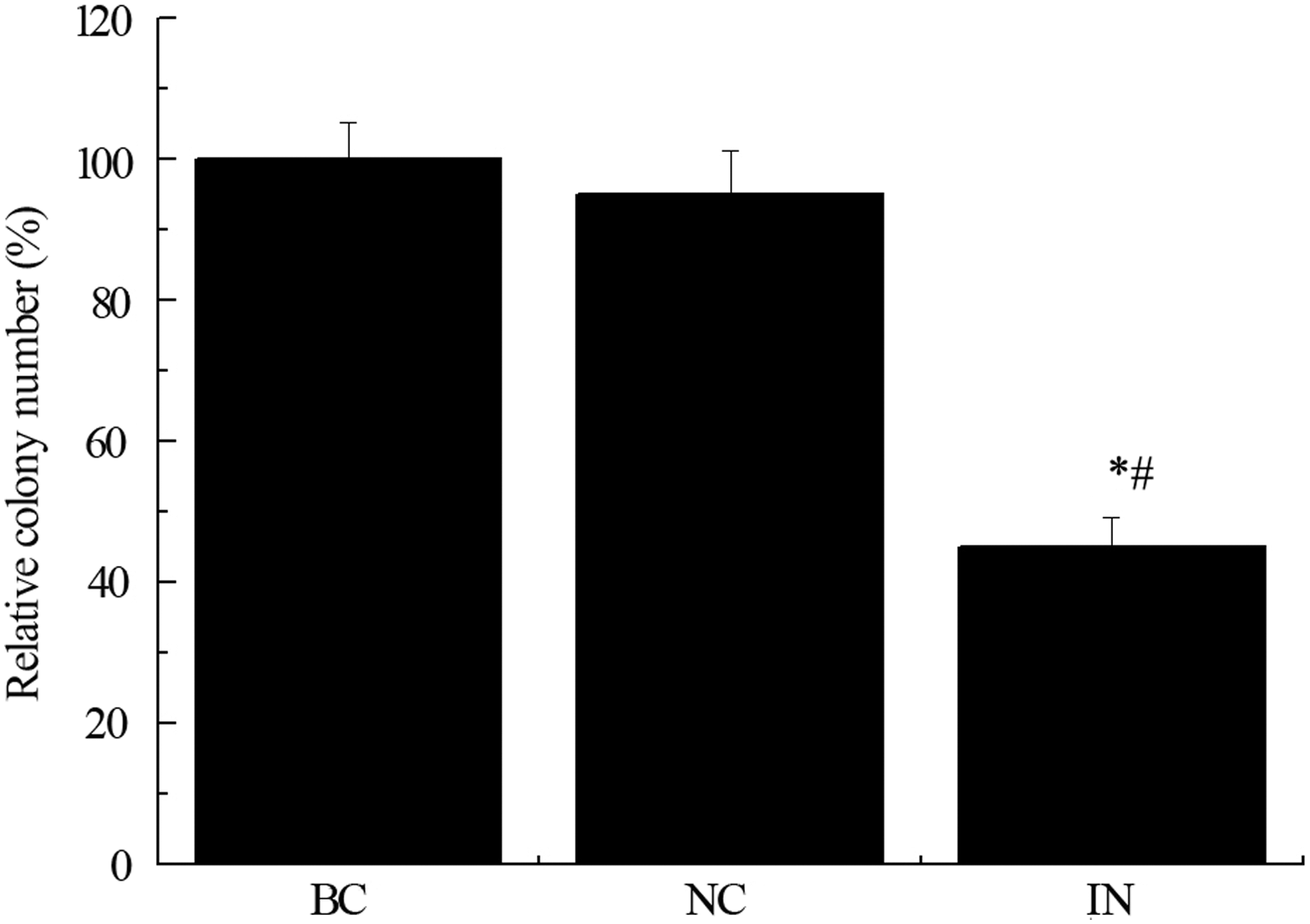
Effect of anti-miR-21 on colony formation of MKN74. MKN74 cells treated with miR-21 inhibitor (IN), negative control (NC), or PBS (blank control, BC) were plated at concentration of 500 cells/well in 24-well plates. Clonogenic assay showed IN group had significant reduced colony formation ability compared to BC and NC groups. n = 3 independent experiments, *p < 0.05 compared with BC group, # p < 0.05 compared with NC group.
