
Abstract
Abstract
While the recovery of nutrients (nitrogen and phosphorus) from anaerobic digestion (AD) of lipid-extracted microalgae biomass has been suggested to increase microalgae biofuel sustainability, most studies use assumed biogas and nutrient recovery values that do not take into consideration how microalgae speciation and biomass composition may affect these recovery values. This study is among the first to examine how microalgae speciation and biomass composition influence the recovery of biogas and nutrients from the AD of lipid-extracted microalgae. Batch AD of lipid-extracted Chlorella vulgaris (a model freshwater microalgae) and lipid-extracted Cyclotella sp. (a model marine diatomic microalgae with a high silica composition) resulted in similar 30-day biogas yields of 0.37 ± 0.02 L-biogas/g-microalgaeVS and 0.38 ± 0.02 L-biogas/g-microalgaeVS, respectively. However, biogas production rates varied significantly between species with lipid-extracted Cyclotella producing 79% ± 1% of its biogas within the first 5 days compared to 49% ± 1% for Cyclotella. In addition, microalgae speciation and composition significantly influenced nutrient recovery with an average nitrogen recovery of 46% ± 9% for lipid-extracted Cyclotella, while a negligible nitrogen recovery (6.3% ± 7.6%) was observed for lipid-extracted Chlorella. Interestingly, for phosphate, the opposite trend occurred with an average phosphorus recovery of 78% ± 14% for lipid-extracted Chlorella and a negative phosphorus recovery from lipid-extracted Cyclotella, possibly due to its high salt content, which can result in the precipitation of phosphorus-containing salts. Thus, unlike biogas yields, nutrient recovery appears to be highly sensitive to microalgae speciation and composition.
Introduction
W
However, for large-scale production of biofuel from microalgae, the management of large quantities of residual microalgal biomass must be considered after lipid extraction. The residual microalgae biomass, which has high protein and carbohydrate content, accounts for more than 65% of the initial whole-cell microalgae biomass (Kwietniewska and Tys, 2014). The anaerobic digestion (AD) of this residual microalgae biomass has the potential to increase the environmental and economical sustainability of microalgae biofuels through the production of biogas [which consists primarily of methane (CH4) and carbon dioxide (CO2)], soluble nutrients [including ammonium (NH4+) and phosphate (PO43−)], and stabilized sludge (Mattocks, 1984; Zamalloa et al., 2011).
Combustion of produced biogas can lower the energy requirements of the production facility, while the resulting CO2 can be recycled to the cultivation ponds, thus lowering the carbon footprint of the microalgae process (Chisti, 2007; Kao et al., 2012). In addition, AD can stabilize the large quantities of residual microalgae biomass expected to be produced at these facilities, thereby being a key process in its proper disposal (Davis et al., 2012).
In addition to energy, the high demand of nitrogen and phosphate nutrients for microalgae growth also challenges the sustainability of large-scale microalgae cultivation (Sialve et al., 2009). Phosphate is a nonrenewable resource that is expected to experience significant depletion in the next 50–100 years (Cordell et al., 2009). Nitrogen can be fixed from the atmosphere utilizing the Haber-Bosch process, but the process is energy intensive and results in significant releases of greenhouse gases (Razon, 2012). Thus, the AD of lipid-extracted microalgae has gained interest as a potential process to recover nitrogen and phosphorus to lower the overall nutrient demand of large-scale microalgae cultivation processes (Sialve et al., 2009).
AD of lipid-extracted microalgae has been reported in the literature previously, but these studies focused on biogas production using primarily freshwater microalgae with relatively low inorganic content (Ehimen et al., 2009; Park and Li, 2012; Keymer et al., 2013; Alzate et al., 2014; Ramos-Suarez and Carreras, 2014). In addition, while the recovery of nitrogen and phosphate nutrients from the AD of lipid-extracted microalgae has been suggested for some time as a means to increase microalgal biofuel sustainability, the suggested recovery yields have by in large remained theoretical with a lack of laboratory studies to validate these values (Zamalloa et al., 2011; Davis et al., 2012). Thus, using batch AD reactors, this study is among the first to validate the literature predicted nutrient recovery yields from the AD of lipid-extracted microalgae. In addition, this study is the first to explore how the relatively high salt and silicon content of the diatom Cyclotella sp. (a model marine unicellular eukaryotic microalgae) affect the observed yields for nutrient recovery and biogas production during AD compared to that of Chlorella vulgaris (a model freshwater unicellular eukaryotic microalgae), which has a relatively low salt and silicon content (Martin-Jezequel et al., 2000; Jeffryes et al., 2013; Safi et al., 2014).
