
Abstract
The mitotic spindle, assembled by microtubule organization center (MTOC), is a critical structure required for proper chromosome segregation during mitosis. Studies demonstrated that defects of mitotic spindle assembling would result in chromosome instability, associating with oncogenesis. In this study, we identified that heparanase (HPSE) was localized on MTOC in mitotic cells. Induced expression of HPSE in human airway epithelial cells promoted mitotic spindle formation, and silencing of HPSE expression disrupted the mitotic spindle formation, resulting in chromosome instability, including chromosome mis-segregation and increased micronuclei formation. In conclusion, HPSE is an important protein in regulation of mitotic spindle and loss of HPSE function on MTOC associates with chromosome instability, indicating that loss of HPSE on MTOC may play critical roles in oncogenesis.
Introduction
T
Heparanase (HPSE) is the only known mammalian endoglycosidase. It regulates the function of HSPGs through remodeling their HS side chain (Fernandes et al., 2014). Previous studies showed that increased expression of HPSE was found to be associated with tumor angiogenesis and metastasis through modifying the HS side chain of its substrate proteins in extracellular matrix (ECM) (Nakajima et al., 1988; Vlodavsky et al., 1994; Bergers et al., 2000; Elkin et al., 2001). Studies also identified that HPSE located in cell nuclei or cytoplasm in lung cancer tissues and the cytoplasmic location significantly correlated with poorer prognosis of the patients (Schubert et al., 2004; Cohen et al., 2008), indicating that HPSE has different functions at different cellular compartments. Whether HPSE locates on mitotic spindle and functions in maintaining chromosome stability of the cell are unknown. In this research, we found that HPSE located on MTOC and silencing of HPSE expression disrupted mitotic spindle formation, resulting in chromosomal mis-segregation and micronuclei formation. These data demonstrated that HPSE was an important regulator of the mitotic spindle, and dysregulation of HPSE may associate with induction of chromosome instability during oncogenesis.
Materials and Methods
Cell culture
Human bronchial epithelial cells (HBECs), a human airway epithelial cell line, were cultured in Keratinocyte-SFM Basal medium (GIBCO) supplemented with Bovine Pituitary Extract (GIBCO) and Recombinant Epidermal Growth Factor (GIBCO).
Calculation of mitotic cells with normal mitotic spindle (%)
pcDNA3.1 or pcDNA3.1-HPSE was transfected into cultured cells using Lipofectamine 2000 (Invitrogen, Grand Island, NY). Forty-eight hours after transfection, microtubule inhibitor nocodazole (Andreassen and Margolis, 1994) was added (final concentration: 100 nM) to the media for 18 h to disrupt the mitotic spindle, then cells were washed with PBS thrice to release nocodazole, and fixed at different time points. Immunofluorescence staining of α-tubulin and DAPI was carried out, and pictures were taken using Olympus BX51 microscope. Total numbers of mitotic cells at prophase, metaphase, and anaphase were counted (n = 200), and among those cells, the number of mitotic cells with normal mitotic spindle (Fig. 3A′) were counted. Percentage of mitotic cells with normal mitotic spindle was calculated as per following equation: (number of mitotic cells with normal mitotic spindle/total numbers of mitotic cells) × 100%. Independently repeated experiments (n = 3) were performed, and data are expressed as the mean ± standard error of the mean (SEM).
Calculation of mitotic cells with abnormal mitotic spindle (%)
HBECs were transfected with Hpse siRNA (oligo ID HSS116734 or oligo ID HSS116735; Invitrogen) or control siRNA (Cat. No. 12935-300; Invitrogen) and harvested at 48 h after transfection. Immunofluorescence staining of α-tubulin and DAPI was carried out. And pictures were taken using Olympus BX51 microscope. Total mitotic cells at prophase, metaphase, and anaphase (n = 200) were counted, and among those cells, mitotic cells with abnormal spindles (Fig. 4A–F) were counted, and the percentage of mitotic cells with abnormal spindle was calculated as per following equation: (the number of mitotic cells with abnormal spindle/total mitotic cells at prophase, metaphase, and anaphase) × 100%. Independently repeated experiments (n = 3) were performed, and data are expressed as the mean ± SEM.