Inhibition of miR-21 suppresses migration and invasion of MKN74 gastric cancer cells
We investigated the effect of miR-21 on cell migration and invasion of MKN74 gastric cancer cells. Migration of MNK74 cells was significantly suppressed by exposure of cells of miR-21 inhibitor (Fig. 3B, *p < 0.05 compared with BC group, # p < 0.05 compared with NC group). To examine the effect of anti-miR-21 on the invasive activity of MKN74 cells, we loaded the cells on Matrigel in the upper chamber of Transwell chambers. After incubation for 15 h, the cells that had invaded the lower chamber were counted. Negative control treated cells had no change in invasion compared to blank control group while the invasion capacity was significantly suppressed by miR-21 inhibitor (Fig. 3A, *p < 0.05 compared with BC group, # p < 0.05 compared with NC group).
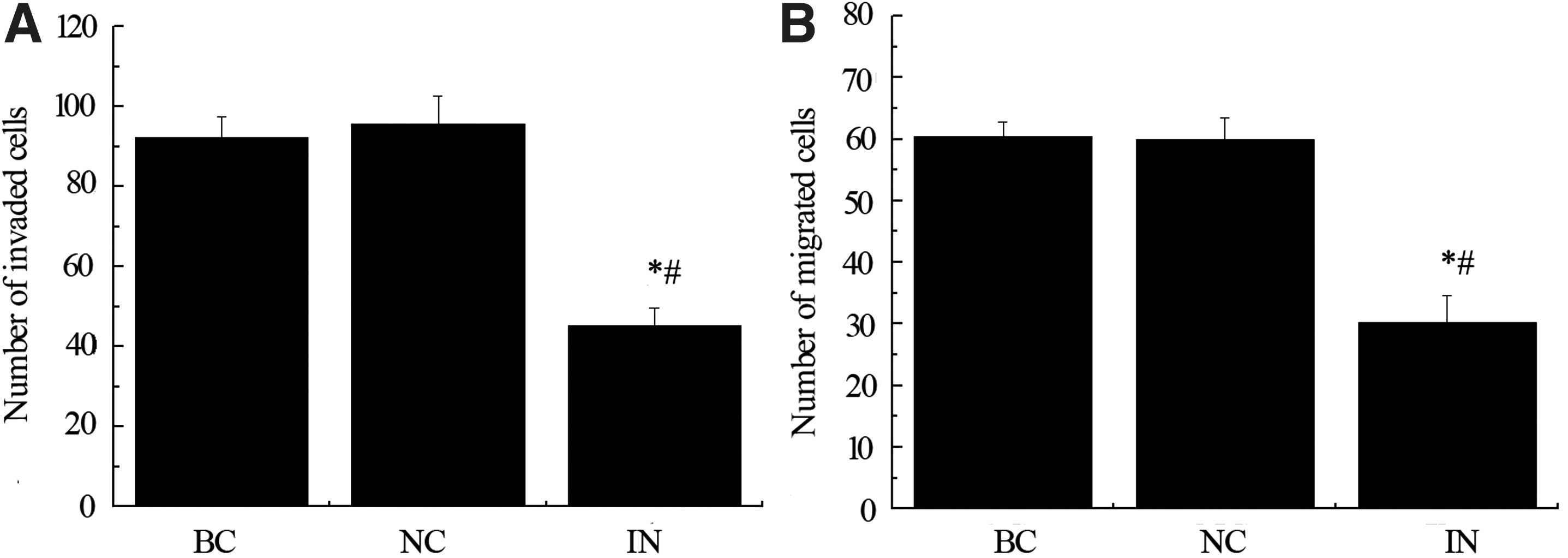
Inhibition of miR-21 suppresses migration and invasion of MKN74 gastric cancer cells. The effect of miR-21 on cell invasion and migration of MKN74 gastric cancer was tested by using Transwell chamber system with or without Matrigel. Migration of MNK74 cells was significantly suppressed by exposure of cells of miR-21 inhibitor. After 15 h incubation time, the cells that had invaded the lower chamber in negative control group had no difference to the number of invaded cells in blank control group
Apoptosis related genes are regulated by miR-21 inhibitor
Knock down of miR-21 in cultured glioblastoma cells triggered activation of caspases and lead to increased apoptotic cell death. To dissect the molecular basis underlying anti-miR-21 associated alteration of tumor growth, we examined expression levels of apoptosis related genes by qPCR and found that Bcl-2 mRNA was downregulated in the miR-21 inhibitor group (Fig. 4A; *p < 0.05 compared with BC group, # p < 0.05 compared with NC group). In contrast, the proapoptotic molecule Bax expression was upregulated with the same treatment compared to negative control and blank control groups (Fig. 4B; *p < 0.05 compared with BC group, # p < 0.05 compared with NC group).