Materials and Methods
Preparation of microalgae substrate
C. vulgaris freshwater microalgae used in this study was obtained from the Culture Collection of Microalgae at the University of Texas at Austin (UTEX), and was cultivated in the BG-11 medium with nutrients (Allen and Stanier, 1968; Andersen, 2005). The marine diatom, Cyclotella sp., was also obtained from UTEX and was cultivated with Harrison's and Guillard's f/2 enrichment artificial seawater medium (Guillard, 1975; Harrison et al., 1980). Both microalgae species were initially cultivated in 100 mL flasks that were shook at 100 rpm and 21°C for 2 weeks. The microalgae were transferred into clear 1 L (3.8 cm diameter) polycarbonate tubular photobioreactors (PBRs), which were operated as batch reactors. The tubes were illuminated using a 16-h fluorescent light [at a light intensity of 50–100 μmol/(m2·s)] and an 8-h dark period. The PBRs were kept at 21°C and were aerated at an air flow rate of 0.2 L/min to mix the microalgal cells. CO2 was provided to the PBR to maintain a pH of 8.5, which was continuously monitored using a pH electrode. After 11–14 days of cultivation in the PBRs, both Chlorella and Cyclotella were harvested by centrifugation at 3,000 rpm for 10 min to reach a microalgal content of 5–10 wt%. The concentrated microalgae species were dried and lipids were extracted thrice with 100% hexane (Demirbas, 2008; Halim et al., 2011; Keymer et al., 2013). After extraction, the remaining microalgae residue was dried at 105°C for 1 h and stored at −20°C until use.
Preparation of batch anaerobic digesters
Batch AD reactors were constructed using 155 mL glass bottles with butyl rubber septa-sealed caps. The anaerobic inoculum was obtained from the AD reactor at the Corvallis Wastewater Treatment Facility (Corvallis, OR) and contained 20.5 ± 0.2 g/L total solids (TS) and 12.1 ± 0.5 g/L volatile solids (VS). Each batch AD reactor was set up with a consistent anaerobic inoculum concentration of 12.1 g-VS/L, which was adjusted by deionized water to reach 100 mL capacity. Dried lipid-extracted microalgal biomass (1.24–0.21 g-VS/100 mL reactor) was added to obtain an I/S VS ratio of 0.97–5.8, which corresponds to a total VS range of 24.5–14.1 g-VS/L. After the microalgal substrate and the anaerobic inoculum were added, the AD reactors were purged with nitrogen gas for 5 min to remove oxygen from the headspace. The AD reactors were then continuously shaken at 150 rpm and at 30°C for 30 days. All conditions were set up in quadruplicate. Two of the AD reactors were used for biogas measurements, while the other two AD reactors were used for liquid and solid sampling.
AD of pure anaerobic inoculum without microalgae loading was selected as the blank case since organic compounds in the anaerobic inoculum contained carbon that can be digested to produce biogas. Thus, the biogas and methane yields from lipid-extracted microalgae were calculated as the biogas and methane production from the AD of lipid-extracted microalgae minus those from the AD of pure inoculum without the microalgae. The statistical significance of biogas production and nutrient recovery was determined using the one-way analysis of variance (ANOVA) test and the Student's t-test. A p-value <0.05 was defined as being statistically significant.
Analytical methods
TS and VS of the microalgae and anaerobic inoculum were quantified separately and combined before and after the AD process by utilizing Standard Method 2540 (APHA, 2012). To avoid potential biomass measurement error due to the high salt content of the Cyclotella growth medium, Cyclotella cells were washed thrice with a 0.5 M ammonium biocarbonae solution to remove sea salts, as previously described (Zhu and Lee, 1997). After washing, the cells were dried and ammonia and carbonate were removed through volatilization. It should be noted that this was only done for aliquots used for TS/VS measurements. Cyclotella cells that were loaded into the AD reactors were not washed to keep consistent with what would be expected at full-scale facilities.
Total carbon, hydrogen, and nitrogen measurements of the microalgae and AD inoculum were performed using a Flash 2000 organic elementary analyzer (Thermo Fisher Scientific, Inc.) (Park and Li, 2012; Tran et al., 2014). Elemental analyzer combusts organic carbon, hydrogen, and nitrogen in the sample at ∼1,800°C and conveys to the thermal conductivity detector (TCD) to generate the signal for organic element contents. Total phosphorus content was quantified colorimetrically using the phosphor-vanadomolybdate method (Barton, 1948; Malhotra and Sacher, 1969).
