
Abstract
Abstract
Kinetics of low nitrate and ammonia concentration removal by oligotrophic Acinetobacter sp. SYF26 were examined in this study. Heterotrophic nitrification–aerobic denitrification followed first-order reaction kinetics. The maximum removal rate was achieved under pH = 7.0, 30°C, and a carbon-to-nitrogen ratio (C/N) = 7.5. In a landscape water experiment, strain SYF26 was capable of both heterotrophic nitrification and aerobic denitrification and exhibited no nitrite accumulation during the whole process. Both three-dimensional excitation–emission matrix fluorescence spectroscopy and Fourier transform near-infrared spectroscopy assessments of landscape water illustrated that strain SYF26 could use protein-like compounds and humus as carbon sources for heterotrophic nitrification–aerobic denitrification. Results illustrated that strain SYF26 is a suitable candidate for the bioremediation of landscape water.
Introduction
A
During the last several decades, an increasing number of studies have focused on new biological nitrogen removal technologies. Several heterotrophic nitrifying–aerobic denitrifying bacteria had been reported, including Bacillus sp. (Kim et al., 2005), Providencia rettgeri (Taylor et al., 2009), P. stutzeri (Zhang et al., 2011), and Bacillus methylotrophicus L7 (Zhang et al., 2012). These microorganisms could be utilized to reduce the costs associated with maintaining an anoxic tank or to reduce the size of the tank required. The mechanism of nitrogen removal by certain heterotrophic nitrifying–aerobic denitrifying bacteria was NH4+→NH2OH→NO2−→NO3− and then NO3−→NO2−→N2. Therefore, nitrogen removal by heterotrophic nitrification and aerobic denitrification bacteria has gained greater attention in wastewater treatment (Chen et al., 2012).
Recently, the heterotrophic nitrification–aerobic denitrification mainly focused on substrate removal and accumulation of intermediate product (Wan et al., 2011). However, the research on the characteristics of denitrification using nitrite was rarely reported (Wan et al., 2011). Furthermore, although a number of heterotrophic nitrification–aerobic denitrification bacteria have been reported, the heterotrophic nitrification–aerobic denitrification bacteria with low nitrogen source concentration (total nitrogen [TN] <5 mg/L) have been reported rarely. Meanwhile, kinetic analysis is a useful tool to evaluate the heterotrophic nitrification–aerobic denitrification process. Therefore, the kinetic evaluation of nitrogenous substances removal was applied by many researches (An et al., 2010; Bi et al., 2015).
In the present study, the strain Acinetobacter sp. SYF26 (GenBank accession No. LC010332) was isolated from the Hei He oligotrophic drinking-water reservoir in China. Strain SYF26 was characterized based on its heterotrophic nitrifying–aerobic denitrifying performance using ammonia, nitrite, and nitrate as substrates under low-nutrient conditions. The first-order reaction kinetics equation was used to model the kinetic processes of heterotrophic nitrification–aerobic denitrification. The effects of pH, temperature, and carbon-to-nitrogen (C/N) ratio on ammonium and nitrate removal were investigated. Furthermore, heterotrophic nitrification–aerobic denitrification was characterized in the landscape water by oligotrophic Acinetobacter sp. SYF26. Changes in dissolved organic materials were examined in the landscape water after the addition of strain SYF26 by three-dimensional excitation–emission matrix (3D-EEM). In addition, Fourier transform infrared spectroscopy (FTIR) was used to analyze changes in the composition of substances in landscape water after the addition of strain SYF26.
Materials and Methods
Strain and culture media
Strain Acinetobacter sp. SYF26 (GenBank accession No. LC010332) (Su et al., 2013) was isolated from the sediment of Hei He reservoir, Xi'an, China. The strain SYF26 was grown in a modified medium (MM), and the MM had two forms as follows: MM1 and MM2. The components of the MM1 were as follows per liter: CH3COONa, 0.0605 g; NaNO3, 0.0121 g; K2HPO4, 0.0121 g; MgCl2, 0.0061 g; and CaCl2, 0.0061 g. The components of the MM2 were as follows per liter: CH3COONa, 0.0380 g; NH4Cl, 0.0076 g; K2HPO4, 0.0076 g; MgCl2, 0.0038 g; and CaCl2, 0.0038 g. The components of trace element solution were as follows per liter: MgSO4·7H2O, 0.5 g; EDTA, 1.0 g; ZnSO4, 0.2 g; MnCl2·4H2O, 0.1 g; FeSO4·7H2O, 0.5 g; CuSO4·5H2O, 0.5 g; and CoCl2·6H2O, 0.2 g. The final pH of the medium was adjusted to 7.0 by 1 mol/L NaOH or HCl solution.