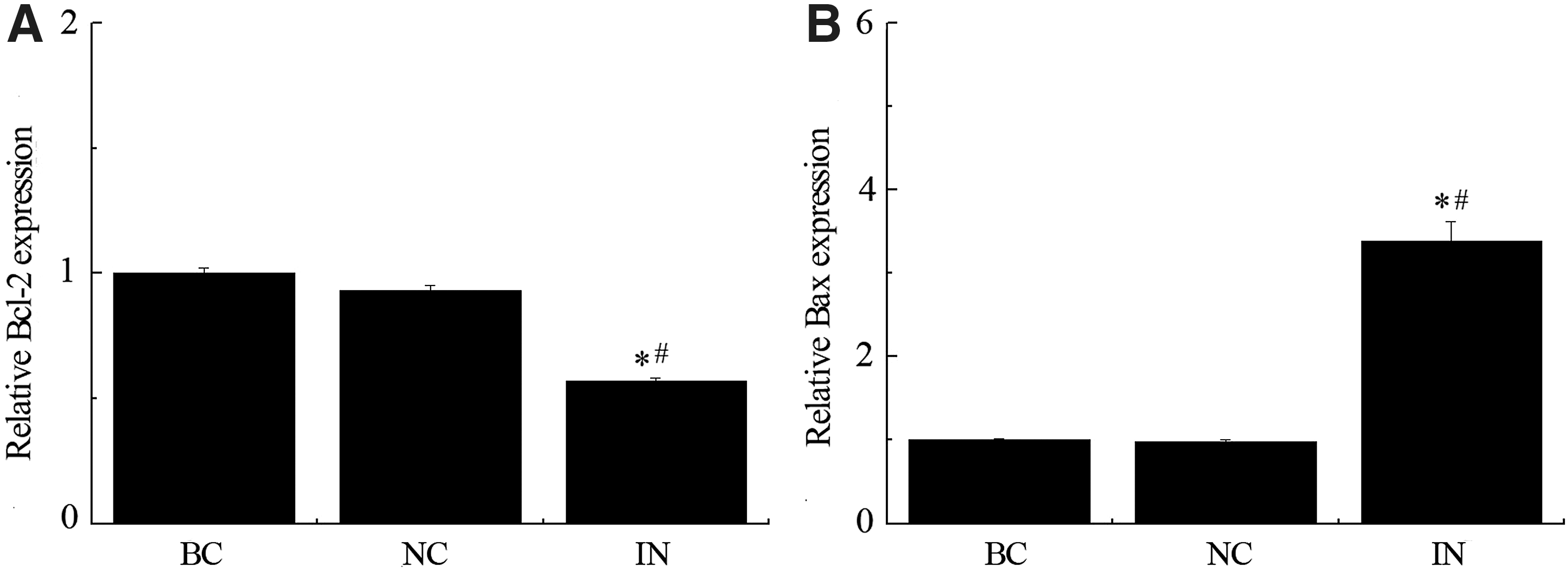
Apoptosis related genes are regulated by miR-21 inhibitor. Expression level of apoptosis related genes was carried out by qPCR. Bcl-2 mRNA was downregulated in the miR-21 inhibitor group
We next performed western blot analysis to determine the effects of miR-21 inhibitor on the protein levels of Bcl-2, Bax, PARP, and caspase 3 (Fig. 5). We found that Bcl-2 and caspase 3 proteins in gastric cancer cells were reduced by the treatment of miR-21 inhibitor and Bax and PARP proteins level were induced at the same time. Using the relative density ratio of Bcl-2/Bax, it clearly showed that inhibition of miR-21 significantly induced proapoptotic molecule while it reduced the antiapoptotic molecule.