Alkalinity and pH were measured throughout the AD process. Alkalinity was quantified using the acid titration method based on Standard Method 2320B (APHA et al., 2012). The pH was measured using a VWR sympHony benchtop pH/ion meter and probe (VWR Scientific). Ammonia was determined colorimetrically using the automated phenate method as described in Standard Method 4500-NH3 G (APHA et al., 2012). Soluble chloride, nitrate, nitrite and phosphate were measured using an ion chromatograph (IC) equipped with a Dionex IonPac AS14 column (Thermo Fisher Scientific, Inc.) with suppressed conductivity detection. The eluent carrier fluid was 3.5 mM Na2CO3/1.0 mM NaHCO3 with a flow rate of 1.5 mL/min. The AD effluent samples were centrifuged at 9,000 rpm for 15 min, and the supernatant was filtered through a 0.45 μm nylon syringe filter to remove solids from the sample before being placed through the IC unit and ammonia tests.
Volumes of biogas produced by the batch AD reactors were measured on a daily basis with a 20 mL Micro-Mate glass syringe. The needle was placed into the batch AD reactors through the butyl rubber septa top. Biogas that accumulated in the batch AD reactors flowed into the glass syringe, and the displaced volume was recorded. The methane content of the captured biogas was quantified using an Agilent 6890 gas chromatograph (GC) equipped with TCD and GS-Q column (Agilent Technologies). The temperature of the detector and oven of GC were maintained at 150°C. Helium was used as the carrier gas with a flow rate of 4.2 mL/min.
Results and Discussion
Microalgal biomass and anaerobic digester inoculum characterization
Both Chlorella and Cyclotella were cultivated in batch reactors before lipid extraction. Chlorella had an average productivity of 0.13 ± 0.04 g-microalgae/L-day (Supplementary Table S1) and reached a density of 1.1 ± 0.1 g-microalgae/L after 9–10 days of cultivation (Supplementary Fig. S1A). After 11–14 days of cultivation, Chlorella had completely consumed the initial 18 mM/NO3− and was harvested by centrifugation. Cyclotella had a similar growth rate of 0.13 ± 0.02 g-microalgae/L-day and consumed the initial 5.3 mM/NO3− within 12–14 days, at which time it was also harvested by centrifugation (Supplementary Fig. S1B).
According to the Redfield ratio and microalgal composition, microalgae contain an average dry weight content of 52% carbon, 7.7% hydrogen, 7.6% nitrogen, and 1.3% phosphorus (Richmond, 2004; Davis et al., 2012). The whole-cell Chlorella used in this study followed these theoretical ratios; however, the whole-cell Cyclotella contained less nitrogen and phosphorous than the Redfield ratio (Supplementary Table S2). It should be noted that the chemical composition of microalgae is not an intrinsic constant since the constituents of some microalgae can be modified by varying culture conditions, such as nitrogen or phosphorus concentration in the medium (Becker, 1994). Thus, the low nitrogen and phosphorus content of Cyclotella may be due to the low nutrient concentrations and differing nutrient ratio in its cultivation medium (Shafik et al., 1997).
The Cyclotella used in this study comprised 25.8% extractable lipids (Supplementary Fig. S2), which compare favorably to 19–39% lipid content reported previously (Ozkan and Rorrer, 2017). However, the cultivated C. vulgaris in this study had only 2.1% extractable lipids, which is much less than 14–25% lipid content reported in previous studies (Becker, 1994; Griffiths and Harrison, 2009). The low extractable lipid content of Chlorella may be due to initial wastewater treatment cultivation conditions from which this particular strain was grown.
Whole-cell and lipid-extracted Chlorella had a high VS/TS content of 95% ± 0.2% and 95% ± 0.1%, respectively (Supplementary Table S2). This is similar to previous studies that reported 85–90% and 92% VS/TS content of whole-cell and lipid-extracted Chlorella, respectively (Ras et al., 2011; Zhao et al., 2014). Compared to Chlorella, both the whole-cell Cyclotella and the lipid-extracted Cyclotella had a lower VS/TS content (54% ± 4.3% and 38% ± 1.2%, respectively) due to the high content of salt and other inorganic compounds (e.g., silica) found in the biomass samples (Rorrer et al., 2016), which adds additional TS to each cell compared to the freshwater Chlorella. The lower VS/TS content in marine microalgae has been reported in previous studies with the lipid-extracted marine diatoms Nanofrustulum sp., which had a VS/TS content of 49% (Zhao et al., 2014).
