
Abstract
Circular RNAs (circRNAs) and eukaryotic translation initiation factor 4A3 (EIF4A3) have been reported to participate in the pathogenesis of nasopharyngeal carcinoma (NPC), but their mechanism has not been fully understood. This research aimed to confirm the role and regulatory mechanism of hsa_circ_0049396 interacting with EIF4A3 in NPC tumorigenesis. Quantitative real time polymerase chain reaction (qRT-PCR) was executed to detect the levels of hsa_circ_0049396 and EIF4A3. Cell function experiments and nude mice xenograft assay were used to confirm the role of hsa_circ_0049396 in NPC. The regulatory effect of EIA4A3 on hsa_circ_0049396 was determined by circInteractome prediction, RNA binding protein immunoprecipitation (RIP) assay, and qRT-PCR. In addition, the Hippo–YAP pathway-related proteins and EIF4A3 protein were detected by western blotting. hsa_circ_0049396 was proved to be downregulated in NPC samples, and its low expression indicated the poor prognosis of NPC. After upregulating hsa_circ_0049396 in NPC cells, the proliferation, migration, invasion, and tumor growth in vivo were suppressed by inhibiting the Hippo–YAP pathway. Moreover, EIF4A3 bound to the flanking regions of the hsa_circ_0049396 to enhance hsa_circ_0049396 expression in NPC cells. hsa_circ_0049396 mediated by EIF4A3 in NPC can attenuate NPC tumorigenesis by inhibiting the Hippo–YAP pathway. This finding may provide a potential early diagnostic biomarker or drug target to improve the precision medicine approaches of NPC.
Introduction
Nasopharyngeal carcinoma (NPC) originating from nasopharyngeal epithelial cells is a malignant cancer with characterized geographical distribution, especially in South China (Chen et al., 2019). Despite improvements in the current diagnosis of NPC such as magnetic resonance imaging, Epstein-Barr virus DNA testing, and so forth, the therapeutic landscape of NPC remains challenging (Le et al., 2019, Su et al., 2023). Due to no obvious symptoms of NPC in early stage, some patients diagnosed with NPC have neck lymph node metastasis, thereby often occurring recurrence and metastasis (Jiang et al., 2020, Mo et al., 2022b, Mo et al., 2019). Hence, uncovering the molecular mechanism of NPC is beneficial to identify early diagnostic biomarkers, thereby enhancing precision medicine approaches of NPC.
Circular RNAs (circRNAs) with a closed-loop structure have emerged as critical players in the intricate landscape of gene regulation, exhibiting diverse cellular functions with implications in various cancers (Lei et al., 2020, Li et al., 2020a, Vo et al., 2019). In NPC, recent studies have implicated the function of circRNAs in the tumorigenesis. For examples, circ-ABCB10 upregulating ROCK1 could enhance growth and metastasis of NPC (Duan et al., 2020). Inversely, hsa_circ_0000345 upregulating PTEN suppressed cell metastasis in NPC (Jiang et al., 2022). A circRNA hsa_circ_0049396 formed by linear gene LDLR was reported to exert antitumor function in oral squamous cell carcinoma (Lou et al., 2023). However, the function and mechanism of hsa_circ_0049396 in NPC have not been revealed.
Eukaryotic translation initiation factor 4A3 (EIF4A3) is a pivotal component of the exon junction complex (EJC), using an RNA-binding protein to play a key role in post-transcriptional gene regulation and mRNA splicing (Kanellis et al., 2021). As we know, circRNA is formed by splicing from mRNA (Szabo and Salzman, 2016). Therefore, whether EIF4A3 can regulate circRNA expression attracts the attention of many researchers. EIF4A3 induced circSEPT9 to facilitate the carcinogenesis of triple-negative breast cancer (Zheng et al., 2020). EIF4A3 mediated the downregulation of circ_0087429 in cervical cancer and enhanced the tumorigenesis (Yang et al., 2022). In NPC, EIF4A3 was reported to promote circFIP1L1 formation, thereby repressing NPC cell malignancy and enhancing radiosensitivity (Zhou et al., 2022). However, the interaction between EIF4A3 and hsa_circ_0049396 has not been reported.
The Hippo signaling pathway, particularly its core co-activators the Yes-associated protein (YAP)/PDZ-binding motif (TAZ), has emerged as a central player in cancer development (Fang et al., 2022, Lee et al., 2021, Yan et al., 2022). Many studies demonstrate that the Hippo–YAP pathway can be the downstream of circRNAs to participate in tumorigenesis (Li et al., 2020b, Qadir et al., 2022, Zheng et al., 2019). For examples, hsa_circ_0128846 induced the inactivation of the Hippo–YAP pathway, thereby contributing to the tumorigenesis of colorectal cancer (Wang et al., 2020b). Another circRNA hsa_circ_0067741 was reported to impair the cell malignancy of lung adenocarcinoma by activating the Hippo–YAP pathway (Mo et al., 2022a). As for NPC, only one study confirmed that circRILPL1 activated the Hippo–YAP pathway to promote NPC progression (Wu et al., 2023). Here, bioinformatics analysis predicted that the Hippo–YAP pathway might be the downstream of hsa_circ_0049396. However, the correlation between the Hippo–YAP pathway and hsa_circ_0049396 has not been confirmed in any human diseases.