Immunofluorescence staining
Cells were cultured on coverslips, harvested and immunostained as previously described (Wang et al., 2014). Primary antibodies are listed in Table 1. Fluorochrome-conjugated secondary antibodies (Alexa Fluor 488 or 594; Invitrogen) were used. Images were captured with an Olympus BX51 microscope.
HPSE, heparanase; IF, immunofluorescence; WB, Western Blot.
Western Blot
Cells lysates were harvested at 72 h after transfection, and Western Blot was performed. Antibodies are listed in Table 1. Independently repeated experiments were performed (n = 3), densitometry quantification of the Western Blot bands were carried out using Quality One Software (Bio-Rad Laboratories, Philadelphia, PA), and data are expressed as the mean ± SEM. All images are representative of corresponding experiments.
Protein immunoprecipitation assay
Immunoprecipitation assays were performed using protein G agarose beads according to the manufacturer's protocol (Roche). Briefly, total cell lysates were collected at 72 h after transfection and precleared with protein G agarose, then incubated with HPSE specific antibody for 3 h at 4°C, followed by incubation with protein G agarose for 3 h at 4°C. Binding proteins were eluted from pelleted beads in the gel-loading buffer. The γ-tubulin protein level was analyzed by Western blot. Independently repeated experiments were performed (n = 3); all images are representative of corresponding experiments.
Flow cytometry analysis
Cells were treated as previously described (Crasta et al., 2012). Briefly, cells were cultured and transfected with control siRNA or Hpse siRNA. Seventy-two hours after transfection, cells were harvested and fixed in 75% ethanol at 4°C overnight, followed by incubation in RNase A solution (50 μg/mL) at 37°C for 15 min, and stained with propidium iodide (Sigma) for 30 min in the dark at 37°C. The percentage of cells in each cell cycle phase was measured according to the S-fit program in the FACSta. Independently repeated experiments (n = 3) were performed, and data are expressed as the mean ± SEM.