The low cost and boiling point of hexane make it suitable for solvent stripping and recovery, and hexane extraction has been suggested as an optimal solvent for lipid extraction at large-scale microalgae facilities (Stephenson et al., 2010; Frank et al., 2011; Davis et al., 2012). In addition, hexane extraction has similar lipid removal capabilities in the chloroform/methanol (2:1 v/v) extraction process (Lee et al., 1998) (Supplementary Fig. S2) without the added risk of introducing chloroform into the AD reactors. Previous studies have shown that sub-ppm chloroform levels (∼0.5 ppm) can inhibit methanogenesis in AD reactors (Weathers and Parkin, 2000; Hu and Chen, 2007). Thus, due to its full-scale relevancy, comparable extraction capability, and lower toxicity, hexane lipid extraction was chosen over chloroform/methanol lipid extraction for this study.
AD inoculum had a VS/TS content of 59% ± 0.7% and comprised 52% ± 0.6% carbon, 7.6% ± 0.04% hydrogen, 6.9% ± 0.07% nitrogen, and 6.6% ± 0.1% phosphorus (Supplementary Table S2). The carbon, hydrogen, and nitrogen content were similar to that of the whole-cell and lipid-extracted microalgae. However, the phosphorus content of the inoculum was more than five times greater than that in both whole-cell and lipid-extracted microalgae (6.6% vs. 0.9–1.3%).