This study was committed to reveal the function and mechanism of hsa_circ_0049396 interacting with EIF4A3 and the Hippo–YAP pathway in NPC by a series of in vitro and in vivo experiments. Our study might provide a potential early diagnostic biomarker to improve the precision medicine approaches of NPC.
Materials and Methods
Clinical samples and cell lines
The NPC and adjacent normal tissues (>2 cm away from tumor) from 46 patients diagnosed with NPC in Puren Hospital between February 2016 and February 2023 were collected. All clinical samples were stored at −80°C and used with the approval of the ethics committee of Puren Hospital (approval number: A01401-LL202212-007). Written informed consents were obtained from all patients, and Table 1 together with Supplementary Data S1 outlines their clinical characteristics.
Clinical Characteristics of the Patients with NPC
NPC, nasopharyngeal carcinoma.
All cell lines including NPC cell lines (c666-1, BNCC337872; NPC/HK1, BNCC360149) and human nasopharyngeal epithelial cell line (NP69, BNCC359708) were purchased from BeNa Culture Collection (BNCC, China). NP69 cells were cultured in keratinocyte medium (BNCC) with 1% keratinocyte growth supplement (BNCC), whereas NPC cells were cultured in Roswell Park Memorial Institute (RPMI)-1640 medium (BNCC) with 10% fetal bovine serum (FBS) (BNCC). All cells were incubated at 5% carbon dioxide and 37°C.
Quantitative real time polymerase chain reaction (qRT-PCR)
VeZol Reagent purchased from Vazyme (China) was used to isolate RNA from tissues or cells. HiScript III RT SuperMix for quantitative PCR (Vazyme) was purchased to synthesize 1 μg RNA to cDNA, and HiScript II One Step qRT-PCR SYBR Green Kit (Vazyme) was purchased to perform quantitative reverse transcription PCR (qRT-PCR). Using 2−ΔΔCT method with GAPDH as an internal reference, the hsa_circ_0049396 expression was confirmed. Table 2 shows the primers sequences.
qRT-PCR Primers Used in This Study
RNase R treatment
Due to the RNase R resistance of circRNA, the cyclic structure of hsa_circ_0049396 was verified using RNase R. Briefly, RNase R (5 U/μg, R7092S, Beyotime, China) incubated 2 μg RNA at 37°C. After 30 min, qRT-PCR was employed to examine the levels of hsa_circ_0049396 and its linear gene LDLR according to the protocol mentioned above.
Cell transfection
hsa_circ_0049396/EIF4A3 overexpression vector (circRNA vector or EIF4A3 vector), siRNA targeting EIF4A3 (si-EIF4A3), and negative control of siRNA were constructed by OBiO Technology (China). The empty vectors were used as the negative control of overexpression vectors (empty vector). Then, the vectors mentioned above were transfected into NPC cells by OriFect Transfection Reagent (CC101, Oriscience, China). Finally, qRT-PCR was performed to confirm the transfection efficiency after 2 days cell transfection.
Cell proliferation detection
For evaluating NPC cell proliferation, Cell Counting Kit-8 (A311-01, CCK-8, Vazyme, China) was purchased. NPC cells (5 × 104) in 100 µL/well were seeded into 96-well plates. Incubation of cells occurred at 0, 24, 48, and 72 h; 10 µL/well of CCK-8 was added into NPC cells for 2 h incubation. A microplate reader (HM-96A, Hengmei, China) at 450 nm detected the optical density of each well to assess NPC cell proliferation.
Another assay called 5-Ethynyl-2’-deoxyuridine (EdU) assay was performed to further confirm NPC cell proliferation using YF594 Click-iT EdU Imaging Kit (YEASEN, China). NPC cells (5 × 104) in 100 µL/well were seeded into 96-well plates for 24 incubation. Then, 50 µL/well of 4% neutral paraformaldehyde was used to incubate NPC cells after removing cell medium. After 5 min, the NPC cells were washed using 0.1 mL/well of 3% bovine serum albumin, added 0.1 mL/well of 0.5% Triton X-100, and stained with 0.1 mL/well of Click-iT reaction mixture provided by the kit. After incubation for 30 min without light, the NPC cells were continued to incubate with 0.1 mL/well of Hoechst 33342 solution for 20 min without light. The images of NPC cells were photographed using a fluorescence microscope (Olympus, Japan).
Wound healing assay
NPC cells (1 × 105) after transfection seeded in 6-well plates were cultured until over 90% cell confluence. Then, sterilized yellow pipettes were used to scratch NPC cells. The images of NPC cell were photographed at 0 and 24 h. The migration rate was calculated by detecting the width of the wound to assess the ability of cell migration.
Transwell assay
Ceturegel Matrix LDEV-Free Matrigel (40183ES08, YEASEN) was used to incubate the upper chamber of transwell overnight at 4°C. Next day, the transfected NPC cells were added into the upper chamber with the serum-free medium, as well as the medium with 20% FBS were added to the top chamber. After 36 h, the NPC cells invading the top chamber were observed after fixing and staining.
