
Abstract
Lack of mechanical load leads to skeletal muscle atrophy, and one major underlying mechanism involves the myostatin pathway that negatively regulates protein synthesis and also activates Atrogin-1/MAFbx and MuRF1 genes. In hindlimb immobilization, leucine was observed to attenuate the upregulation of the referred atrogenes, thereby shortening the impact on fiber cross-sectional area, nonetheless, the possible connection with myostatin is still elusive. This study sought to verify the impact of leucine supplementation on myostatin expression. Male Wistar rats were supplemented with leucine and hindlimb immobilized for 3 and 7 days, after which soleus muscles were removed for morphometric measurements and analyzed for gene and protein expression by real-time PCR and Western blotting, respectively. Muscle wasting was prominent 7 days after immobilization, as expected, leucine feeding mitigated this effect. Atrogin-1/MAFbx gene expression was upregulated only after 3 days of immobilization, and this effect was attenuated by leucine supplementation. Atrogin-1/MAFbx protein levels were elevated after 7 days of immobilization, which leucine supplementation was not able to lessen. On the other hand, myostatin gene expression was upregulated in immobilization for 3 and 7 days, which returned to normal levels after leucine supplementation. Myostatin protein levels followed gene expression at a 3-day time point only. Follistatin gene expression was upregulated during immobilization and accentuated by leucine after 3 days of supplementation. Concerning protein expression, follistatin was not altered neither by immobilization nor in immobilized animals treated with leucine. In conclusion, leucine protects against skeletal muscle mass loss during disuse, and the underlying molecular mechanisms appear to involve myostatin inhibition and Atrogin-1 normalization independently of follistatin signaling.
Introduction
In humans, ∼40
Regarding load status, increased stimulation will often induce muscle hypertrophy. In contrast, the lack of mechanical load often induces skeletal muscle atrophy. This phenomenon, known as disuse, is evident in cases in which healthy humans undergo a period of inactivity by limb immobilization or bed rest (Brooks and Myburgh, 2014; Wall et al., 2014).
The absence of weight and tension stimuli on skeletal muscle leads to significant physiological and molecular changes, affecting almost every aspect of muscle function. For example, after 2 to 5 weeks of inactivity, metabolic rate and insulin sensitivity adapt to lower loads (Brooks and Myburgh, 2014; Wall et al., 2014). At the single myocyte level, complex and not fully understood mechanisms trigger enhanced proteolysis on top of reduced de novo protein synthesis, leading to an overall loss of sarcomeres. Although the specific gene networks that drive anabolic and catabolic signaling in each type of muscle wasting requires further understanding, the outcome of this altered protein turnover is atrophy (Jackman and Kandarian, 2004). Atrophy, also known as muscle wasting, is characterized by depletion of organelles and proteins with major impact upon myofibrillar content through the ubiquitin/proteasome system (UPS) degradation (Sandri, 2008).
In this regard, myostatin is an interesting signaling factor, since it negatively regulates protein synthesis and myogenesis, while also inducing protein breakdown (Trendelenburg et al., 2009; Elkina et al., 2011; Lokireddy et al., 2011; Lokireddy et al., 2012; Wang et al., 2015). With regard to muscle protein breakdown, the UPS, and particularly the so-called E3 ligases are essential to target proteins, which are then directed and degraded at the 26S proteasome. Among these enzymes, both Atrogin-1/MAFbx and MuRF1 are induced by several atrophic models and are thus designated atrogenes. In disuse, these atrogenes are especially upregulated and are considered a requirement for muscle atrophy and substrate specificity (Bodine et al., 2001; Bodine and Baehr, 2014).
Several studies aimed to downregulate atrogenes and/or to decrease protein breakdown by using amino acid supplementation, including leucine. This amino acid has been shown to attenuate mass loss driven by immobilization in rat soleus muscle (Anthony et al., 2000; Ventrucci et al., 2001; van Norren et al., 2009; Peters et al., 2011; Baptista et al., 2013; Pereira et al., 2014).
Baptista et al. (2010) noted that leucine supplementation severely minimizes the Atrogin-1/MAFbx and MuRF-1 upregulation peak induced by cast immobilization for 3 days, shortening the effects of disuse on the cross-sectional area (CSA) of type I fibers. The capacity of leucine to induce protein synthesis while attenuating atrogene expression raises the possibility of its interaction with the myostatin pathway. Therefore, we investigated, in this study, in a rat skeletal muscle atrophy model, whether leucine effects are correlated to myostatin expression.
Materials and Methods
Experimental groups and animals
Experimental procedures were allowed by COBEA (Colégio Brasileiro de Experimentação Animal) under #88 approval from the Ethics Committee CEUA (Comissão de Ética no Uso de Animais). The animals, 2-month-old Wistar rats, were supplied by Central Bioterium at University of São Paulo.