Effect of lipid extraction on biogas and methane yields
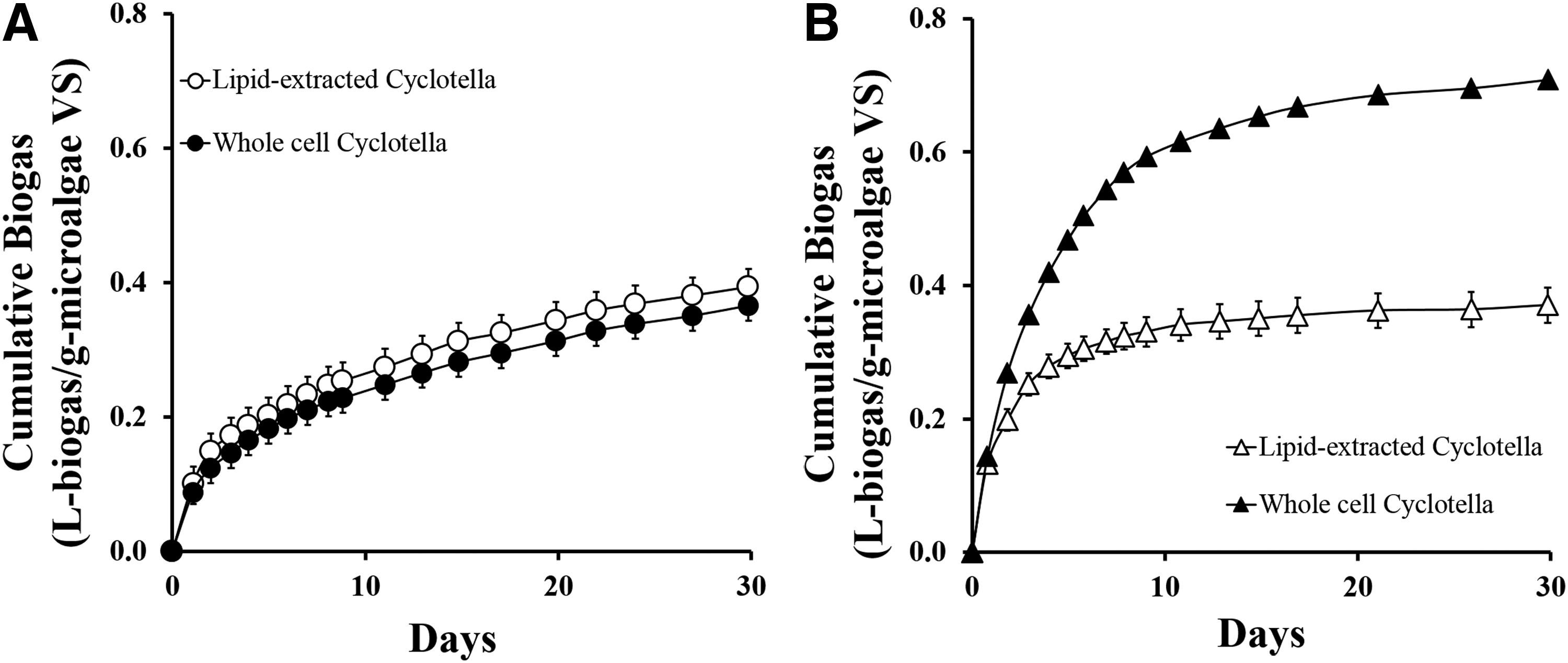
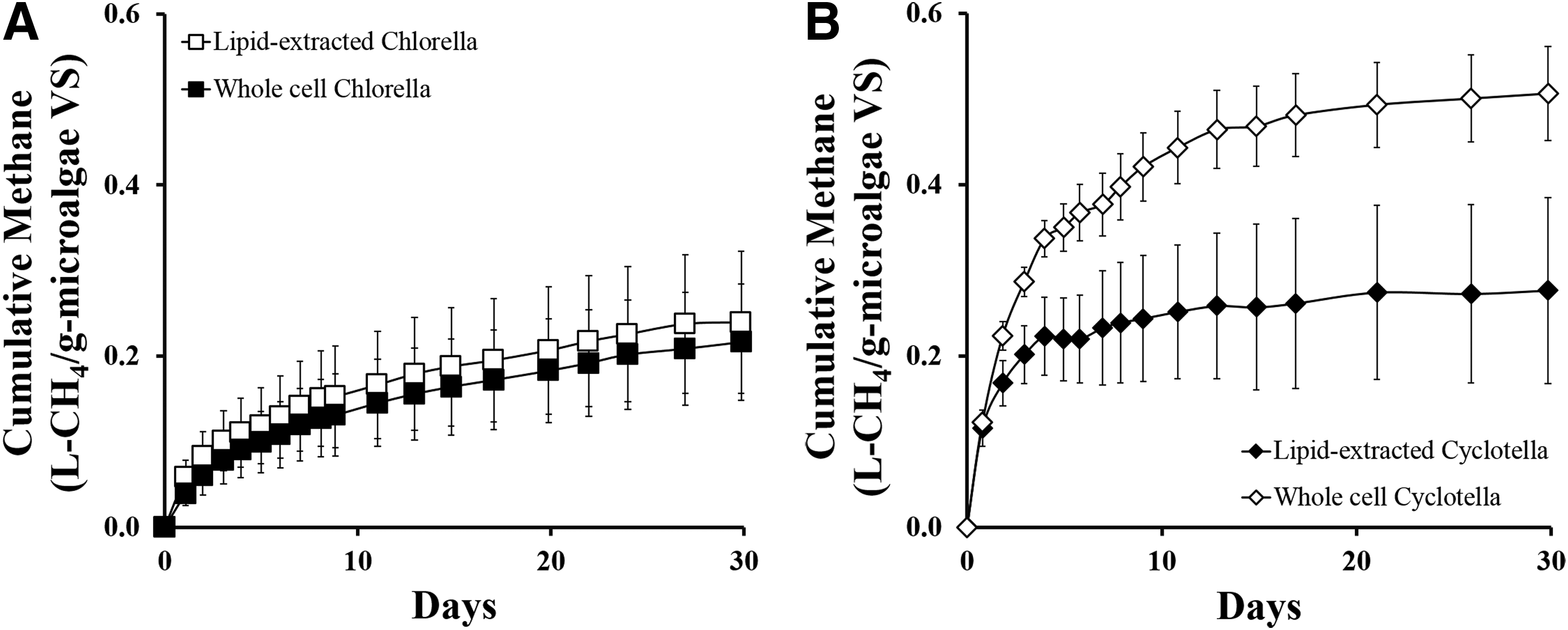
AD reactors containing a 3.9 I/S VS ratio of whole-cell or lipid-extracted Chlorella, or a 5.8 I/S VS ratio of Cyclotella were run to determine the effect of lipid extraction on biogas and methane yields. With a low amount of extractable lipids in Chlorella (2.1% ± 0.1%), the difference in the cumulative biogas volume produced by whole-cell and lipid-extracted Chlorella was almost negligible at 0.37 ± 0.02 L-biogas/g-microalgaeVS and 0.39 ± 0.03 L-biogas/g-microalgaeVS, respectively (Fig. 1). The methane content in the biogas produced from both whole-cell Chlorella and lipid-extracted Chlorella was also similar at 66% ± 6.3% and 64% ± 9%, respectively (Fig. 2A). This is in contrast with Cyclotella in which significant differences (p < 0.05) in the cumulative biogas volumes produced by whole-cell and lipid-extracted Cyclotella were observed at 0.71 ± 0.03 L-biogas/g-microalgaeVS and 0.37 ± 0.01 L-biogas/g-microalgaeVS, respectively (Fig. 1B). However, the methane content in the biogas produced by both whole-cell and lipid-extracted Cyclotella remained similar at 72% ± 10% and 74% ± 6%, respectively (Fig. 2B).