Assessment of heterotrophic nitrification and aerobic denitrification efficiency at low nitrate and ammonia concentration by strain SYF26
To assess how initial pH, temperature, and C/N ratio affected the heterotrophic nitrification and aerobic denitrification efficiency at low nitrate and ammonia concentration, the single-factor experiments were carried out in flasks using the MM1 and MM2, respectively. The MM1 and MM2 corresponding supplied with 2 mg/L of NO3-N(MM1) or NH4−-N(MM2) as a sole nitrogen source, respectively, and the CH3COONa as a sole carbon source. In the pH experiment, the initial pH was adjusted to 5, 6, 7, 8, and 9 by the addition of 1 mol/L HCl or 1 mol/L NaOH, meanwhile, the temperature and C/N were 30°C and 6°C, respectively. To determine the effect of temperature, the temperatures were set at 15°C, 20°C, 25°C, 30°C, and 35°C, respectively, and the pH and C/N were 7 and 6. In the C/N experiment, the amount of sodium acetate was changed to adjust the C/N ratios in the MM1 or MM2 by maintaining a constant nitrate or ammonia nitrogen concentration at 2 mg/L. Finally, the C/N ratio was adjusted to 3, 4.5, 6, 7.5, and 9, respectively. Meanwhile, the pH and temperature were 7°C and 30°C. All the single-factor experiments were conducted with the inoculation size of 1% (v/v) in 250-mL flasks with 100 mL sterile medium, and the shaking speed was adjusted to 120 rpm in this study.
Application in landscape water
To evaluate the efficiency of heterotrophic nitrification and aerobic denitrification, 1% (v/v) inoculum of sample SYF26 was added to 250-mL conical flask with 200 mL landscape water (34.202534°N–34.211416°N, 108.989245°E– 108.993611°E). All the samples were collected from the middle of the conical flask, and then, all the samples were filtered using cellulose acetate syringe filters with a pore size of 0.45 nm before the measurements of ammonium, nitrate, nitrite, TN, and total organic carbon (TOC) concentrations. All the samples were measured from day 0 to 5.
Fluorescence assessment of landscape water components
Both samples C1 and C2 were collected at first day and fifth day after addition of strain SYF26, and they come from landscape water. The 3D-EEM fluorescence spectra can be used to measure the change of dissolved organic matter in landscape water and measurements of 3D-EEM fluorescence spectra were obtained using a fluorescence spectrophotometer (FP-6500; Jasco) equipped with a xenon lamp source, excitation and emission monochromators, and a front-face sample-cell holder. The emission wavelength was set from 250 to 550 nm by varying in 10 nm increments, while the excitation wavelength was set from 220 to 450 nm at 5 nm increments. Landscape water substances were compared on the first and fifth days to demonstrate the heterotrophic nitrification–aerobic denitrification efficiency by strain SYF26.
Fourier transform near-infrared spectroscopy
Both samples C1 and C2 were collected at first and fifth day after addition of strain SYF26, and they come from landscape water. Each sample was dried at 110°C for about 10 h. The solid sample was grounded into powder and then grounded again and pressed into a tablet. For each sample, three spectra of the same film were recorded from an accumulation of 16 successive scans in the range of 4,000–500 cm−1 with a resolution of 4 cm−1 to assess the reproducibility. The background scan was always conducted with a golden slit before acquiring the spectrum from each sample. All spectral data were acquired using an FT-NIR spectrometer (Antaris II FT-NIR analyzer; Thermo Scientific Co.). In this research, the FTIR can be used to measure the change of organic matter in landscape water, furthermore, it can distinguish different functional groups and their changes.
Analytical methods
The culture samples were centrifuged at 10,000 rpm and filtered through 0.45 μm membrane filter. TOC was measured by a TOC analyzer (Jena Multi N/C). NO2−-N level was determined by N-(1-naphthyl)-1, 2-diaminoethane dihydrochloride spectrophotometry (Mahmood et al., 2009). NH4+-N level was determined by Nessler assay (Zhang, 2009). TN content was determined by peroxydisulfate oxidation with a UV spectrophotometric method (Ebina et al., 1983). NO3−-N level was determined by ultraviolet spectrophotometric method (APHA, 1998).
Statistical analysis
NH4+-N, NO2−-N, NO3−-N, TN, and TOC removal ratio and removal rate formula were 100 × (Ci − Cf)/Ci and (Ci − Cf)/t, respectively, where Ci was the initial concentration (mg/L), Cf was the final concentration (mg/L), and t was the time (hour). The statistical method was as follows: The data presented are the mean and standard deviation of the triplicate experiments. Statistical analysis and graphical work were carried out by using Excel and Origin 9.0.