Statistical analysis
All data were analyzed using SAS software (version9.0; SAS Institute, Cary, NC). Student's t-test was used to determine differences between groups. The value p < 0.05 was considered as statistically significant.
Results
The highest level of HPSE protein was identified at metaphase in epithelial cells
To explore the function of HPSE protein during mitosis, HBECs were synchronized as described previously (Whitfield et al., 2002) and harvested at different time points after releasing of cell cycle synchronization. Western blot analysis demonstrated that protein level of HPSE was dynamically changed during mitosis, associating with changes of phospho-histone H3, a mitosis marker (Demion et al., 2014) (Fig. 1A). Consistent with these findings, fluorescent immunocytochemistry staining of HPSE in HBEC demonstrated that cytoplasmic staining of HPSE was significantly increased in cells at prophase, metaphase, and anaphase (Fig. 1B′–B′″), compared with cells at interphase (Fig. 1B), and the highest level of HPSE protein was observed at metaphase. Similar findings were also identified in other cell lines, for example, HaCaT cells (Supplementary Fig. S1A–A′″; Supplementary Data are available online at
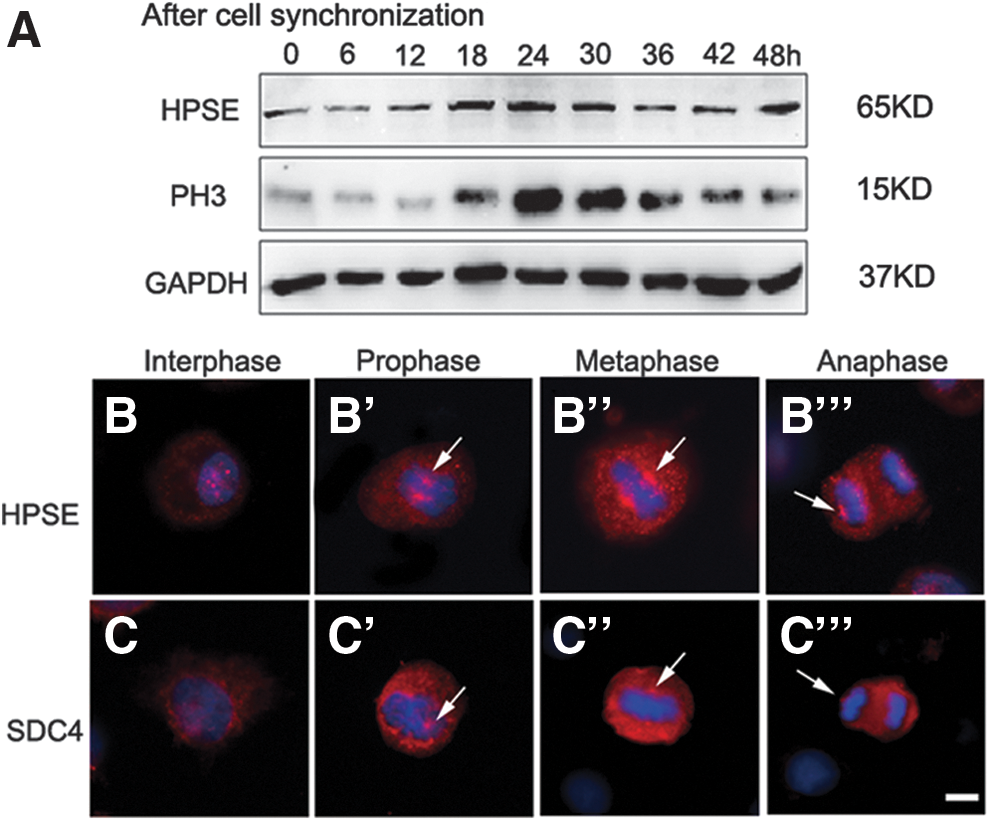
The dynamic change of HPSE in HBECs during mitosis.
HPSE located on MTOC
To determine whether HPSE locates on MTOC, co-fluorescent immunohistochemistry staining of HPSE with γ-tubulin, a MTOC associating protein, was carried out in HBEC. Although HPSE staining was observed throughout the cytoplasm, enriched staining of HPSE was identified in regions with γ-tubulin staining (Fig. 2A–C), indicating that HPSE was located on MTOC. To test whether HPSE and γ-tubulin interact with each other, HBECs were transfected