Nude mice xenograft assay
Ten BALB/C nude mice (male, about 4 weeks old) from Shulaibao Biotechnology Co., Ltd. (China) were divided into two groups: empty vector group and circRNA vector group. Lentivirus hsa_circ_0049396 overexpression vector (circRNA vector) and its empty vector as negative control were constructed by OBiO Technology (China) and transfected into c666-1 cells for stably overexpressed hsa_circ_0049396. Then, the c-666 cells were subcutaneously injected into mice according to the different groups. Every week, tumor volume measurements were conducted until 5 weeks. At the 5th week, euthanization of mice were performed, and the tumor was removed from mice for measuring tumor weight. The ethics committee of Puren Hospital approved this animal experiment (approval number: 2022009). The experiment was repeated three times.
Bioinformatic analysis
Circular RNA Interactome, another circInteractome, is often used to predict miRNAs binding to circRNA. Here, the miRNAs binding to hsa_circ_0049396 was firstly predicted by circInteractome. Then, miEAA 2.1, a tool of miRNA enrichment analysis, was used to perform Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment for the predicted miRNAs mentioned above. In addition, circInteractome was also used to predict the RNA binding proteins of hsa_circ_0049396.
Western blotting
Protein extraction was carried out by purchasing Radio Immunoprecipitation Assay (RIPA) lysis buffer (20101ES60, YEASEN), and the quantification of proteins was performed by purchasing BCA Protein Quantification Kit (20201ES76, YEASEN). Subsequently, the 20 μg of protein samples that underwent separation on sodium dodecyl sulphate-polyacrylamide gel electrophoresis (SDS‐PAGE) were transferred onto a polyvinylidene fluoride (PVDF) membrane using semi-dry method, blocked by soaking in 5% nonfat milk, and incubated with primary antibodies including Phospho-MST1 (p-MST1, 49332, Cell Signaling Technology [CST], China), MST1 (3682, CST), Phospho-LATS1 (p-LATS1, 8654, CST), LATS1 (3477, CST), YAP1 (14074, CST), EIF4A3 (KMA0226669R, KeMiao Biological Technology, China), and GAPDH (2118T, CST) at 4°C overnight. After that, anti-rabbit IgG (H + L) (14708S, CST) was used to incubate the membranes for 3 h. Finally, Super ECL Detection Reagent (36208ES60, YEASEN) was purchased to visualize protein.
RNA binding protein immunoprecipitation (RIP)
EZ-Magna RIP kit (17-704, Millipore, USA) was purchased to perform RIP assay to confirm the interaction between EIF4A3 and hsa_circ_0049396. The NPC cells were lysed in RIP buffer and subjected to incubation with magnetic beads with anti-EIF4A3 or anti-IgG. Finally, the immunoprecipitated RNAs were isolated and subsequently analyzed via qRT-PCR.
Statistical analysis
Data were analyzed using Graphpad Prism and displayed as mean ± standard deviation from three independent experiments. Paired or unpaired t-test and analysis of variance were applied to compare the difference between two groups and over two groups with p < 0.05 as the significant difference, respectively.
Results
hsa_circ_0049396 is downregulated in NPC
qRT-PCR verified the downregulation of hsa_circ_0049396 in NPC samples from 46 patients with NPC (Fig. 1A). Then, the average expression of hsa_circ_0049396 was used to divide the patients to high expression group and low expression group. Using Log-rank (Mantel–Cox) test, hsa_circ_0049396 with low expression indicated the poor prognosis in patients with NPC (Fig. 1B). In NPC cells (c666-1 and NPC/HK1), hsa_circ_0049396 was proved to be downregulated by >50% compared with NP69 cells (Fig. 1C). The construction of hsa_circ_0049396 was shown in Figure 1D, and LDLR was the linear gene of hsa_circ_0049396. RNase R treatment assay confirmed the cyclic structure of hsa_circ_0049396 due to the resistance of hsa_circ_0049396 to RNase R (Fig. 1E). The above data demonstrated that hsa_circ_0049396 with cyclic structure was downregulated in NPC.