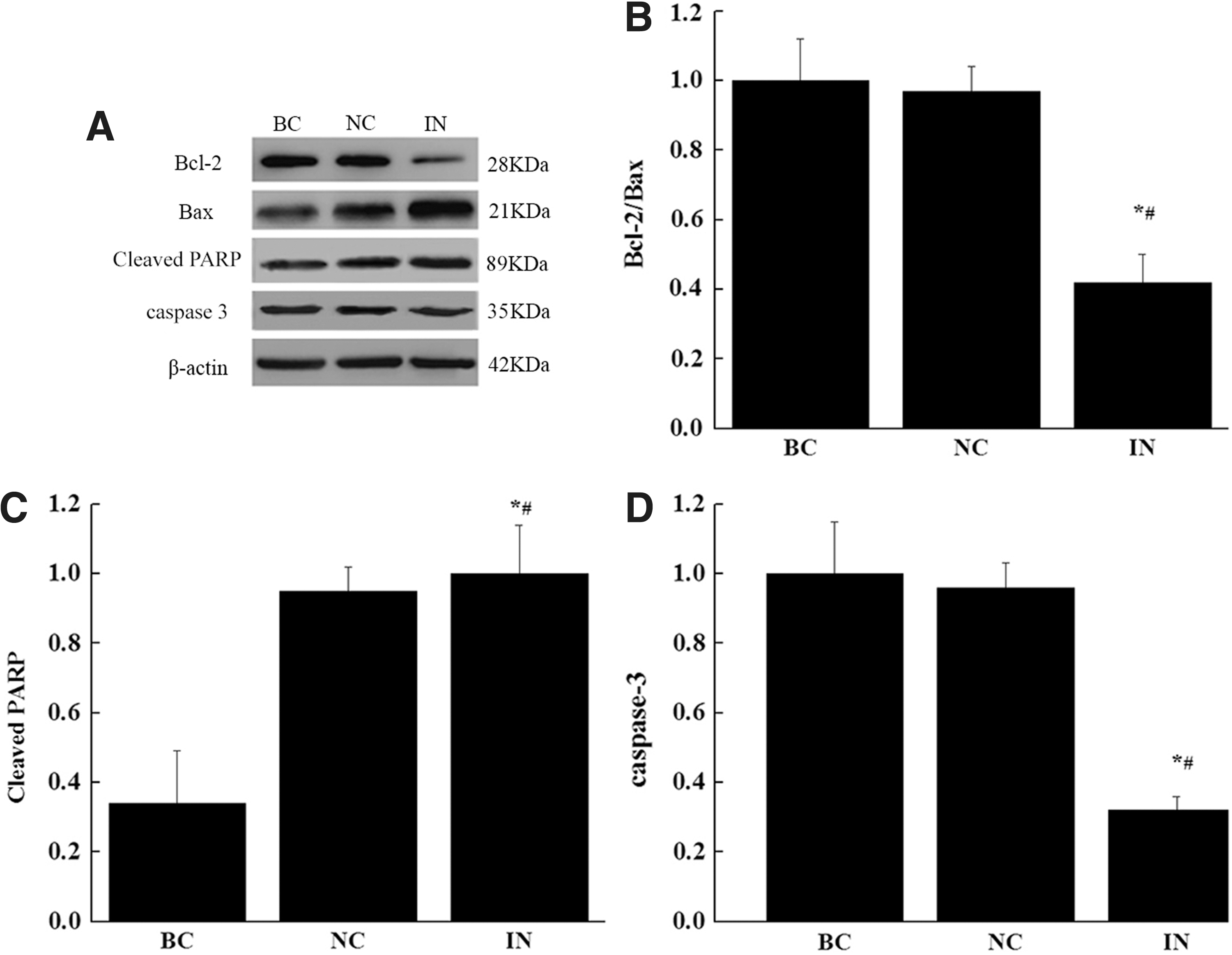
Effect of anti-miR-21 on the protein levels of Bcl-2, Bax, PARP, and caspase 3
miR-21 regulates expression of PTEN protein and PI3K/AKT/mTOR pathway
As expected, the relative expression level of miR-21 in miR-21 inhibitor group was reduced to about 50% compared to negative control and blank control groups (*p < 0.05 compared with BC group, # p < 0.05 compared with NC group). Interestingly, the mRNA level of PTEN in the same group (IN) was significantly higher than BC and NC groups (*p < 0.05 compared with BC group, # p < 0.05 compared with NC group).
Consistent with the findings in colorectal cancer cells (Xiong et al., 2013), an inverse association of PTEN and miR-21 expression was observed in our study of gastric cancer cells (Fig. 6).