Cumulative biogas production from the AD of whole-cell and lipid-extracted Chlorella
Cumulative methane production from the AD of whole-cell and lipid-extracted Chlorella
The decrease in biogas production by lipid-extracted Cyclotella was surprising considering that the same amount of VS (2.1 g-microalgae VS/L) was added for both whole-cell and lipid-extracted Cyclotella AD reactors. However, due to the high lipid extraction efficiency for Cyclotella (25.8%), the lipid-extracted Cyclotella only had a carbon content of 44%, while the whole-cell Cyclotella had a greater carbon content at 58%. This 24% reduction in available carbon to produce biogas and correlated with the 31% decrease in biogas production observed. This is in agreement with previous estimations that have indicated that lipids, with high carbon contents (>70%), have a higher methane yield potential than the carbohydrates and proteins that would remain after lipid extraction (Sialve et al., 2009; Kwietniewska and Tys, 2014). However, while microalgal lipids provide a much greater source of carbon for biogas production, this work has shown that the residual lipid-extracted microalgal biomass can still produce high-quality biogas with high methane content (Table 1).
Biogas and methane yield per g-algaeVS were calculated from the AD of lipid-extracted Chlorella or lipid-extracted Cyclotella minus the AD of the pure inoculum without microalgae substrate.
The unit of biogas and methane yield for pure inoculum is L/g-inoculumVS (VS in inoculum).
Effect of I/S VS ratio on biogas and methane yields
AD experiments with I/S VS ratios ranging from 1.9 to 5.8 for lipid-extracted Chlorella and from 0.97 to 5.8 for lipid-extracted Cyclotella were undertaken to determine the effect of microalgal speciation and loadings on AD performance. The cumulative biogas and methane production from the AD of both lipid-extracted Chlorella and Cyclotella increased with increases in substrate loading (Table 1). This resulted in statistically similar (p > 0.05), 30-day biogas yields (0.37 ± 0.02 and 0.38 ± 0.02 L-biogas/g-micoalgaeVS, respectively), methane yields (0.21 ± 0.01 and 0.25 ± 0.03 L-CH4/g-microalgae VS, respectively), and methane content (63.25% ± 6.25% CH4 and 68.75% ± 6.75% CH4, respectively).
These 30-day methane yields are very similar to those previously reported for lipid-extracted Chlorella (0.21–0.23 L-CH4/g-microalgaeVS) and the lipid-extracted marine diatom Nanofrustulum sp. (0.30 L-CH4/g-microalgaeVS) (Ehimen et al., 2009; Zhao et al., 2014). In addition, the methane content falls within the 59–79% methane content range reported from the AD of five species of lipid-extracted microalgae (Zhao et al., 2014). Thus, at a 30-day retention time, there does not appear to be significant AD performance differences between lipid-extracted Chlorella and lipid-extracted Cyclotella, even though the Cyclotella introduced a significant increase in dissolved chloride concentration at 1,440–1,540 mg/L (Supplementary Table S3) and has a high silicon content in its biomass (Rorrer et al., 2016).