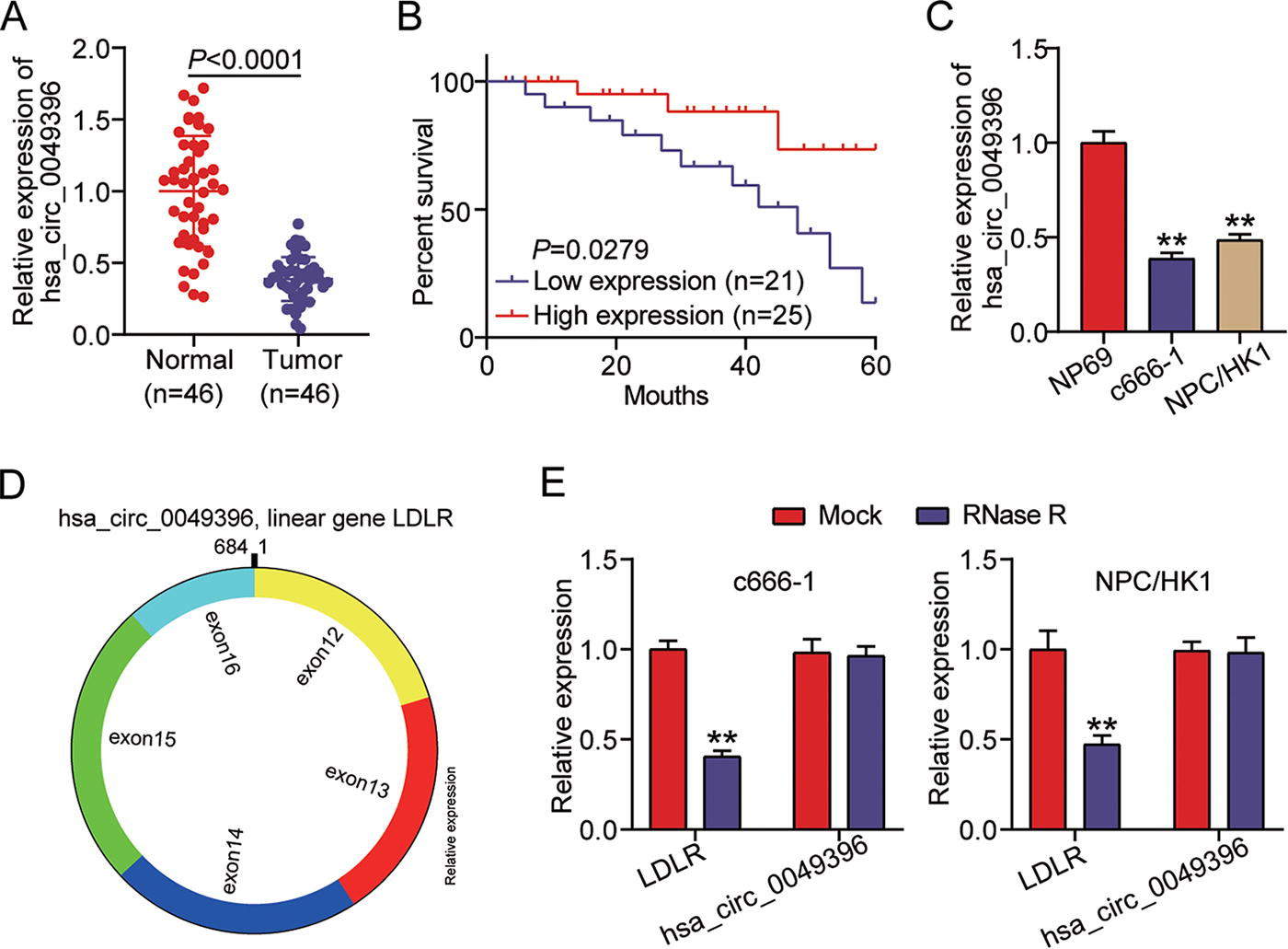
hsa_circ_0049396 is downregulated in NPC.
hsa_circ_0049396 overexpression attenuates the NPC tumorigenesis in vitro
To assess the role of hsa_circ_0049396 in NPC, the hsa_circ_0049396 overexpression vectors were used to transfect NPC cells, which led to 8.5–10.2-fold hsa_circ_0049396 upregulation in NPC cells (Fig. 2A). Then, CCK-8 and EdU assays assessed the changes of NPC cell proliferation, showing that hsa_circ_0049396 overexpression inhibited NPC cell proliferation (Fig. 2B, C). Using wound healing assay to confirm the changes of NPC cell migration, it was found that hsa_circ_0049396 overexpression reduced 21–29% of migration rate in NPC cells (Fig. 2D). As for cell invasion detected by transwell assay, the number of invaded NPC cells was proved to be reduced by 63–70% when hsa_circ_0049396 was overexpressed (Fig. 2E). These data proved that enhancing hsa_circ_0049396 expression attenuated the NPC.