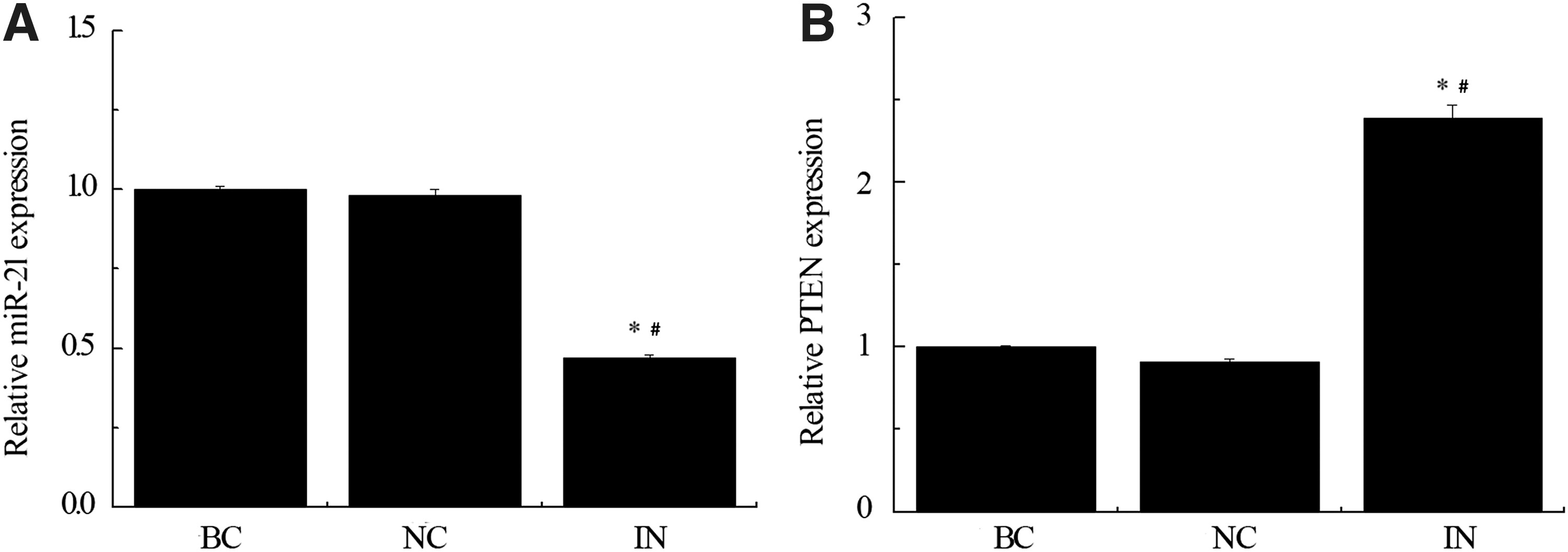
Inhibition of miR-21 upregulates expression of PTEN. miR-21
To gain insight into the underlying mechanisms responsible for the altered cellular behavior observed in miR-21 inhibitor-treated gastric cancer cells, we investigated potential associations with several cancer-related signaling molecules. We assessed whether the downregulation of miR-21 affected the protein expression of PTEN, p-AKT, AKT, S235/236, and S240/244 by western blot in MKN74 cell lines, which were transiently transfected with miR-21 inhibitor, negative control, or blank control (Fig. 7). Inhibition of miR-21 significantly increased PTEN protein leve, which was consistent with the upregulation of PTEN mRNA level. The ratio of S235/236, S240/244, and p-AK/AKT in miR-21 inhibitor group was significantly lower than the negative control and blank control groups.