with pcDNA3.1-Hpse or pcDNA3.1 plasmid, respectively; cell extracts were prepared 72 h after transfection and were analyzed by immunoprecipitation with HPSE antibody. The precipitated material was separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis, and the presence of γ-tubulin was detected by western blotting with a γ-tubulin antibody (Fig. 2D, E). These data demonstrated that HPSE was located on MTOC and that it may play a critical role in regulating mitotic spindle.
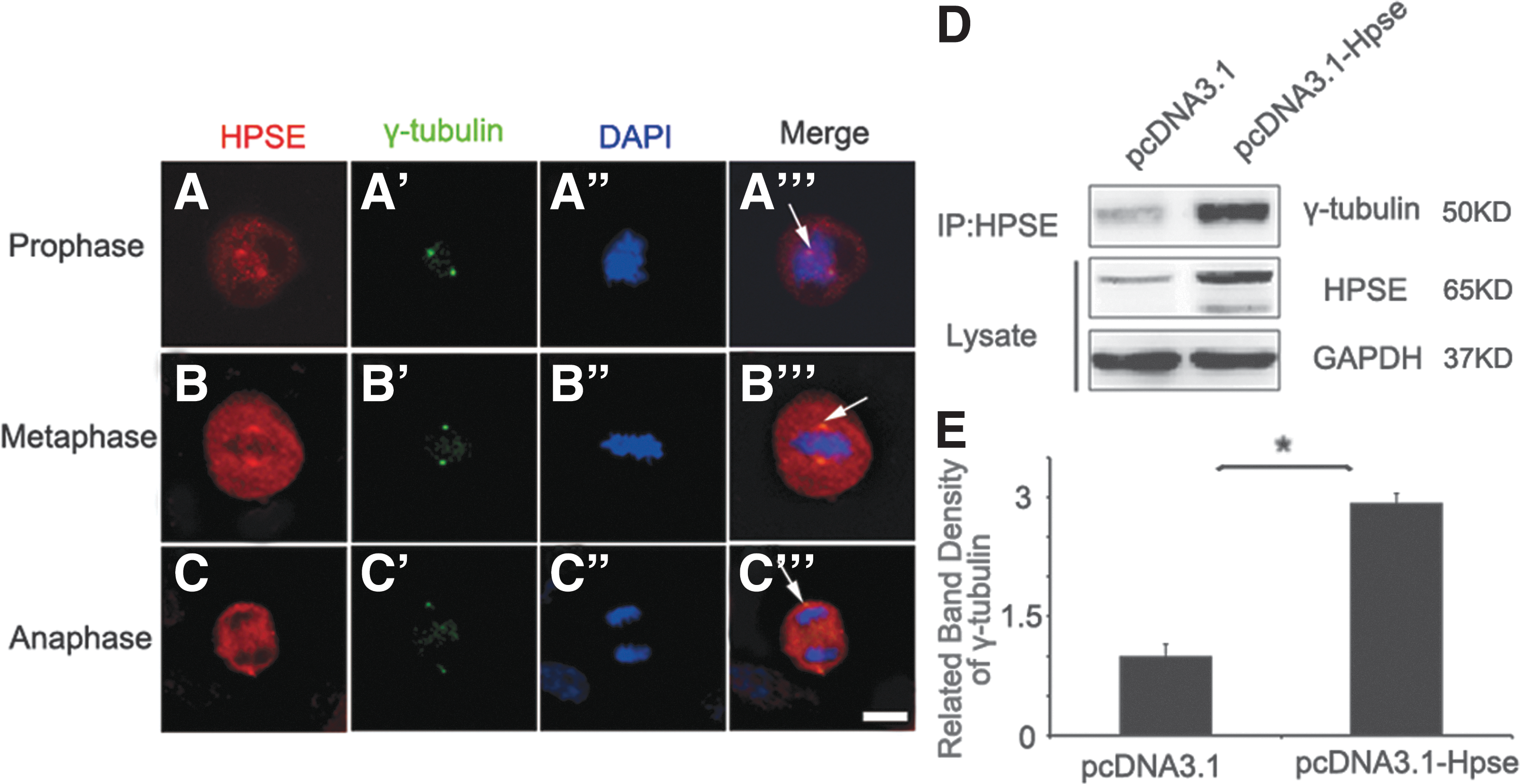
HPSE interacts with γ-tubulin.
Induced expression of HPSE promoted mitotic spindle formation
To access the role of HPSE in mitotic spindle formation, pcDNA3.1-Hpse or pcDNA3.1 was transfected into HBECs, respectively, 48 h after transfection. Nocodazole, a microtubule inhibitor, was added to the media for 18 h to stop the spindle formation. Then, cells were harvested at 0- and 10-min time points after nocodazole releasing. Immunofluorescent staining of α-tubulin, a spindle marker, was carried out to show the mitotic spindle structure. At 0-min time point, metaphase spindle formation was stopped in all the mitotic cells (Fig. 3A); no difference was identified between pcDNA3.1-Hpse transfected cells and pcDNA3.1 transfected cells. At 10-min time point, some cells with normal mitotic spindle structure were visualized (Fig. 3A′); percentage of mitotic cells with normal mitotic spindle was significantly increased in cells transfected with pcDNA3.1-Hpse, compared with the controls (Fig. 3B), indicating that HPSE promoted mitotic spindle formation. Consistent with this, increased levels of proteins functioning on mitotic spindle, including α-tubulin, acetylated α-tubulin, and γ-tubulin, were identified by Western blot in HBEC transfected with pcDNA3.1-Hpse, compared with controls (Fig. 3C, D). These data demonstrated that induced expression of HPSE could promote mitotic spindle formation.