Results and Discussion
Kinetic analysis of heterotrophic nitrification–aerobic denitrification by strain SYF26
The following first-order reaction kinetics equation was to model kinetic processes of heterotrophic nitrification–aerobic denitrification.
Taking the logarithm of both the sides of equation, the following is obtained:
where C represents the ammonia or nitrate or nitrite concentration (mg/L); t is the time (hour) of the heterotrophic nitrification or aerobic denitrification reaction; k is the heterotrophic nitrification or aerobic denitrification reaction rate (mg/[L·h]) constant, and b is a constant.
Tables 1 and 2 illustrate that Y = Ln (C) and t have a good linear relationship, with most correlation coefficients R2 exceeding 0.90, conforming to the first-order reaction kinetics equation.
C/N, carbon-to-nitrogen ratio.
Heterotrophic nitrification–aerobic denitrification ability of strain SYF26 under various initial pH conditions when NO3−-N (2.03 mg/L) or NH4+-N (2.09 mg/L) was provided as the sole nitrogen source is shown in Fig. 1. The data in Figure 1 is also available as Supplementary Data online at www.liebertpub.com/ees. As demonstrated in Fig. 1a and b, different heterotrophic nitrification or aerobic denitrification rates were obtained at different initial pH values. For aerobic denitrification, the nitrate removal rate was greater at pH 7, 8, and 9 than at pH 5 or 6, suggesting that low pH values had affected the activity levels of certain enzymes and thereby reduced the removal rate. However, in heterotrophic nitrification, a relatively high ammonia removal rate was obtained in all pH conditions, except for pH = 5. Furthermore, both the maximum nitrate and ammonia removal rates were observed at pH = 8, and the highest removal rates were 0.0184 and 0.0189 mg/[L·h], respectively. Single-factor experiments have suggested that the optimal pH for heterotrophic nitrification by strain B. methylotrophicus L7 is pH 7–8 (Zhang et al., 2012). The optimal running pH for the MBR was pH = 7.5 by a heterotrophic nitrifying–aerobic denitrifying bacterial strain (Yao et al., 2013a). In general, most heterotrophic nitrification–aerobic denitrification bacteria prefer neutral or slightly alkaline environments (Zheng et al., 2012).
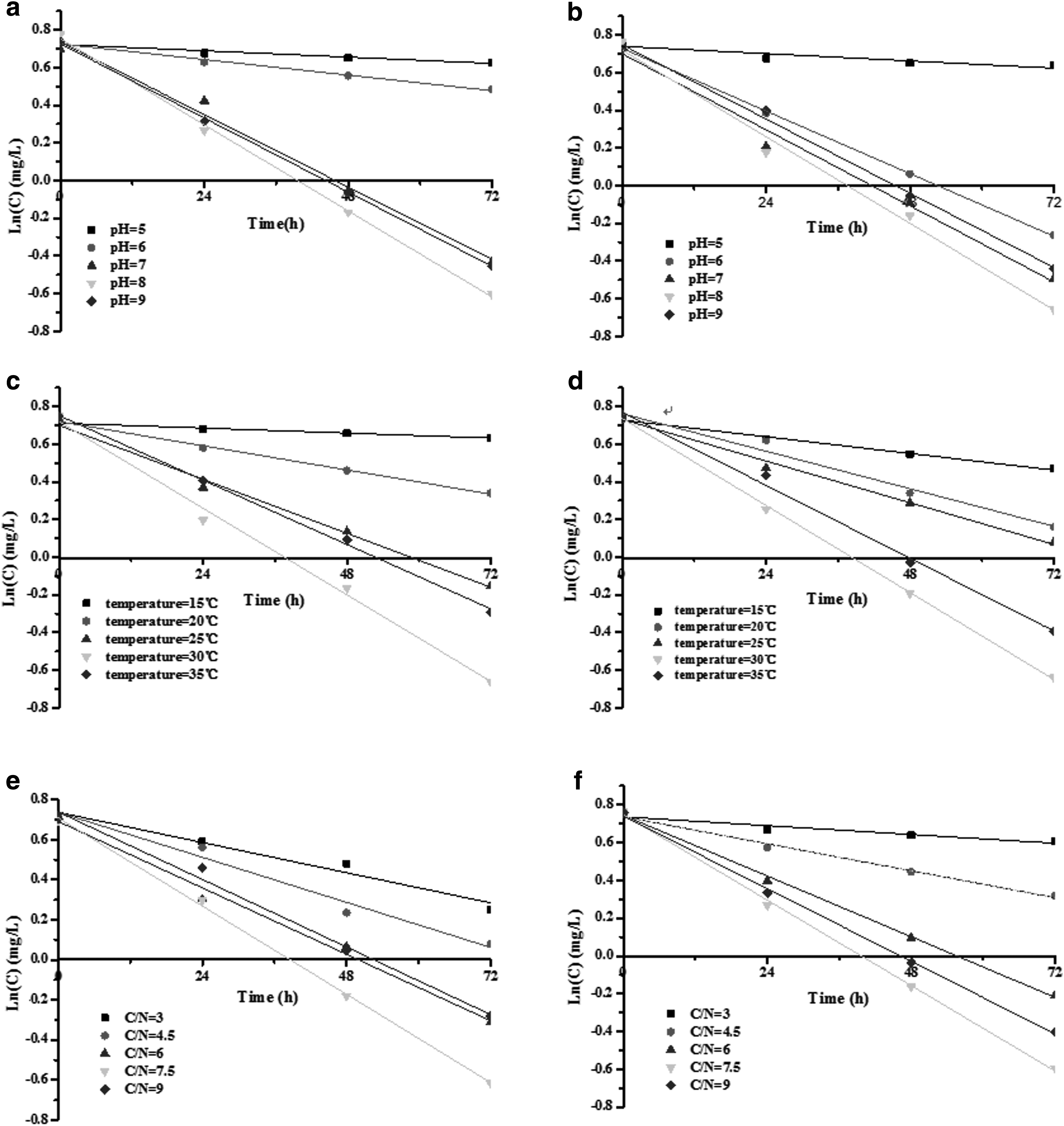