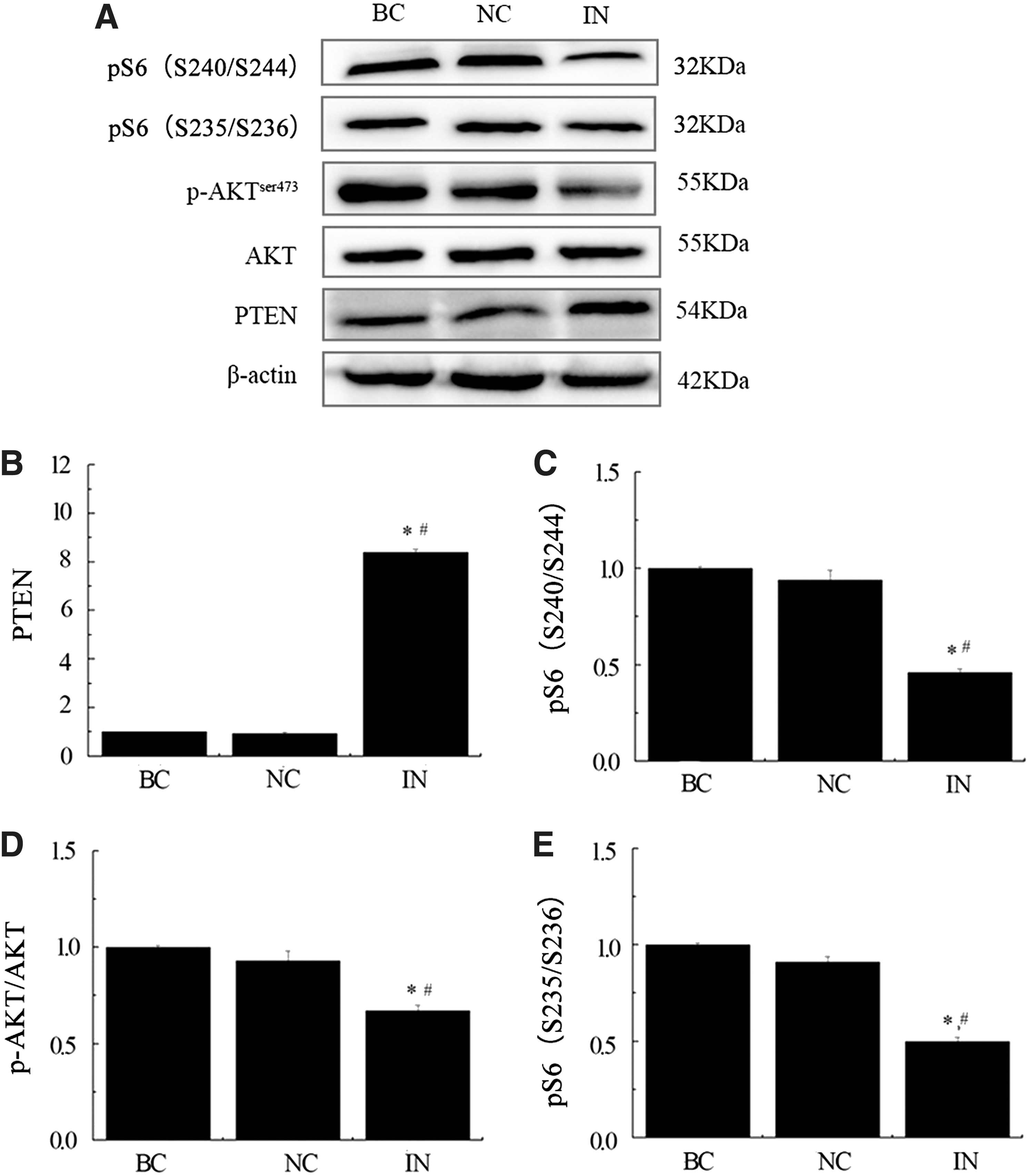
miR-21 inhibitor modulates PTEN protein and PI3K/Akt/mTOR pathway. Western blot analysis of protein levels of PTEN, p-AKT, AKT, S235/236, and S240/244 were representative data of three separate experiments
In summary, these findings showed that inhibition of miR-21 induced PTEN protein production, which in turn inhibited p-PI3K and activation of its downstream targets.
Discussion
As a new gene regulation mechanism, miRNAs have been the object of extensive research in recent years. Vast majority of people suffered from gastric tumorigenesis (Ying, 2016). In the search of miRNAs potentially involved in gastric tumorigenesis, Petrocca et al. (2008) found that miR-21 showed the highest overexpression in primary gastric tumors compared to nontumor controls by paired significance analysis of microarrays, with 4.5 median fold changes. This result was consistent with another large-scale miRNA profiling from six solid tumors, which showed that miR-21 was overexpressed in all six types of human malignancies including gastric cancer.
Studies on murine tumor models also found that conditional expression of miR-21 could promote lymphomagenesis in mice while silencing of miR-21 lead to disease regression, in part, by promoting apoptosis (Sun et al., 2016). Similarly, research on human glioblastoma showed that the use of miRNA inhibitors directed against miR-21 can inhibit the proliferation of glioblastoma cells that overexpressed miR-21. A systematic review and meta-analysis recently indicated that miR-21 overexpression was associated with decreased overall survival in patients with a wide variety of tumors and miR-21 expression was a potentially useful biomarker for predicting prognosis (Wang et al., 2014).