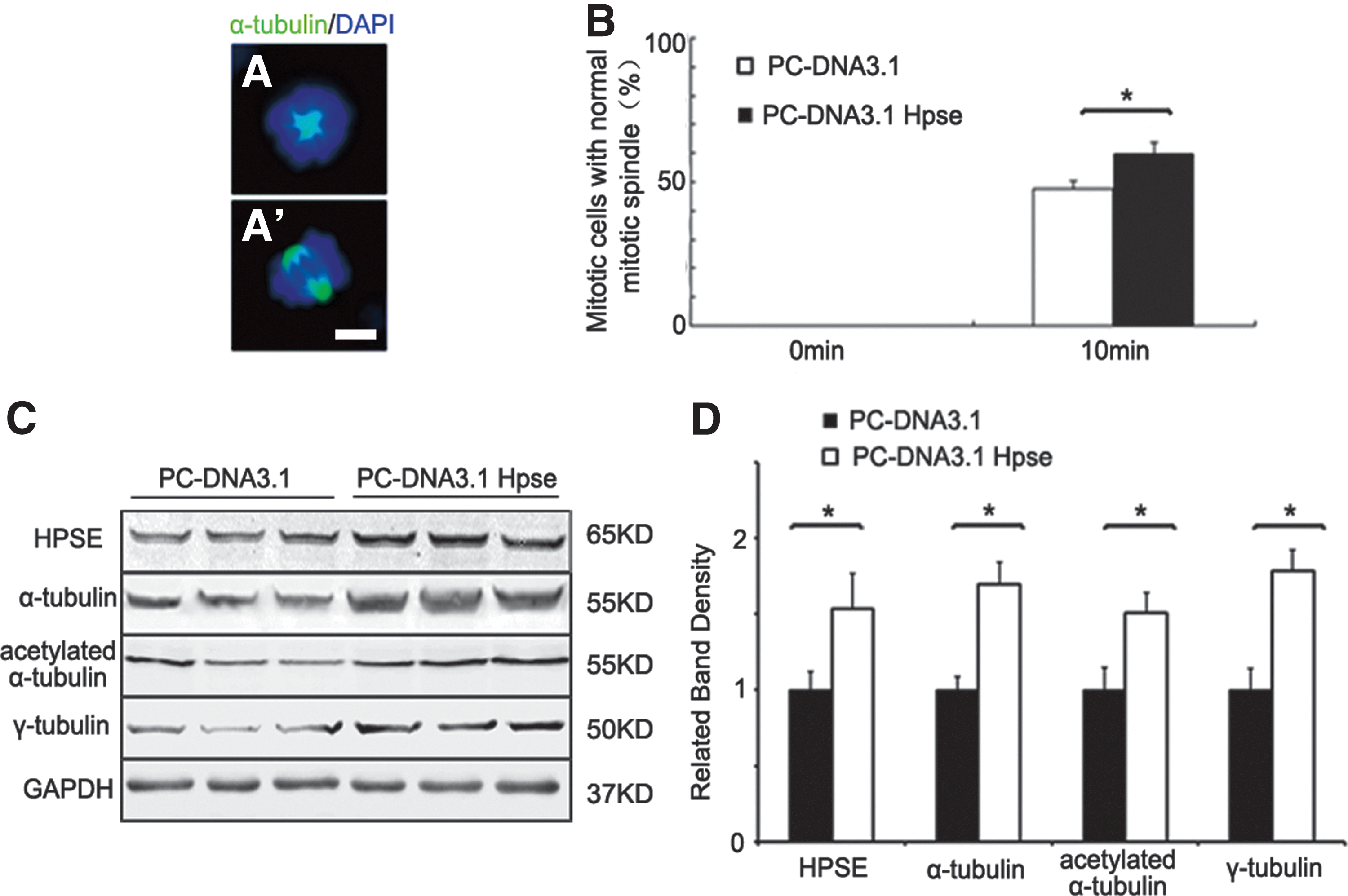
Induced expression of HPSE promoted mitotic spindle formation.
Silencing of HPSE expression resulted in mitotic spindle defects
To further identify the function of HPSE in mitotic spindle, Hpse siRNA or control siRNA was, respectively, transfected into HBECs. Cells were harvested 72 h after transfection, and immunofluorescent staining of α-tubulin was carried out. Mitotic spindle structure was outlined by α-tubulin staining. Compared with controls, percentage of mitotic cells with abnormal mitotic spindles, including cells with decreasing α-tubulin staining and cells with collapsed spindle, was significantly increased in Hpse silenced cells (Fig. 4A–G). Consistent with these findings, Western blot demonstrated that proteins functioning in spindle assembling, including α-tubulin, acetylated α-tubulin, and γ-tubulin, were significantly decreased in Hpse silenced cells, compared with controls (Fig. 4H, I). These results demonstrated that HPSE was critical for mitotic spindle.