Removal rates for low nitrate or ammonia concentrations under different temperatures are shown in Fig. 1c and d. Strain SYF26 had different nitrate or ammonia removal rates depending on temperature. As shown in Fig. 1c and d, aerobic denitrification or heterotrophic nitrification capacities were optimal at 30°C, and nitrate or ammonia removal rate was 30°C > 35°C > 25°C > 20°C > 15°C. The highest nitrate and ammonia removal rates were 0.0192 and 0.0193 mg/[L·h], respectively. Furthermore, a nearly identical removal rate was achieved at 25°C and 35°C for aerobic denitrification. However, for heterotrophic nitrification, the removal rate at 35°C was significantly greater than that at 25°C. The removal rate increased as the temperature increased from 15°C to 30°C, and further increases in temperature (to above 30°C) did not result in further improvements in the nitrate removal rate. Although the removal rate in 35°C was slightly higher than in 25°C, there was no statistically significant difference between the results. However, the removal rate in 30°C was obviously higher than in 35°C, 25°C, 20°C, and 15°C, and there were significant differences between 30°C and 20°C and 30°C and 25°C (p < 0.05), respectively. Temperature is among the external factors that play a significant role in determining bacterial growth, and normal metabolism can only occur within a certain temperature range. The Su et al. (2017) showed that the effect of heterotrophic nitrification of stain H117 gradually increased with increasing temperature from 25°C to 30°C in the HM, and then gradually decreased above a temperature of 30°C. The bacterium P. rettgeri YL exhibits an unusual ability to heterotrophically nitrify and aerobically denitrify various concentrations of ammonium (NH4+-N), and the optimal temperature was 30°C (Taylor et al., 2009). P. stutzeri YZN-001 can utilize not only nitrate and nitrite but also ammonium, and at 30°C, the utilization of ammonium is ∼95% in 18 h (Zhang et al., 2011). The optimal temperature for heterotrophic nitrification–aerobic denitrification by strain HN-02 is 30°C (Chen et al., 2014).
As shown in Fig.1e and f, the effects of C/N on the low nitrate or ammonium concentration removal rate were also investigated. For C/N ratios of 3–7.5, the nitrate or ammonia removal rate increased significantly as the C/N ratio increased. However, this rate decreased when C/N ratio increased from 7.5 to 9. The optimal removal rates of nitrate and ammonia were obtained at C/N = 7.5, and the highest removal rates were 0.0191 and 0.0192 mg/[L·h], respectively. The bacterium P. stutzeri YG-24 exhibited efficient heterotrophic nitrification and aerobic denitrification. Duan et al. (2015) reported that C/N = 10 is the optimal condition for ammonia removal by strain SF16.
Based on the above kinetic experiments, not only for low nitrate concentrations but also for low ammonia concentrations, strain SYF26 could effectively remove ammonium and nitrate. Furthermore, the optimum conditions were pH = 8, 30°C, and C/N = 7.5.