Animals accessed food and water ad libitum, and the circadian cycle was maintained by a 12-h light/12-h dark protocol. Then, animals were arbitrarily sorted into groups as follows: (1) control group (CON; nonchallenged rats that received saline solution through gavage); (2) leucine group (LEU; nonchallenged rats that received leucine solution through gavage); (3) immobilized group (IMOB; hindlimb immobilized rats that received saline solution through gavage; and (4) immobilized and supplemented with leucine (IMOB+LEU; hindlimb immobilized rats that received leucine solution through gavage). The immobilization and supplementation procedures were conducted for 3 and 7 days.
Hindlimb Immobilization and supplementation process
For hindlimb immobilization, animals were anesthetized by a ketamine and xylazine mixture (30 mg/kg rat of ketamine and 10 mg/kg rat of xylazine). Then, micropore tape was attached at the left rat hindlimb such that the whole limb was maintained in the extended position, and the soleus muscle fixed in a shortened position. After drying, the cast was involved in a galvanized steel wire mesh (2 mm opening), which was subsequently fixed by a 0.5 mm diameter steel galvanized wire. The purpose of involving the cast with a mesh was to prevent it to be chewed by the animal.
A standard rat chow was provided for all groups (Nuvilab CR-1, Nuvital-Quimtia, Brazil) and a high dose of leucine was applied through gavage. The animals received two rounds of leucine supplementation (L-Leucine, Ajinomoto) consisting of a 2.7 g/kg/day body mass, prepared in 0.9% saline. All the treatments began 3 days before immobilization, and the control group received 0.9% saline on the same gavage volumes. After each set period, the animals were euthanized by cervical dislocation. The soleus muscle was then isolated, measured, and dry weighed. To obtain dry weight, muscles were kept at 60°C until stable mass was reached (Gissel, 2010). Finally, the muscle samples were frozen in liquid nitrogen and kept at −80°C until they were analyzed.
Gene expression analysis by real-time PCR
Real-time polymerase chain reaction (PCR) was utilized to measure mRNA levels. First, from soleus muscle, total RNA was isolated by TRIzol reagent (Life Technologies, Carlsbad, CA). Then, cDNA was synthesized by reverse transcriptase reaction.
This reaction involved 1 μg of total RNA, 500 μg/mL of oligo-dT, 0.1 M dithiothreitol (DTT), 200 U reverse transcriptase (SuperScript II; Invitrogen, San Diego, California), 10 mM of deoxyribonucleoside phosphate (dNTP), and 5 × first-strand buffer. The reaction was carried on a thermal cycler at 70°C for 10 min, 42°C for 60 min, and 95°C for 10 min.
Primer sets for rats mice were designed on the Primer Blast platform targeting the following genes: Myostatin, Follistatin, MAFbx/Atrogin-1, MuRF-1, and Cyclophilin A. Either 5′ or 3′ primer spanned exon-exon junctions (Table 1).
Primer Sets Used in Real-Time PCR
Real-time PCR templates were mixed at SYBR Green Universal Master Mix II (Life Technologies, Carlsbad, CA), 5` primer, and 3` primer. Then, the samples were incubated at 50°C for 2 min and 95°C for 10 min, followed by 40 cycles of 95°C for 15 s and 60°C for 1 min. The fluorescence intensity was quantified by a RotorGene 6000 (Qiagen, Germany).
A comparative threshold method in which gene expression was defined as the fold change over control values was utilized to obtain the results. Cyclophilin A was standardized as a housekeeping gene.
Western blotting
Soleus muscle was homogenized in modified RIPA buffer (1 mM EDTA pH 8.0, 0.00625 M sodium phosphate pH 7.2, 0.625% deoxycholic acid, 0.625% Nonidet P-40), including 10 μg/mL of protease inhibitor cocktail (Sigma-Aldrich). Then, the samples were centrifuged at 10,000 g for 10 min at 4°C. The total amount of protein was quantified by Bradford Assay (Bio-Rad, Hercules, CA) using bovine serum albumin (BSA) as a standard.
Samples of those homogenates were passed through SDS-PAGE gel (sodium dodecyl sulfate/polyacrylamide) by electrophoresis and then transferred to a nitrocellulose membrane (Bio-Rad). To confirm the transfer and the equal protein load, Ponceau S staining was applied. After that step, the membranes were blocked for 1 h (5% BSA in tris-buffered saline + 0.1% Tween-20 at room temperature) followed by primary antibody overnight incubation at 4°C. Next, the membranes were washed for 15 min and maintained for 1 h in secondary antibody solution at room temperature. After a 15-min wash, the alkaline phosphatase system detected label proteins. Primary antibodies were Atrogin-1/MAFbx (#AP2041, 1:1000; ECM Biosciences), Myostatin (#19142-1-AP, 1:1000; Proteintech), Follistatin (#PA5-19787, 1:500; Thermo Fisher), GAPDH (#PA1-987, 1:3000; Thermo Fisher) and alpha-tubulin (#2125S, 1:1000; Cell Signaling). Secondary antibodies were rabbit anti-mouse immunoglobulin/AP (polyclonal, 1:10,000; Dako) and goat anti-rabbit immunoglobulin/AP (polyclonal, 1:10,000; Dako). Images were obtained and quantified by Vision-Capt software (Vilber Lourmart, France).