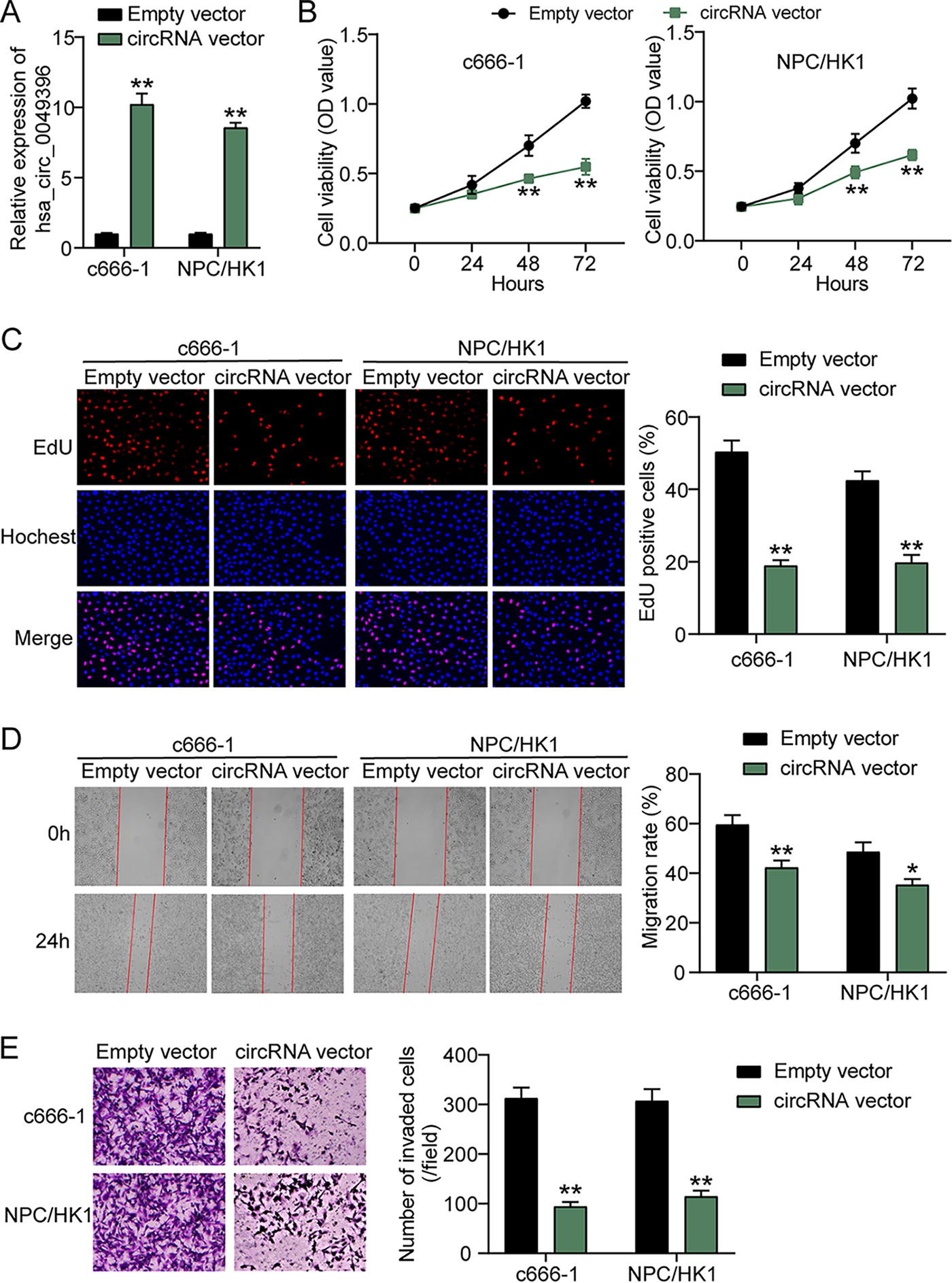
hsa_circ_0049396 overexpression attenuates the tumorigenesis of NPC cells in vitro.
hsa_circ_0049396 overexpression impairs tumor growth of NPC cell in vivo
Subsequently, the effect of hsa_circ_0049396 on NPC cells was verified using nude mice xenograft assay. The data indicated that hsa_circ_0049396 overexpression declined tumor volume (Fig. 3A), tumor size (Fig. 3B), and tumor weight (Fig. 3C). These results confirmed that the tumor growth of NPC cell could be suppressed by hsa_circ_0049396 overexpression in vivo.
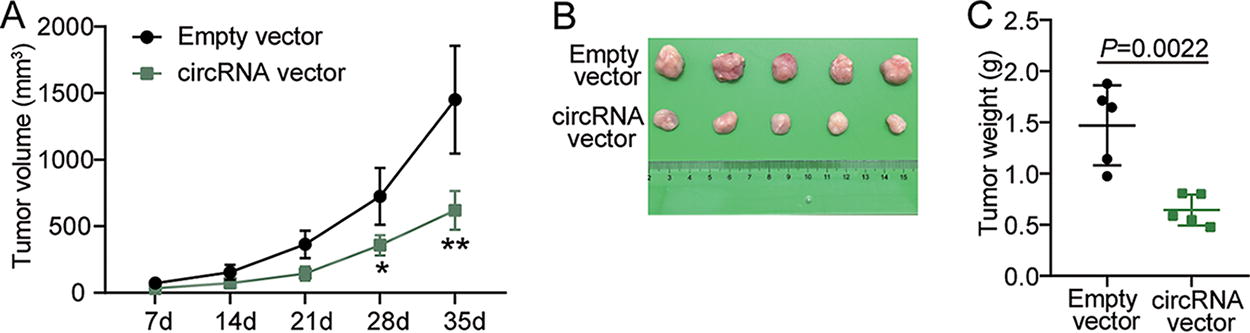
hsa_circ_0049396 overexpression impairs tumor growth of NPC cell in vivo.
hsa_circ_0049396 overexpression induces the inactivation of the Hippo–YAP pathway in NPC cells
To confirm how hsa_circ_0049396 regulated the tumorigenesis of NPC cells, we used circInteractome to predict the miRNAs binding to hsa_circ_0049396 and used miEAA 2.1 to perform KEGG enrichment for predicted miRNAs. The results showed that the Hippo signaling pathway might be the downstream pathway of hsa_circ_0049396 (Fig. 4A). Western blotting further detected the key protein levels of the Hippo signaling pathway in NPC cells transfected with hsa_circ_0049396 overexpression. As shown in Figure 4B, hsa_circ_0049396 overexpression upregulated the levels of p-MST1 and p-LATS1 by around 1.5-fold, but it downregulated the level of YAP1 by around 43% in NPC cells. Taken together, the upregulation of hsa_circ_0049396 in NPC cells suppressed the activation of the Hippo–YAP pathway.