Assessment of efficiency of heterotrophic nitrification–aerobic denitrification in landscape water by strain SYF26
As shown in Fig. 2, strain SYF26 exhibited a good removal efficiency of NH4+-N, NO2−-N, NO3−-N, TN, and TOC throughout the experiment. NH4+-N decreased from 1.92 to 0.25 mg/L and the removal efficiency was 86.98% with a removal rate of 0.014 mg/[L·h] for the whole process. The NO3−-N concentration decreased significantly from 0 to 96 h and remained relatively stable after 96 h, indicating that the TOC was sufficient for denitrification from 0 to 96 h in landscape water. In addition, the NO3−-N removal ratio was 98.75%, with a removal rate of 0.013 mg/[L·h]. Strain SYF26 could reduce the TN concentration from 3.53 to 0.29 mg/L in 96 h, and the removal efficiency was as high as 91.78%, with a removal rate of 0.034 mg/[L·h].
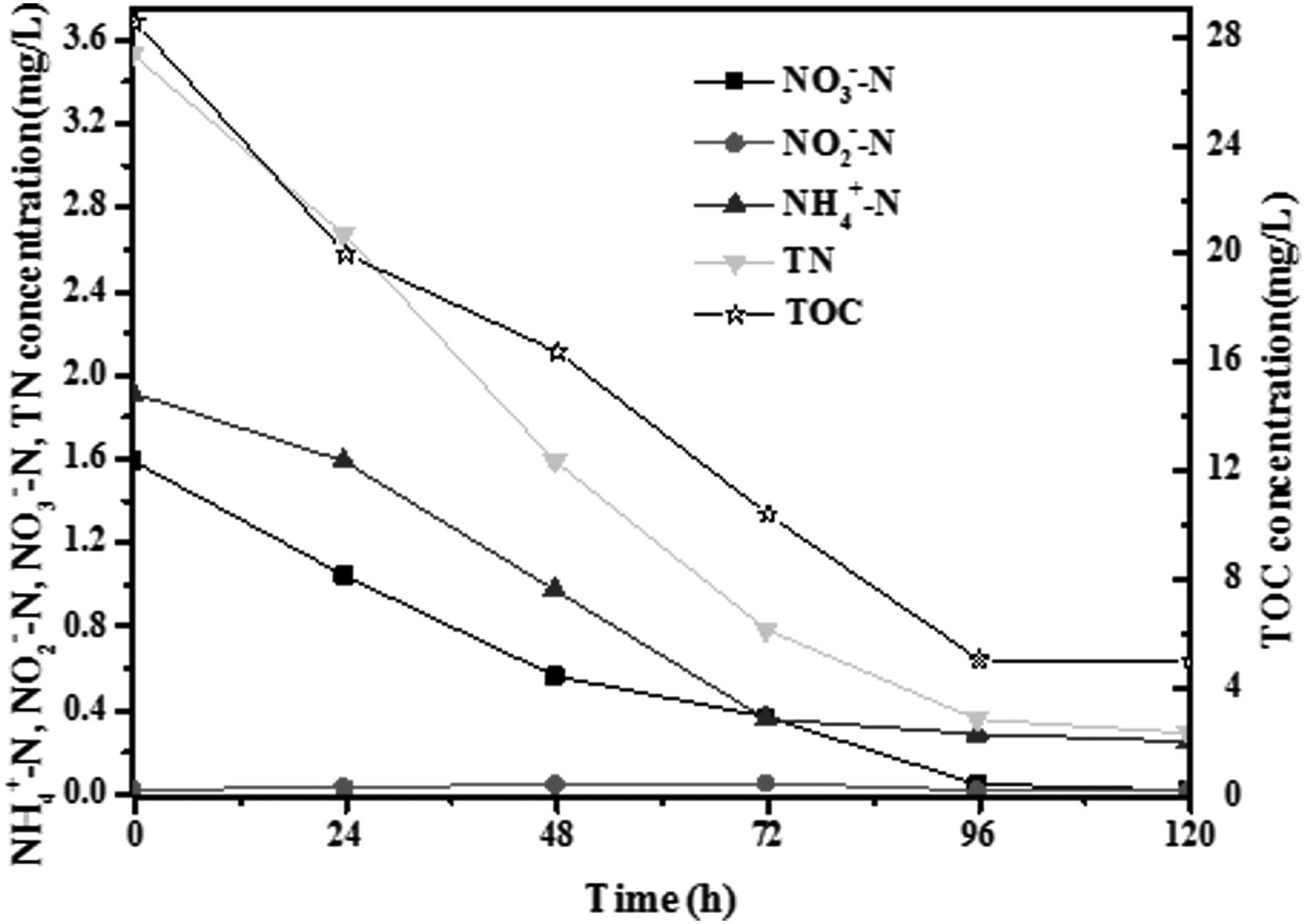
Changes of NH4+-N, NO2−-N, NO3−-N, TN, and TOC by strain SYF26 in the landscape water. TN, total nitrogen; TOC, total organic carbon.