Histological analysis
Cross-sections measuring 10 μm were made in a cryostat (Leica CM3050, Nussloch, Germany). These transverse sections were stained with Hematoxylin and Eosin, and pictures were subsequently acquired under a light microscope. CSA of the fibers were measured by the freehand tool on ImageJ software (version 2.0; NIH), ∼500 fibers per group were measured and procedures were conducted blinded. The total number of fibers was estimated as shown in Ceglia et al. (2013).
Statistical analysis
The results were reported as the mean ± standard deviation (SD). As recommended, two-way analysis of variance followed by Tukey's posttest was applied. p < 0.05 was considered to be statistically significant.
Results
Leucine mitigates disuse-induced muscle wasting on the soleus muscle of rats
First, we confirmed the recognized effect of leucine in the protection of muscle mass in a disuse atrophy model (Figs. 1 and 2). After 3 days of cast immobilization, rat soleus muscle from nonsupplemented animals showed reduced mass (∼20%; Fig. 1B), although no changes were detected in fiber number and CSA (Fig. 1C, D, respectively).
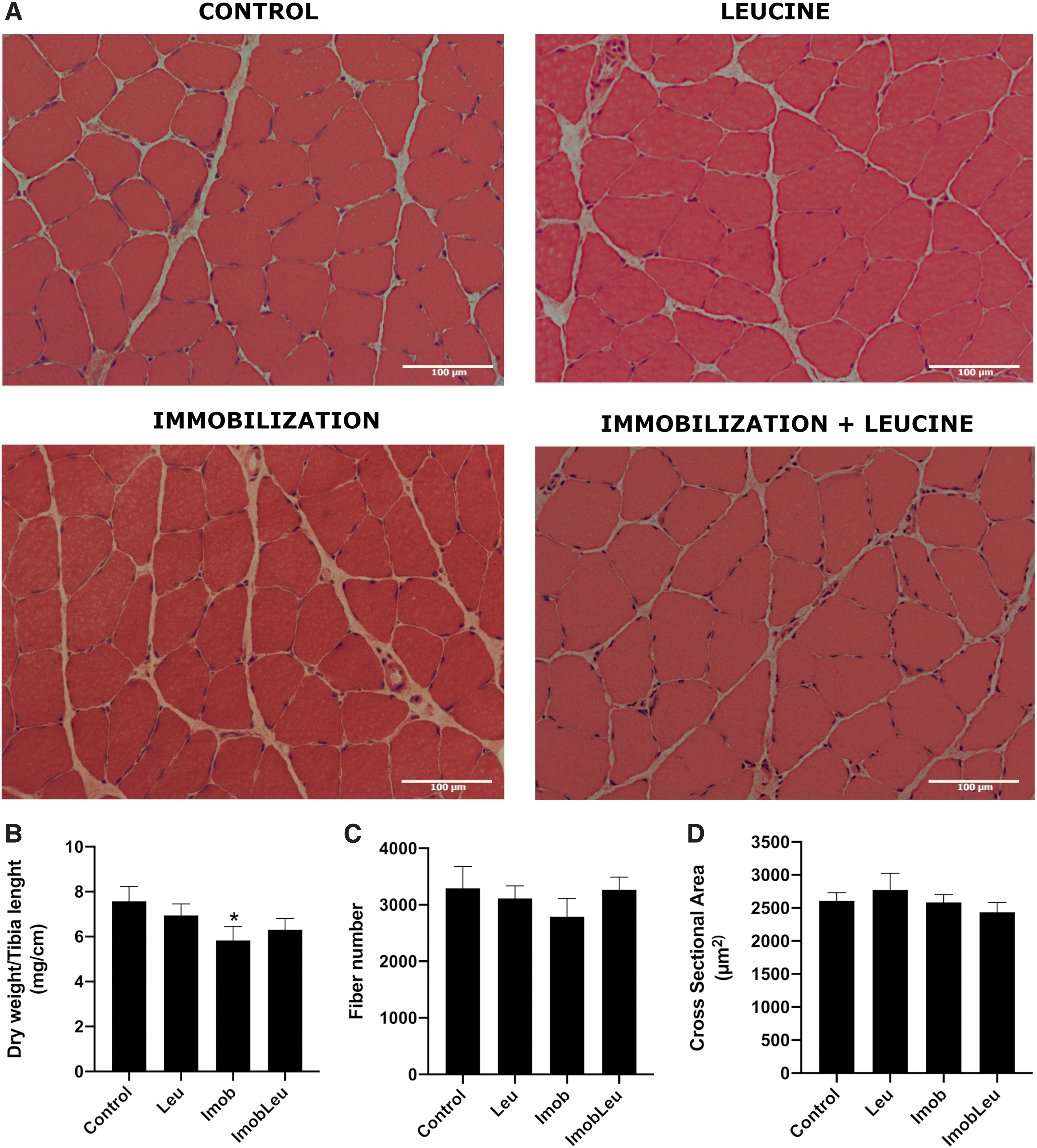

One week of hindlimb immobilization induced a 40% mass loss, whereas immobilized and leucine-supplemented animals exhibited a 30% decrease (Fig. 2B). Interestingly, leucine supplementation alone induced a 15% increase in mass (Fig. 2B). Regarding CSA of fibers, immobilized animals exhibited a 50% decrease in area, in contrast, immobilized animals that received daily doses of leucine for 7 days showed only about 17% reduction when compared with the control group (Fig. 2A, D). No changes were observed in total fiber counts. The effect of leucine on CSAs was restricted to the atrophy stress state. In summary, these results certify that the model utilized herein to induce atrophy and supplementation with leucine was accomplished efficiently.