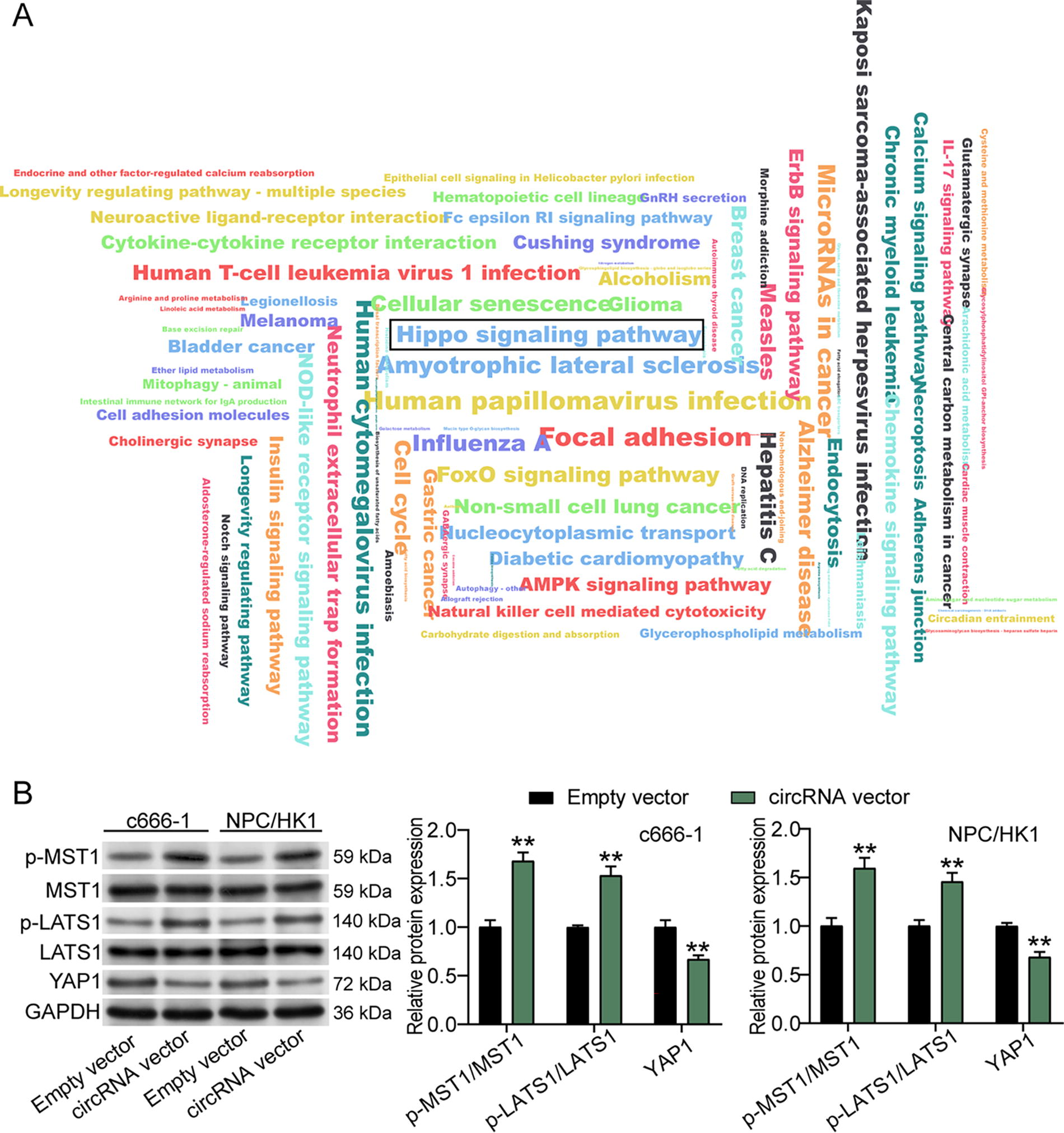
hsa_circ_0049396 overexpression induces the inactivation of the Hippo–YAP pathway in NPC cells.
EIF4A3 mediates the hsa_circ_0049396 upregulation in NPC cells
To investigate the cause of the downregulation of hsa_circ_0049396 in NPC cells, circInteractome predicted the RNA binding proteins of hsa_circ_0049396, of which EIF4A3 was predicted to have the most binding sites on the flanking regions of the hsa_circ_0049396 (Fig. 5A). Then, RIP assay further proved that the flanking regions of the hsa_circ_0049396 contained four binding sites for EIF4A3 (Fig. 5B). After transfecting si-EIF4A3 and EIF4A3 overexpression vector into NPC cells, western blotting revealed that the si-EIF4A3 reduced 40% of EIF4A3 expression and EIF4A3 overexpression vector induced almost 1.5-fold upregulation of EIF4A3 expression (Fig. 5C). As for the effects of EIF4A3 on hsa_circ_0049396, qRT-PCR displayed that si-EIF4A3 induced 50% downregulation of hsa_circ_0049396 overexpression, whereas EIF4A3 overexpression vector induced around 1.8–2.1-fold upregulation of hsa_circ_0049396 expression (Fig. 5D). These data suggested that EIF4A3 in NPC cells could induce the upregulation of hsa_circ_0049396.