TOC decreased from 28.56 to 5.03 mg/L, and the removal efficiency was as high as 82.39%, with a removal rate 0.245 mg/[L·h]. The removal trends for nitrate, ammonia, and TN were basically consistent with the removal trend for TOC, and these results indicated that strain SYF26 is able to simultaneously remove organic carbon and nitrogen at high efficiencies during the whole process. An aerated batch test demonstrated the efficient removal of COD and TN by strain HA2 from synthetic wastewater (Yao et al., 2013b). Meanwhile, NO2−-N remained at a stable and low level, and the maximum nitrite accumulation was 0.055 mg/L during the whole process.
Results were in accordance with those of our previous research (Su et al., 2013), indicating that strain SYF26 is capable of simultaneous heterotrophic nitrification and aerobic denitrification, and both nitrate and nitrite could be used for denitrification, as evidenced by the successful amplification of napA and nirS genes. The bacterium Agrobacterium sp. LAD9 could use nitrite and nitrate as nitrogen sources for aerobic denitrification, and nitrate reductase nitrite reductase genes have been successfully amplified from strain LAD9 (Chen et al., 2012). These results illustrated that strain SYF26 is a suitable candidate for the simultaneous removal of nitrogen and TOC for landscape water treatment.
3D-EEM fluorescence spectroscopy assessment of landscape water components using strain SYF26
To characterize the components and distribution of dissolved organic materials in landscape water, 3D-EEM fluorescence spectroscopy was performed (Fig. 3). EEMs represent fluorescence contour maps, which comprise a series of repeated emission scans recorded at a range of excitation wavelengths (Coble, 1996).
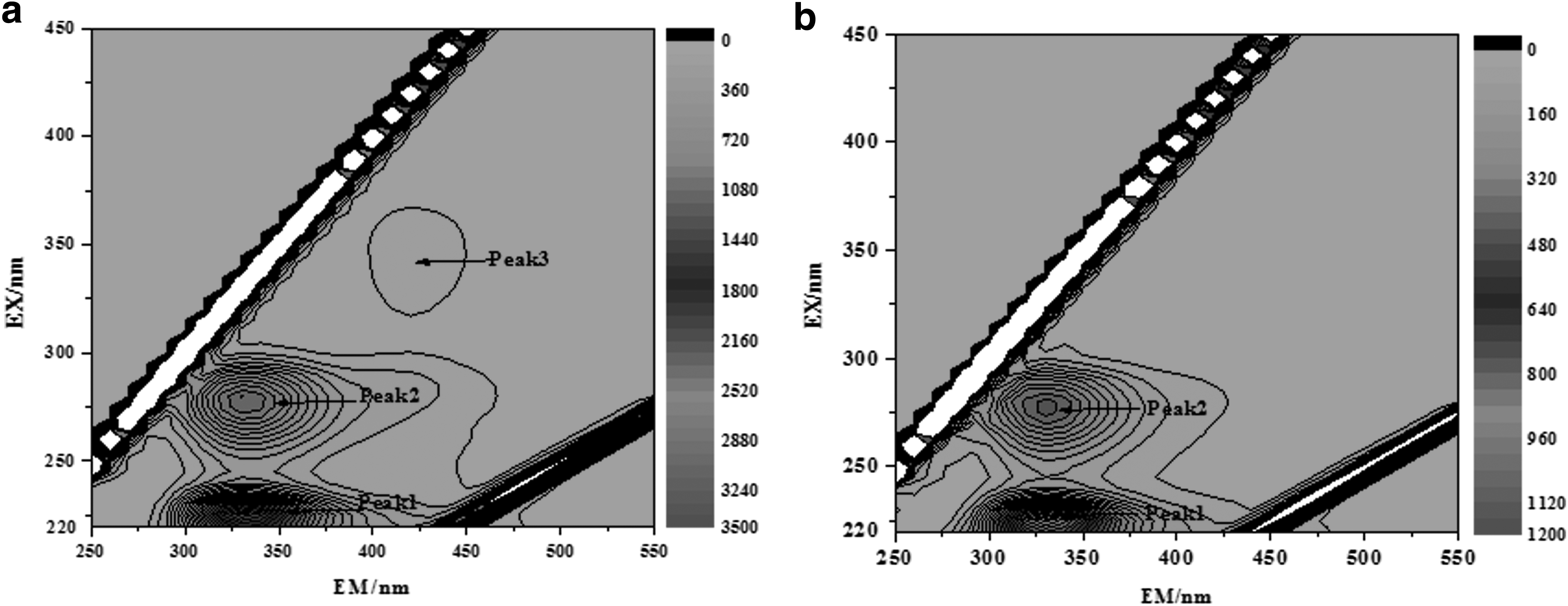
Three-dimensional excitation–emission matrix fluorescence spectra of humus in landscape water
The spectra on the first and fifth day were different. The main components of fluorescent dissolved organic matter in landscape water were UV fulvic-like, protein-like, and visible fulvic-like, and the characteristic fluorescence peaks were peak 1 (λex/λem = 225/330 nm), peak 2 (λex/λem = 280/330 nm), and peak 3 (λex/λem = 345/420 nm). The fluorescence fingerprint of dissolved organic materials showed two peaks at ∼350 nm when excited at 280 nm. These peaks are characteristic of protein-like compounds (Muller et al., 2011). The position and fluorescence intensity of the characteristic fluorescence group changed on day 5. Figure 3 shows that there was no obvious visible fulvic-like fluorescence peak, indicating that the fluorescence intensity was greatly reduced. The fluorescence intensity was significantly lower than that observed after the addition of strain SYF26 on day 1, although peak 1 (λex/λem = 225/330 nm) and peak 2 (λex/λem = 280/330 nm) were observed. Taken together, these results demonstrated significant microbial biodegradation of fluorescent visible fulvic-like organic compounds; at the same time, protein-like and UV fulvic-like organic compounds exhibited some degree of degradation.
The degradation of peak 1 and 2 and peak 3, which disappeared on day 5, was directly caused by strain SYF26. The results of this experiment demonstrated that strain SYF26 could use protein-like compounds, visible fulvic-like and UV fulvic-like as carbon sources for heterotrophic nitrification–aerobic denitrification in landscape water. Strain SZ28 has the ability to use humus as a carbon source for autotrophic and heterotrophic denitrification (Su et al., 2015). The biological degradation of microorganisms contributes substantially to the removal of fluorescent protein-like organic compounds (Hao et al., 2007).
Fourier transform near-infrared spectroscopy
FTIR spectra of landscape water with strain SYF26 were obtained on day 1 and 5. Figure 4 shows that the main absorption position of the infrared spectrogram for landscape water was different on day 1 and 5. Furthermore, compared with the infrared spectrogram of landscape water on day 1, the relative intensity of the absorption peak on day 5 was lower.
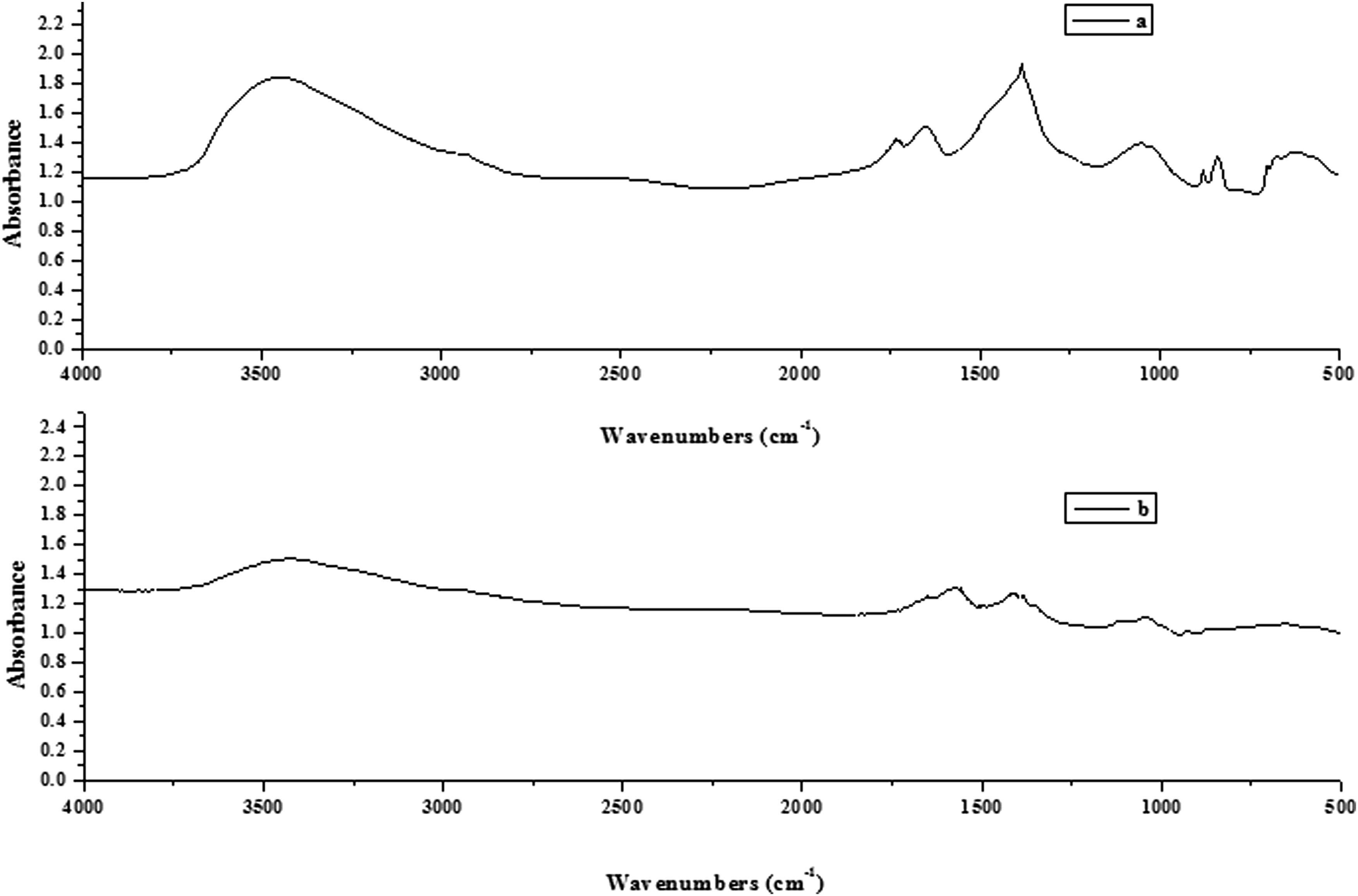
Fourier transform infrared spectroscopy spectra of landscape water by strain SYF26
The O-H stretching vibration peak at 3,500–3,300 cm−1 was observed on both day 1 and 5; however, compared with the first day, the infrared absorption density on the fifth day exhibited an obvious reduction, which was likely due to the degradation of protein-like compounds and fulvic acid in landscape water. Furthermore, the peak at 2,000–1,500 cm−1 was due to a benzene