Atrogin-1/MAFbx gene expression was upregulated upon immobilization and attenuated by leucine supplementation
To further certify the immobilization model and leucine treatment efficiency, we evaluate Atrogin-1/MAFbx gene expression and protein content over 3 and 7 days of immobilization and/or leucine supplementation (Fig. 3).
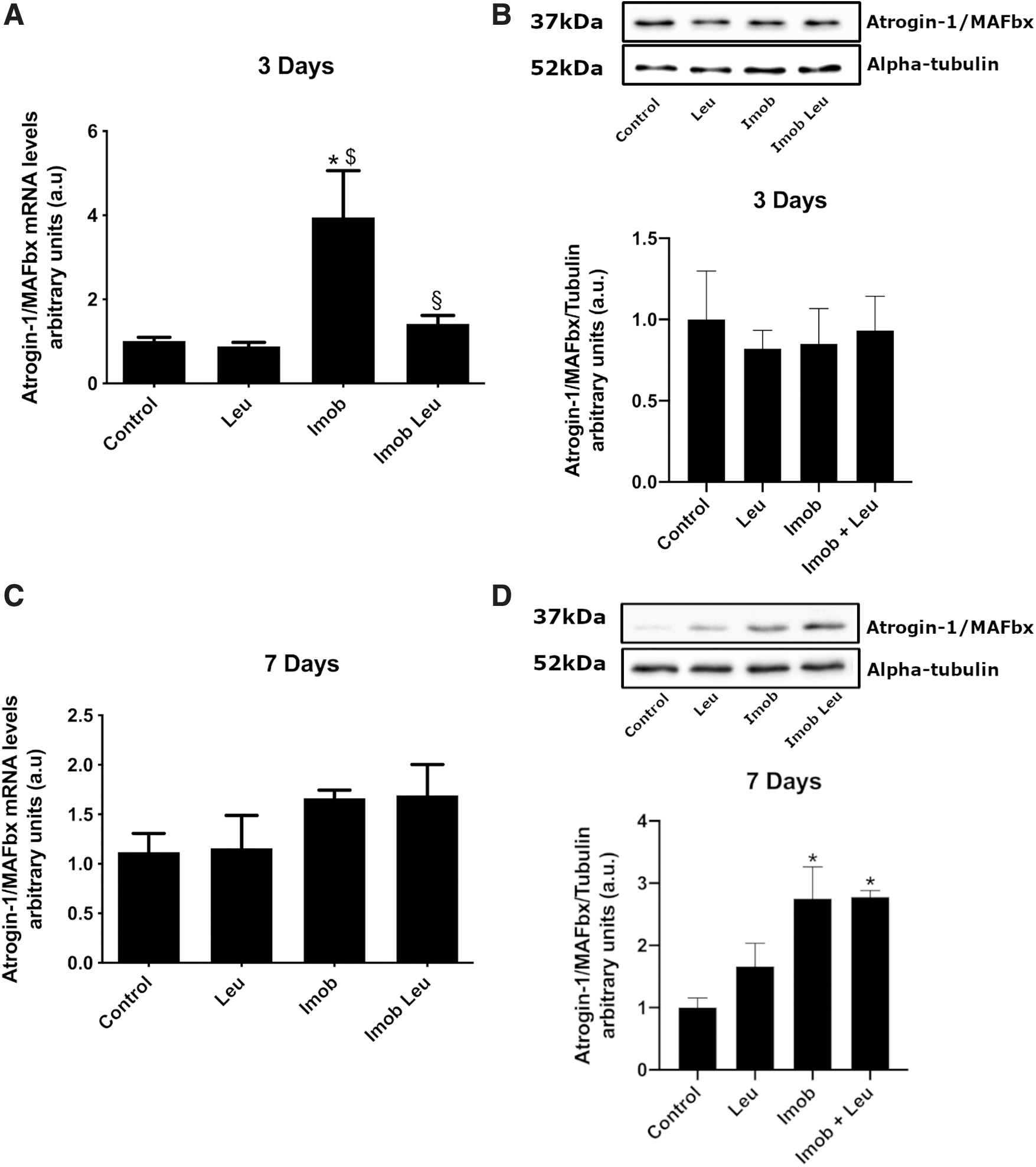
By the third day (Fig. 3A) of immobilization, Atrogin-1/MAFbx gene expression was upregulated (∼4-fold) and mitigated by leucine supplementation (∼1.5-fold). On the seventh day (Fig. 3C), however, Atrogin-1/MAFbx gene expression was similar to control values in immobilization, as well as leucine-treated groups. Leucine supplementation alone did not affect the expression of this gene at any time. Concerning protein expression, no changes among groups were observed at the 3-day time point (Fig. 3B). On the other hand, at 7-day time point Atrogin-1/MAFbx protein levels increased by immobilization, although leucine was not able to impair this rise (Fig. 3D; imob: ∼3-fold; imob leu: ∼3-fold).