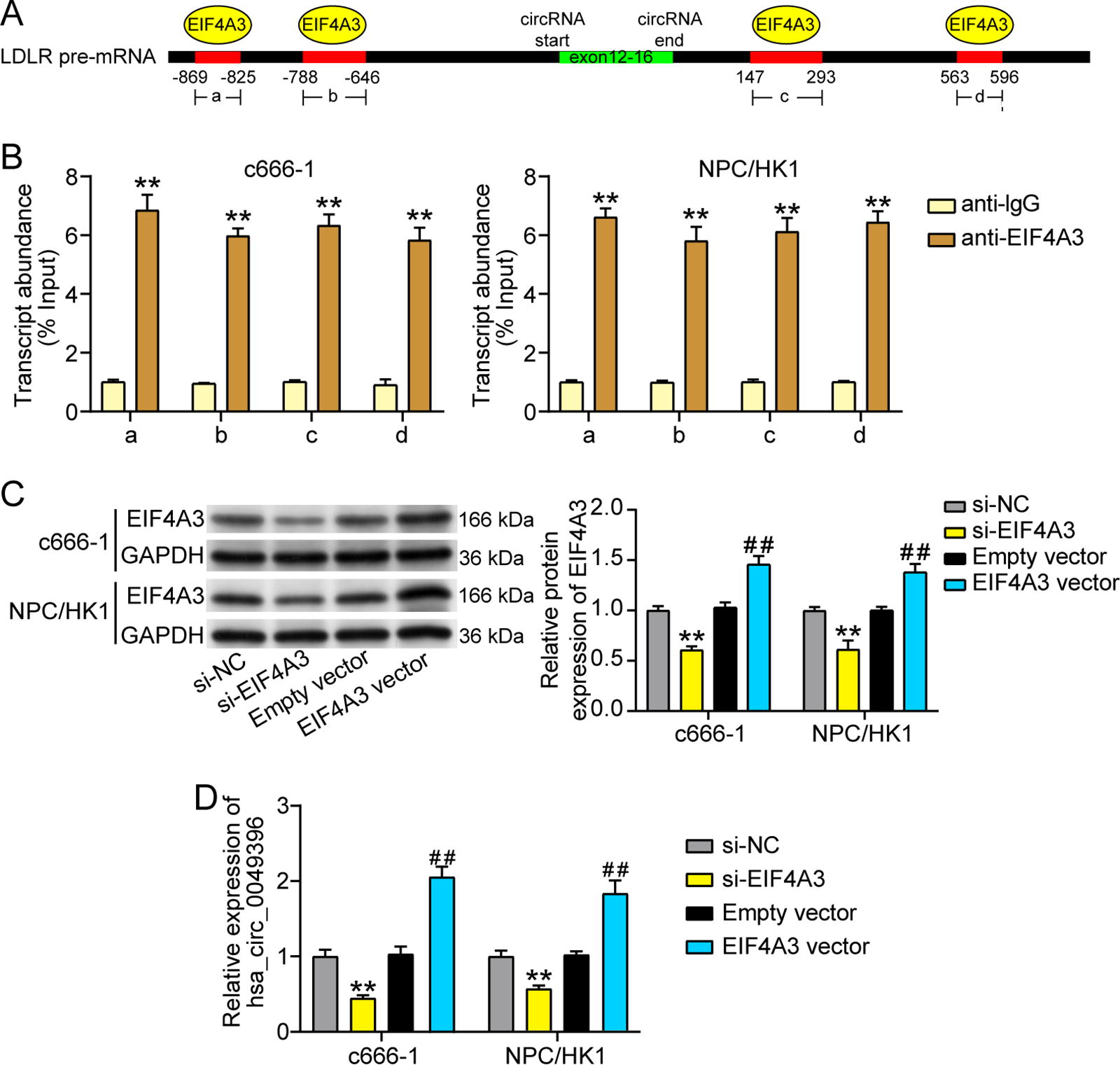
EIF4A3 mediates the hsa_circ_0049396 upregulation in NPC cells.
Discussion
CircRNAs with stable closed-loop structure are key regulators in tumor development (Leng et al., 2021, Sun et al., 2020, Wu et al., 2021). This study found hsa_circ_0049396 with low expression in NPC, and overexpressing hsa_circ_0049396 was able to inhibit NPC cell proliferation, migration, invasion, and tumor growth. After further exploring the regulatory mechanism on hsa_circ_0049396 in NPC, this study revealed that hsa_circ_0049396 expression could be induced by EIF4A3 to inhibit the Hippo–YAP pathway, thereby exerting antitumor function in NPC. These findings prove the key regulatory function of hsa_circ_0049396 in NPC, which may provide a potential early diagnostic marker to improve the NPC treatment.