ring on day 1 and 5, and the absorption density was obviously reduced on day 5. The peak at 1,320–1,210 cm−1 was due to the C-O stretching vibration, and the absorption density exhibited a large reduction on day 5. These above results were basically consistent with the results presented in Kinetic Analysis of Heterotrophic Nitrification-Aerobic Denitrification by Strain SYF26 and Assessment of the Efficiency of Heterotrophic Nitrification-Aerobic Denitrification in Landscape Water by Strain SYF26 sections and demonstrated that strain SYF26 could use organic carbon sources such as protein-like compounds and fulvic acid for heterotrophic nitrification–aerobic denitrification in landscape water. The presence of fulvic acid not only significantly improved the TN removal efficiency but also remarkably reduced nitrite accumulation compared with conditions lacking fulvic acid (Li et al., 2016).
Conclusions
A kinetic model with first-order reactions could be used to depict the heterotrophic nitrification–aerobic denitrification process under low nitrate (2 mg/L) and ammonia (2 mg/L), and the results showed that kinetic of heterotrophic nitrification–aerobic denitrification follow the first-order reaction kinetics equation. Furthermore, the optimum condition could be achieved under pH = 7, 30°C, and C/N = 7.5. In the landscape water experiment, NH4+-N, NO3−-N, TN, and TOC removal ratios were as high as 86.98%, 98.75%, 91.78%, and 82.39%, respectively. Meanwhile, the removal rates were 0.014, 0.013, 0.034, and 0.245 mg/[L·h], respectively. These results demonstrated that the strain SYF26 was capable of both heterotrophic nitrification and aerobic denitrification in landscape water treatment. Both 3D-EEM fluorescence spectroscopy and FTIR spectroscopy assessment of landscape water illustrated that protein-like compounds and humus were decreased by strain SYF26 during the whole process.
Footnotes
Acknowledgments
This research work was partly supported by the National Natural Science Foundation of China (NSFC) (No. 51678471), the National Key Research and Development Project (No. 2016YFC0200706), and Supported by Open Project of State Key Laboratory of Urban Water Resource and Environment, Harbin Institute of Technology (No. QA201518).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.