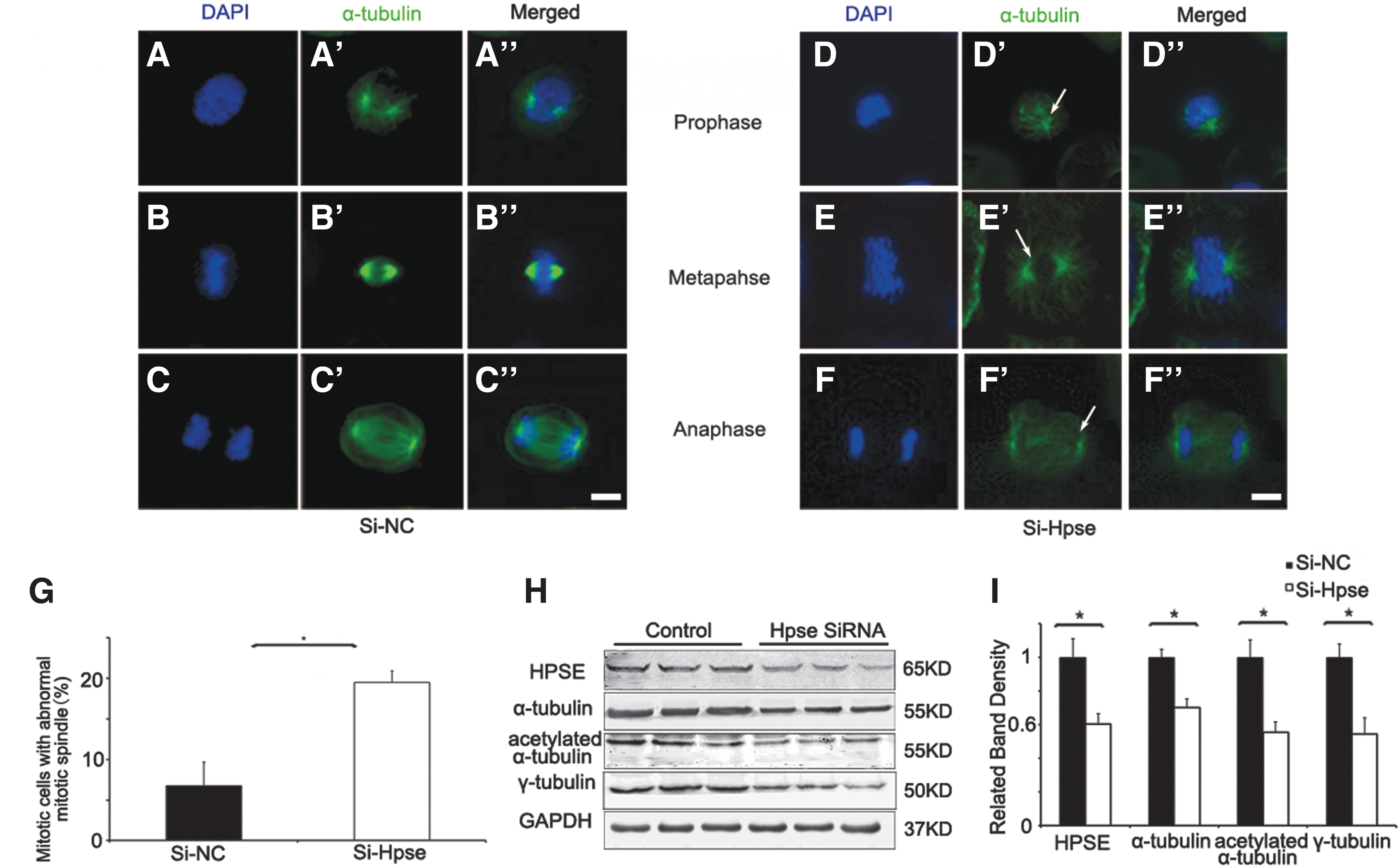
Silencing of HPSE expression induced abnormal mitotic spindle.