Increasing evidences prove the key regulatory function of circRNAs in the tumorigenesis of NPC. For example, circ_0000215 with high expression in NPC was proved to exert oncogenic function by interacting with miR-512-5p (Chen et al., 2021). Jiang et al. used cell experiments to reveal that hsa_circ_0000345 overexpression impaired the malignancy of NPC cell by interacting with miR-513a-3p/PTEN axis (Jiang et al., 2022). Here, we firstly found the downregulation of hsa_circ_0049396 in NPC and upregulating hsa_circ_0049396 in NPC could efficaciously decline the abilities of proliferation, migration, invasion, and tumor growth. After checking the literatures, only one study confirmed the antitumor role of hsa_circ_0049396 in oral squamous cell carcinoma (Lou et al., 2023). Therefore, our study is the first to reveal the function of hsa_circ_0049396 in NPC.
Increasing studies confirm the key regulatory role of the Hippo–YAP signaling pathway in the cancer development (Fang et al., 2022, Lee et al., 2021, Yan et al., 2022). According to the previous studies, YAP only in the nucleus can be a co-activator to accelerate the tumorigenesis, but the phosphorylation of YAP prevents YAP translocation into the nucleus (Barry et al., 2013, Zhao et al., 2011). As the upstream of YAP, the phosphorylation of LATS1/2 by phosphorylated MST1 can phosphorylate YAP, thereby inducing YAP destabilization and preventing YAP entry into the nucleus (Kwan et al., 2016, Li et al., 2022, Wang et al., 2020a). In NPC, the Hippo–YAP pathway as a key signaling pathway has been reported in many researches. Chen et al. found that the inactivation of the Hippo–YAP pathway caused by alisol A could suppress the NPC cell to proliferate, migrate, and invade (Chen and Liu, 2021). Another study reported that circRILPL1 accelerated the NPC progression by mediating the activation of the Hippo–YAP pathway (Wu et al., 2023). However, there is no study to confirm the correlation between the Hippo–YAP pathway and hsa_circ_0049396 in any human diseases. This research innovatively discovered that enhancing hsa_circ_0049396 could upregulate the levels of phosphorylated MST1 and LATS1, as well as downregulate the level of YAP, illustrating that hsa_circ_0049396 inhibits the activation of the Hippo–YAP pathway in NPC. Therefore, this study confirmed that hsa_circ_0049396 impaired the NPC cell malignancy by inhibiting the Hippo–YAP pathway.
Many studies reveal the mechanism of circRNAs in tumorigenesis of NPC from the direction of competitive endogenous RNAs mechanism (Huang et al., 2022, Wang et al., 2021a, Yin et al., 2020, Zeng et al., 2023). This study explored the regulatory mechanism of hsa_circ_0049396 in NPC from the direction of RNA-binding protein. Using molecular biology experiment together with circInteractome prediction, EIF4A3 was proved to interact with the flanking regions of the hsa_circ_0049396 in NPC, thereby enhancing hsa_circ_0049396 expression. EIF4A3, a pivotal component of EJC, has been reported to participate in tumorigenesis by regulating circRNA expression. For example, EIF4A3 bound to the flanking regions of hsa_circ_0068631 to enhance the levels of hsa_circ_0068631, thereby promoting the progression of breast cancer (Wang et al., 2021b). EIF4A3 bound to the flanking regions of circ_0087429 to suppress its expression, leading to the damage of cervical cancer tumorigenesis (Yang et al., 2022). The similarity research was also reported in NPC, showing that EIF4A3 promoted circFIP1L1 by binding to its flanking regions to repress NPC cell malignancy and enhanced radiosensitivity (Zhou et al., 2022). In contrast from the previous study, this research demonstrated that hsa_circ_0049396 expression was enhanced by EIF4A3 in NPC to inhibit NPC progression.
From the discussion above, this study confirmed the function and mechanism on hsa_circ_0049396 in NPC. However, there are some certain important limitations that need to be solved in the future. First, this research confirmed that hsa_circ_0049396 suppressed the Hippo–YAP pathway to inhibit NPC progression. However, how hsa_circ_0049396 regulates the phosphorylation levels of MST1 and LATS1 has not been deeply explored. In addition, whether there is a drug targeting hsa_circ_0049396 to regulate NPC progression needs to be further investigated.
Conclusion
This study discovers that EIF4A3 can mediate hsa_circ_0049396 expression in NPC by binding to its flanking regions, and overexpressing hsa_circ_0049396 impairs the NPC tumorigenesis by inhibiting the Hippo–YAP pathway. The findings of this study suggest that hsa_circ_0049396 may serve as a potential diagnostic biomarker or drug target in NPC to improve the precision medicine approaches of NPC.
Footnotes
Authors’ Contributions
Q.Z. conducted the experiments, analyzed all data, and wrote this article. B.C. designed this research and collected the samples. K.L. performed the interpretation of data. H.C. edited this article. All authors reviewed and approved this article.
Ethics Approval
The research received approval from the ethics committee of Puren Hospital Affiliated to Wuhan University of Science and Technology in Wuhan, China. Clinical tissue specimen processing adhered strictly to the ethical standards outlined in the Declaration of Helsinki, with each patient providing signed consent forms. The animal experiment, also approved by the ethics committee of Puren Hospital, was conducted in accordance with the Animal Research: Reporting of In Vivo Experiments guidelines.
Data Availability
The data in the current study can be obtained from the corresponding author upon reasonable request.
Disclosure Statement
The authors declared that there are no conflicts of interest.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Data S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.