Myostatin was upregulated on immobilization and attenuated by leucine supplementation
As expected, immobilization was able to induce myostatin gene expression by 3 and 7 days (∼3 and 1.7-fold, respectively). This elevation was reduced by leucine supplementation to the level of the control group at all time points (Fig. 4A, C).
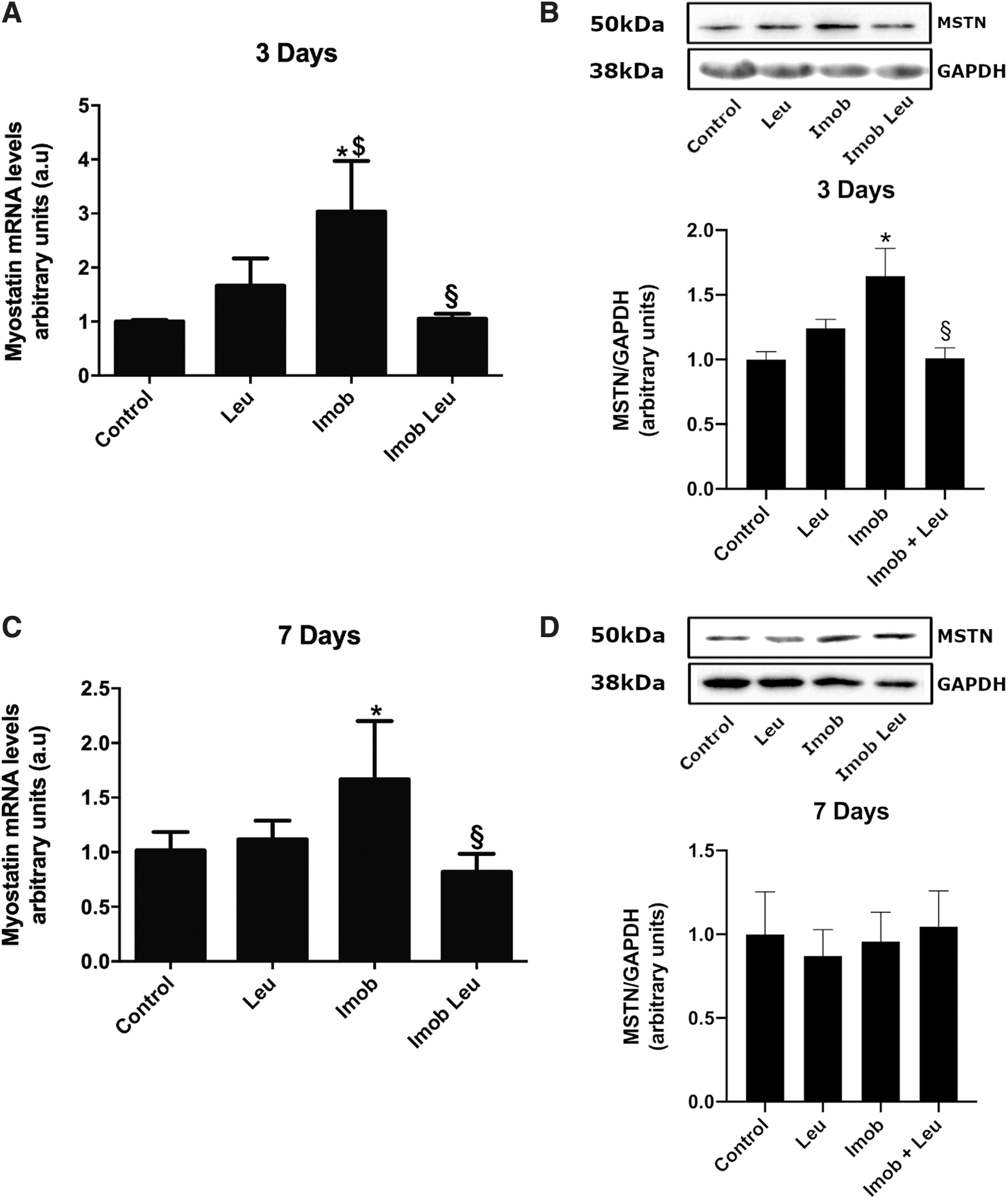
Protein expression was in line with mRNA data on the third day, being elevated by immobilization (∼1.6-fold). At this time point, leucine supplementation in immobilized animals was able to block the rise in myostatin protein expression, setting it to the level of the control group (Fig. 4B). At 7 days, myostatin protein expression remains unchanged in all groups analyzed (Fig. 4D).