Silencing of HPSE expression induced chromosome mis-segregation
Mitotic spindle abnormalities would lead to chromosome mis-segregation, resulting in micronuclei formation (Crasta et al., 2012). To know whether HPSE silencing causes chromosome mis-segregation, immunofluorescence staining of α-tubulin and DAPI was performed in Hpse siRNA or control siRNA transfected HBECs. Lagging chromosomes were observed in Hpse silenced cells at anaphase (Fig. 5B, arrowheads), indicating that Hpse silencing affected the microtubule-chromosome attachment. Significantly increased incidence of micronuclei was observed in Hpse siRNA transfected HBECs compared with control (N = 200, *p < 0.05) (Fig. 5G). Chromosome mis-segregation would activate spindle checkpoint and induce G2/M cell cycle arrest. In agreement with this, G2/M arrest was observed in Hpse silenced HBECs by flow cytometry analysis (Supplementary Fig. S2). Taken together, these observations indicated that HPSE played a significant role in maintaining chromosome stability.
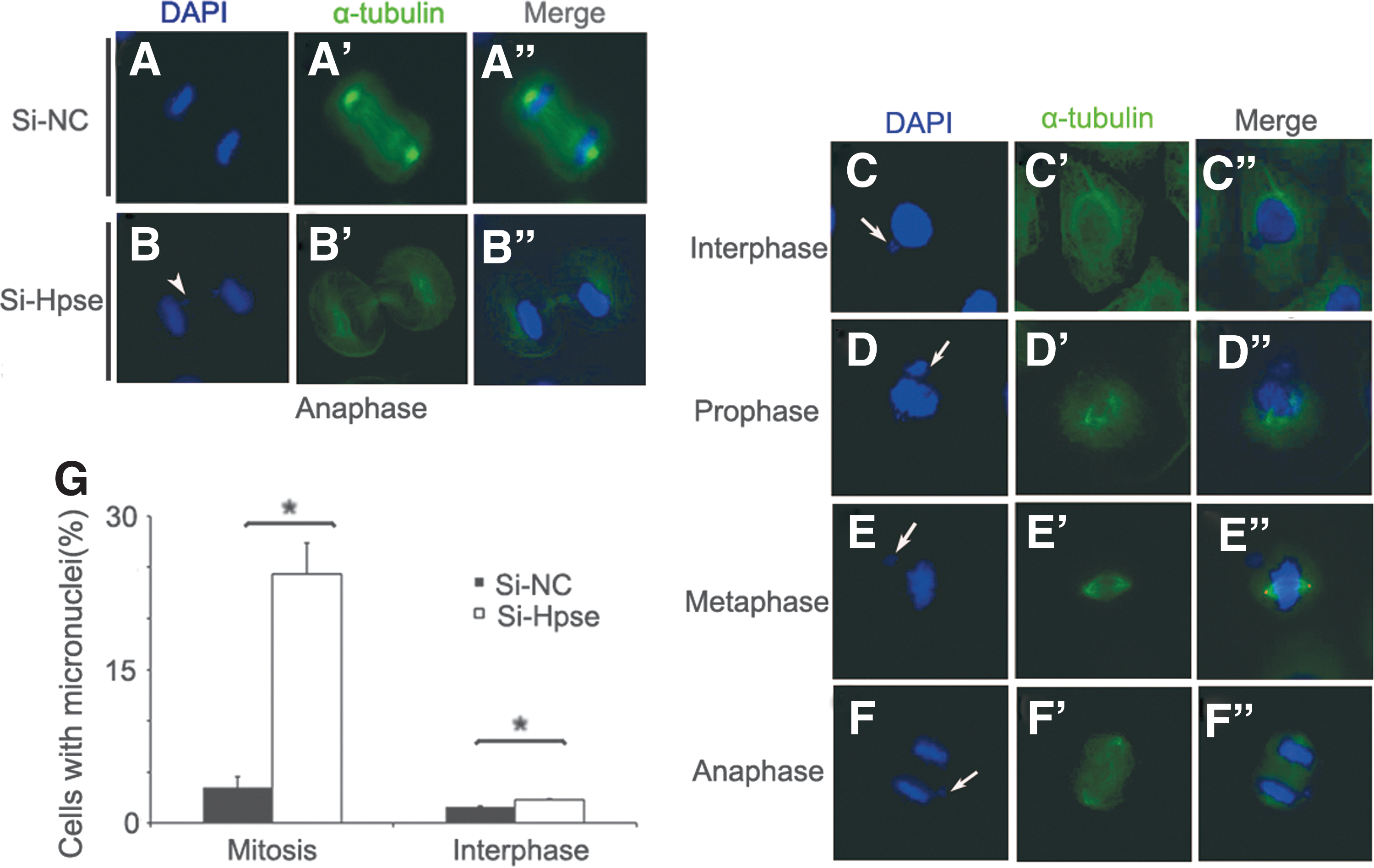
HPSE silencing induced chromosome mis-separation.