Given the important role of this oncomir in tumorigenesis, our current study tested the therapeutic potential of miRNA inhibitor for gastric cancer. We focused on several fundamental characteristics of neoplastic cells and found that anti-miR-21 inhibited MNK74 gastric adenocarcinoma proliferation, colony forming, migration, and invasion. miR-21 inhibitor also promoted apoptosis by inhibiting antiapoptotic molecule Bcl-2 and caspase 3 expression and inducing gene and protein expression of proapoptotic molecule Bax and PARP. These findings further qualify miR-21 as a potential target in gastric cancer treatment.
Next, we investigated the underlying signaling pathway through which miR-21 inhibitor affected the function of gastric cancer cells. PI3K/AKT/mTOR pathway is a prototypic survival pathway that regulates many normal cellular processes including cell survival, proliferation, growth, and motility. Activated growth factor receptors directly stimulate class 1A PI3Ks bound via their regulatory subunit or adapter molecules, which trigger activation of PI3K and conversion by its catalytic domain of phosphatidylinositol (3,4)-bisphosphate (PIP2) lipids to phosphatidylinositol (3,4,5)-trisphosphate (PIP3). Subsequently, PIP3 recruits and activates downstream serine-threonine protein kinase AKT. AKT in turn regulates a wide range of target proteins including mTOR that control protein synthesis and cell growth (Hemmings and Restuccia, 2012).
In the meanwhile, PTEN, a PIP3 phosphatase, acts as a suppressor of this pathway by converting PIP3 back to PIP2 and thus shutting off PI3K signaling. Aberrant activation of PI3K/AKT/mTOR pathway has been widely implicated in many cancers. Activation of this pathway can be a result of loss of PTEN, mutations in the PI3K or AKT genes, or constitutive activation of upstream regulatory pathways such as receptor tyrosine kinases (Courtney et al., 2010).
Our work showed an inverse association of PTEN and miR-21 level in gastric cancer cells. When miR-21 was inhibited, both protein level and gene expression of PTEN was induced. PTEN is one of the most frequently mutated tumor suppressors in human cancer. Early works by Meng et al. found that PTEN was a direct target of miR-21 and it contributed to miR-21 effects on proliferation, migration, and invasion of cholangiocarcinoma and hepatocellular cancer cells. Here, we showed that in gastric cancer cells, miR-21 inhibitor inhibited cancer cell proliferation, migration, and invasion via upregulation of PTEN. pAKT is the readout for PI3K, phosphor S6 (S235/5 or S240/4) is the readout for mTORC1. Modulation of miR-21 reduced the ratios of S235/236, S240/244 and p-AK/AKT.
In addition, miR-21 inhibitor also regulated apoptosis of gastric cancer cells. Though widely observed in other types of cancer (Darido et al., 2011), the importance of miR-21/PTEN pathway in gastric cancer is considerably less clear. Motoyama et al. (2010) investigated the association between the expression levels of miR-21 and PDCD4, another tumor suppressor, in eight human gastric carcinoma cell lines besides MKN74. They found an inverse correlation between the expression of PDCD4 mRNA and miR-21 in these cell lines and PDCD4 was identified as the target gene of miR-21. Recently, a study showed that Frizzled class receptor 6, a family member of noncanonical Wnt signaling, was involved in miR-21-dependent effects in gastric cancer (Yan et al., 2016).
However, our findings are consistent with the work on another gastric cancer cell line BCG-823 in which overexpression of miR-21 promoted BCG-823 cell growth, invasion, and migration whereas downregulation of miR-21 exhibited an inhibitory effect on the biological behavior of gastric cancer cells by targeting PTEN. miR-21/PTEN pathway was also found essential for the sensitivity of gastric cancer cells to chemotherapeutic drugs trastuzumab (Eto et al., 2014) and cisplatin (Yang et al., 2013). Epithelialmesenchymal transition is another important biological function of cancer cells. miR-21 promoted TGFb1-induced epithelial-mesenchymal transition in gastric cancer through upregulating PTEN expression (Li et al., 2016). These results suggest that although multiple signaling pathways may exist in a cell-specific way, PTEN/PI3K/mTOR is one of the most important pathways mediating the effects of miR-21 in gastric cancer and targeting miR-21 and its downstream PTEN/PI3K/mTOR signaling pathway might be a promising strategy for the prevention and treatment of gastric cancer.
Footnotes
Disclosure Statement
No competing financial interests exist.