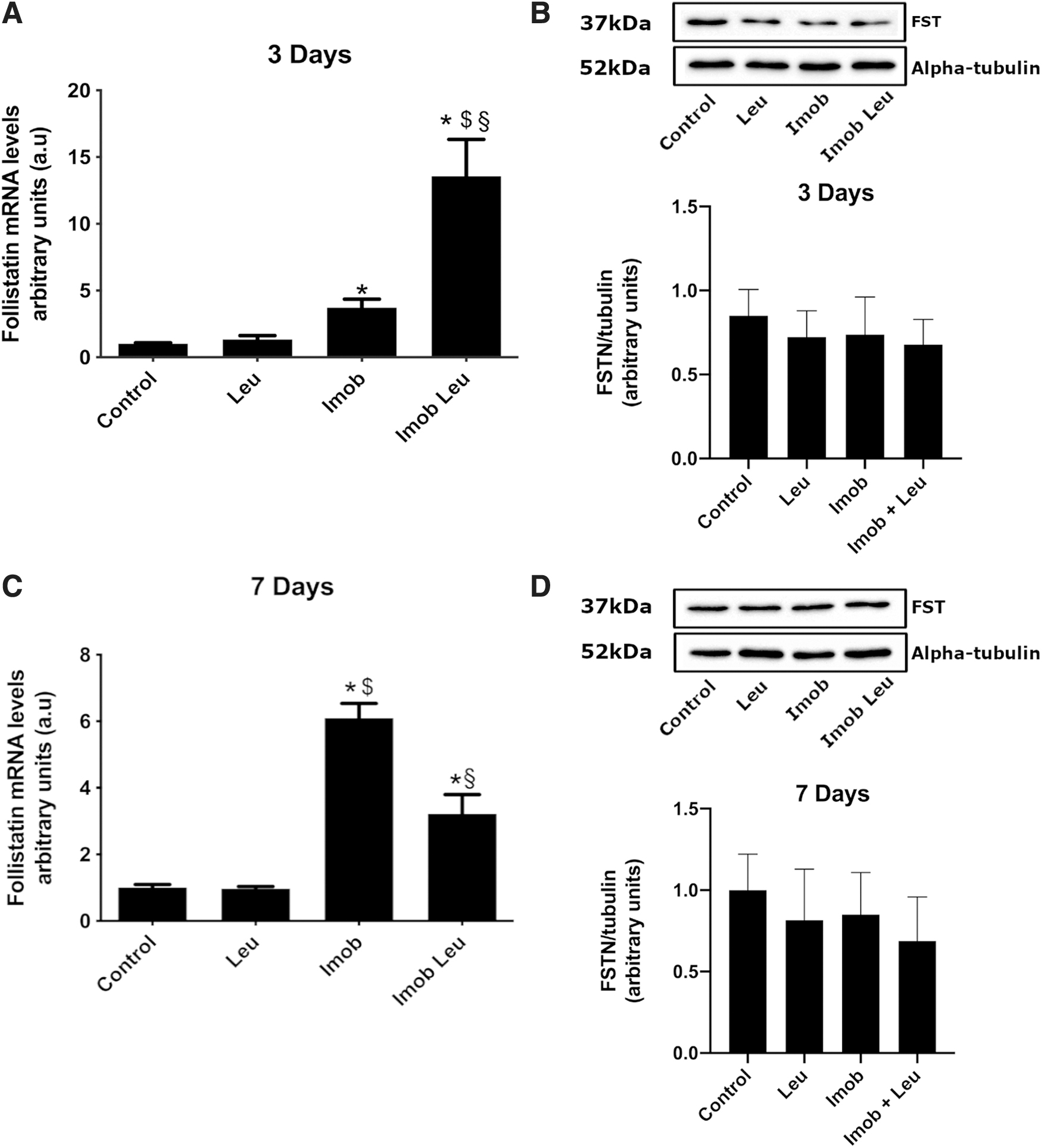
Follistatin gene expression was upregulated on immobilization and accentuated by leucine at third-day
Considering that myostatin expression was modulated by leucine, we decided to verify the expression of the inhibitor of myostatin, follistatin. We observed that immobilization induced increased follistatin expression on the third and on the seventh day (Fig. 5A, C). On the third day of immobilization, leucine supplementation induces upregulation of follistatin gene expression on a significant scale (∼3.7-fold) as compared with control. Remarkably, leucine supplementation in immobilized animals for 3 days further elevated follistatin gene expression very strongly (∼14-fold). On the seventh day, follistatin gene expression persisted elevated in immobilized animals (∼6-fold) and leucine supplementation in immobilized animals did not produce enhanced follistatin mRNA levels as in the 3-day group (Fig. 5C). In contrast to gene expression, follistatin protein content was unchanged at 3 and 7 days (Fig. 5B, D).