Discussion
Defect of mitotic spindle is one of the critical causes of chromosome instability associating with oncogenesis (Venkatesan et al., 2015). In this study, we identified that HPSE locates on MTOC and is indispensable for proper segregation of chromosomes during mitosis; loss of MTOC localization of HPSE associates with chromosome instability.
HPSE is an important protein involved in regulation of mitotic spindle
Previous studies demonstrated that HPSE was a glycosidase and functions in cell proliferation, differentiation, and migration through modifying HSPGs located in different cellular compartments (Gingis-Velitski et al., 2004; Kobayashi et al., 2006; Nobuhisa et al., 2007; Zheng et al., 2015). In late endosome and lysosome, HPSE involved in the processing and recycling of heparin sulfate from internalized HSPGs (Goldshmidt et al., 2002; Cohen et al., 2005). In ECM, HPSE played a key role in both normal and disease processes such as cell migration, wound healing, angiogenesis, and inflammation (McKenzie, 2007). In the nucleus, it was thought to play a role in nuclear HS degradation and stimulation of gene expression (Schubert et al., 2004). In this research, HPSE was found to be enriched at M-phase during mitosis and co-localized with γ-tubulin on MTOC. HPSE silencing caused mitotic spindle defects, including decreasing of spindle protein levels (α-tubulin, acetylated α-tubulin, and γ-tubulin) and disruption of spindle structure, which in turn resulted in chromosome mis-segregation and micronuclei formation. This study identified a new cellular localization of HPSE and demonstrated that HPSE was an indispensable protein required for maintaining normal mitotic spindle structure. Loss of HPSE function on MTOC may play critical roles in chromosome instability during oncogenesis. The underline mechanism needs to be further explored.
Loss of HPSE function on MTOC associated with chromosome instability
Recent studies demonstrated that micronuclei, formed from chromosome mis-segregation and spindle checkpoint defect, can persist overall for several generations and generate more chromosome breaks and DNA mutations (Crasta et al., 2012). These provided strong mechanistic evidence that increased rates of chromosome mis-segregation and spindle checkpoint defect can accelerate oncogenesis. Our study demonstrated that loss of HPSE function on MTOC caused the defects in mitotic spindle, which consequently affected the chromosome segregation and resulted in increased micronuclei formation. These data indicated that loss of HPSE function on MTOC could be one cause of chromosome instability associating with oncogenesis. A significant G2/M arrest has been observed in Hpse silenced cell, indicating that chromosome mis-segregation caused by Hpse silencing activated spindle checkpoint machinery, and the defects could be partially corrected by intact function of spindle checkpoint. Compared with molecules functioning in negative regulation of both chromosome segregation and spindle checkpoint, including TAO1 and ZM447439 (Ditchfield et al., 2003; Viji et al., 2007; Roshan et al., 2014), silencing of Hpse resulted in much less changes of chromosome instability.
A large number of researches clearly link increased expression of HPSE to the process of tumorigenesis in a lot of cancers, including breast, colon, gastric, pancreatic, thyroid and prostate, and so on (Nadir and Brenner, 2014). In the present study, we identified that HPSE was localized on MTOC and that silencing of HPSE disrupted the mitotic spindle formation, resulting in chromosome instability, including chromosome mis-segregation and increased micronuclei formation. Whether the loss of MTOC localization of HPSE existed in cancer tissues and whether loss of MTOC localization of HPSE correlated with poor prognosis of cancer need to be further explored.
Footnotes
Acknowledgments
This research was supported by internal funds from the West China Second University Hospital and grants from the National Natural Science Foundation of China (NSFC81572754 and NSFC81322001, H.W.) and Fundamental Research Funds for the Central Universities (Excellent young scholars program, 2014SCU04B04, H.W.).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.