Discussion
Although starvation, glucocorticoid treatment, cachexia, kidney failure, aging, and mechanical unload are widespread examples of skeletal muscle mass loss, such models can modulate specific genes and even specific pathways (Sacheck et al., 2007). Among those models, mechanical unloading (i.e., cast immobilization) allows significant clinical insights to be reached regarding conditions where patients were submitted to bone fracture, peripheral nerve injury, or joint surgery. In humans, skeletal muscle wasting after mechanical unloading is detectable as early as 5 days following a decrease in strength and local metabolic changes, such as increased insulin resistance (Appell, 1990; Wall et al., 2014).
In rodents, cast immobilization as a model of mechanical unloading results in a rapid and consistent mass loss, also resulting in decreased force. Moreover, in this model, slow-twitch fibers are the most affected, which is in line with clinical conditions involving cast immobilization in humans (Booth and Kelso, 1973; Brooks and Myburgh, 2014). Indeed, in the present study, when we cast-hindlimb-immobilized rats for 3 and 7 days, the soleus muscle (∼90% slow-twitch fibers) showed a consistent loss of mass (∼23% and ∼40% decrease, respectively). After 1 week of cast immobilization, the CSA of the fibers was also reduced ∼48% when compared with control, indicating that the atrophy model used in this study was successfully employed (Figs. 1 and 2). Accordingly, the atrophy obtained in this study using the hindlimb immobilization model agrees with previous studies; for example, Baptista et al. (2013) observed ∼40% decrease in soleus mass and CSA in rat soleus muscle 1 week after immobilization. No changes were found regarding total number of fibers, which is in line with the study of Cardenas et al. (1977) and Nicks et al. (1989), which showed that skeletal muscle atrophy induced by immobilization do not occur through fiber number loss.
Several approaches have been employed aiming to mitigate skeletal muscle wasting, such as resistive physical exercise (Shen et al., 2018), blockage, or stimulation of certain intracellular pathways with antibodies or recombinant proteins, i.e., myostatin antibodies or follistatin overexpression (Sepulveda et al., 2015; Camporez et al., 2016; Desgeorges et al., 2017). Other noninvasive methods have been used, and supplementation with L-leucine appears to be an attractive strategy. It has been shown that leucine is capable of minimizing mass loss in cachexia, cancer, injury, and immobilization in rodents (Anthony et al., 2000; Ventrucci et al., 2001; van Norren et al., 2009; Peters et al., 2011; Baptista et al., 2013; Pereira et al., 2014). However, in humans, the anti-atrophic effects of leucine supplementation are still under debate. In certain studies, the protective effect of leucine is reported, while other authors found no protective effect. The differences in age of subjects, time of leucine supplementation (i.e., pre- vs. postprandial), and differences in dosage might explain those somewhat conflicting results (Ham et al., 2014; English et al., 2016; Ispoglou et al., 2016, 2017; Mitchell et al., 2017; Wolfe, 2017; Backx et al., 2018).
To track the atrophic process at the molecular level, we monitored Atrogin-1/MAFbx gene and protein expression 3 and 7 days after hindlimb immobilization (Fig. 3). It has been previously shown that this atrogene gene expression peaks at a 3-day time point (Bodine et al., 2001; Baptista et al., 2010; Okamoto et al., 2011), and as expected, we observed an increased Atrogin-1/MAFbx expression 3 days after immobilization. Leucine supplementation mitigated this increase, again demonstrating that the atrophy and leucine supplementation model employed in this study was efficient.
Nonetheless, atrogenes may not be the only components in muscle mass control. Indeed, other interrelated pathways, such as myostatin, do play a role, and in this study, we decided to evaluate whether leucine supplementation can modulate myostatin expression in skeletal muscle under disuse.
As expected, we found elevated levels of myostatin gene expression after 3 and 7 days of immobilization, and interestingly, leucine supplementation completely blocked this effect (Fig. 4A, C). Additionally, at the protein level, leucine blocked the rise in myostatin driven by 3 days of immobilization (Fig. 4B). On the other hand, no differences were found in myostatin protein levels among the groups after 7 days of immobilization (Fig. 4D). These data helped to elucidate the anti-atrophic effects of leucine, suggesting that myostatin mRNA regulation plays a role in the skeletal muscle atrophy and is sensitive to leucine.
The leucine effects on myostatin mRNA during immobilization could be related to transcription and posttranscriptional modulation. At transcriptional level, myostatin expression can be induced by the transcription factor FoxO (Allen and Unterman, 2007; Grade et al., 2019). Accordingly, FoxO activity is increased in disuse atrophy (Senf et al., 2010; Baptista et al., 2017) and leucine supplementation is able to decrease FoxO nuclear translocation during immobilization (Baptista et al., 2017). This effect could therefore be related to myostatin mRNA response both to immobilization and leucine.
Another possible mechanism includes posttranscriptional modulation by microRNAs. The miR-27 family of microRNAs has been shown to target and degrade myostatin and FoxOm RNAs, reducing their biological availability (Guttilla and White, 2009; Huang et al., 2012; Miretti et al., 2013). Interestingly, members of this family are upregulated by leucine supplementation (Chen et al., 2013); in this sense, the myostatin downregulation induced by leucine seen in this study could be related to miR-27 action upon myostatin itself and FoxO mRNAs. The dynamics of miR-27 expression during immobilization and supplementation is an interesting topic to address in forthcoming studies.
Although it is well known that regulation of myostatin by follistatin occurs by protein/protein interactions (Amthor, 2004; Sepulveda et al., 2015), we decided to verify follistatin expression because mRNA and protein levels can provide input on the possible amount of follistatin in the extracellular environment.
Follistatin overexpression promotes skeletal muscle hypertrophy and is known to ameliorate pathology in mdx, amyotrophic lateral sclerosis mice, and spinal muscular atrophy (Rose et al., 2009; Sepulveda et al., 2015; Iskenderian et al., 2018).
Interestingly, we were not able to find studies examining the regulation of follistatin expression in cast immobilization. In this study, surprisingly, we show that immobilization alone can strongly induce follistatin gene expression at 3 and 7 days after immobilization (Fig. 5A, C). Notably, leucine supplementation can boost the increase in follistatin driven by immobilization (from ∼3.7-fold to 14-fold). At 7 days of immobilization, follistatin mRNA levels were similar to those of the 3-day group, and the boost in follistatin mRNA driven by leucine was no longer observable.
In spite of changes in mRNA expression, we did not find a corresponding increase in follistatin protein levels; indeed, we found no changes in the levels of this protein in cast-immobilized animals (Fig. 5B, D). These results indicate that leucine can exert its anti-atrophic effect by decreasing myostatin protein expression independently of follistatin at the protein level. On the other hand, it is intriguing that such a strong response of follistatin occurs exclusively at the mRNA level. We envision that follistatin mRNA rise could be related to molecular mechanisms other than protein/protein interactions, such as those related to noncoding RNAs (Nie et al., 2015). For example, follistatin mRNA could generate a currently uncharacterized microRNAs that would, in turn, be able to inhibit mRNAs coding for stimulators of myostatin, such as NF-κβ-p65, MYOD, MYF5, and NFAT, all of which are known to be transactivators of the myostatin promoter. Alternatively, those microRNAs could even target myostatin mRNA itself (Huang et al., 2012; Qiu et al., 2013; Grade et al., 2019).
In fact, several studies have shown that mRNAs are able to produce functional microRNAs, and regulatory loops have been described (Ladewig et al., 2012; Ha and Kim, 2014; Titov and Vorozheykin, 2018). Nonetheless, to the best of our knowledge, at present, no study has directly demonstrated microRNAs originating from follistatin mRNA, although it has been shown that in the follistatin family, follistatin-like 1 (FSLT1) can produce miR-198 on its last exon (Hinske et al., 2010; Sundaram et al., 2013). Finally, additional recently described noncoding RNAs, such as circRNA and lncRNAs, could also be hosted in the follistatin gene and be related to proatrophic pathways.
This type of mechanism involving noncoding RNAs has been recently linked to skeletal muscle plasticity, including myogenesis and atrophy (Weng et al., 2018; Peng et al., 2019).
Conclusion
We demonstrated that leucine protects skeletal muscle mass loss by disuse through myostatin inhibition independently of follistatin signaling.
Footnotes
Acknowledgments
The authors thank FAPESP and CNPq for financial support. They are also grateful to the Foundation Leducq (network 13CVD04), and the European Union (Horizon 2020 grant agreement no. 645648 “Muscle Stress Relief”).
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the São Paulo Research Foundation (FAPESP) fellowships #2017/09398-8, #2013/15040-8, #2018/24419-4, #2012/22488-2, #2013/19387-2, Research Grant 2015/04090-0 and CNPq. This work was also supported by Leducq Foundation, Award #FLQ13CVD04, and the European Union (GA#645648, “Muscle Stress Relief”).