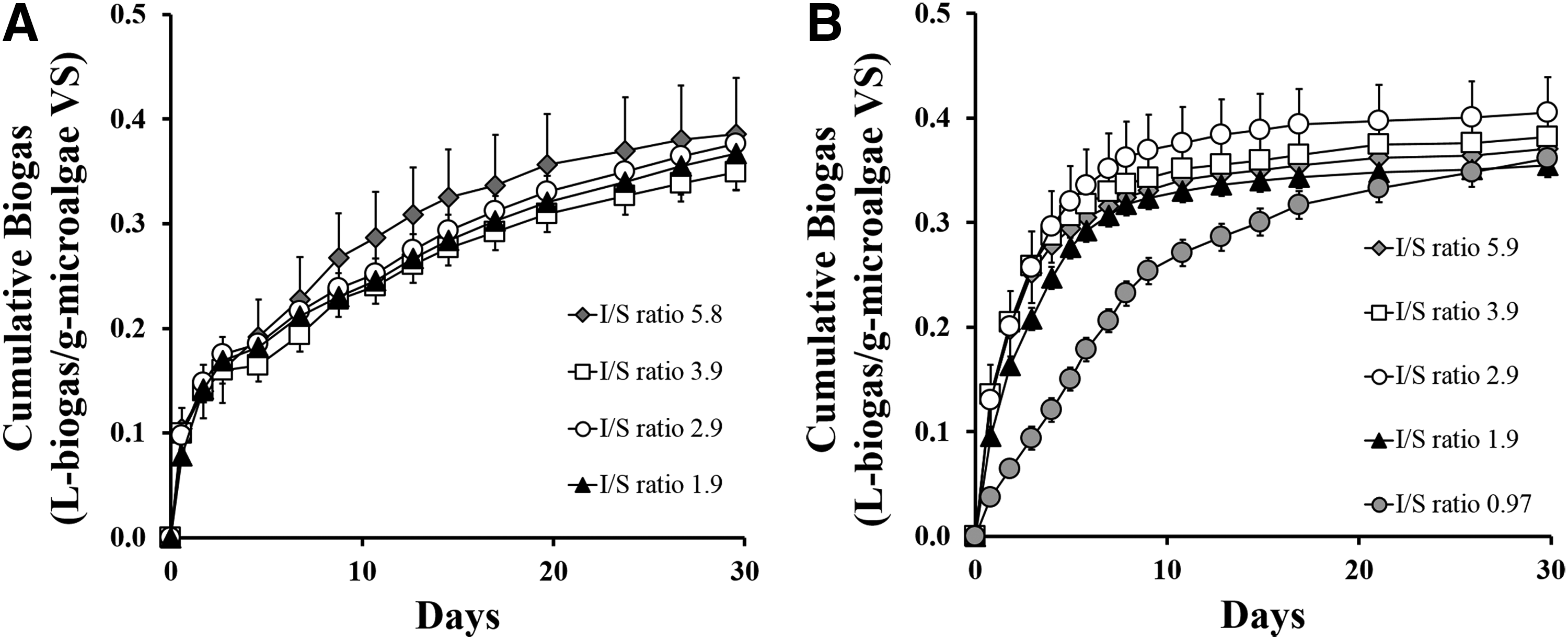
However, when looking at endpoints shorter than 30 days, differences between the two microalgae species emerge. Although the biogas production rates were the fastest during the first 5 days for both lipid-extracted Chlorella and Cyclotella (Fig. 3), a significantly higher (p < 0.05) percentage of the total biogas was produced for lipid-extracted Cyclotella (79% ± 1%) than for lipid-extracted Chlorella (49% ± 1%). It is currently unclear as to why lipid-extracted Cyclotella is digested faster than lipid-extracted Chlorella. However, the faster biogas production rates of lipid-extracted Cyclotella suggest that the AD reactors could be operated under a shorter sludge retention time compared to AD reactors digesting lipid-extracted Chlorella, which can introduce substantial cost savings to full-scale microalgae facilities.
Microalgae VS normalized cumulative biogas production from the AD of lipid-extracted Chlorella
One additional difference observed is the effect of I/S ratio on the 5-day biogas yields (Fig. 3). In the case of lipid-extracted Chlorella, the biogas yield was constant over the entire range of I/S ratios tested. However, in the case of lipid-extracted Cyclotella, a slight decrease in biogas yield was observed between I/S ratio 5.8 (0.30 ± 0.01 L-biogas/g-microalgaeVS) and I/S ratio 1.9 (0.28 ± 0.01 L-biogas/g-microalgaeVS). The decrease in biogas yield with decreasing I/S ratios became more obvious when the I/S ratio dropped to 0.97, which resulted in a 5-day biogas yield of 0.15 ± 0.01 L-biogas/g-microalgaeVS. These results are consistent with previous studies, which suggested that methane yields will be inhibited at I/S VS ratio of less than about 0.5–2, due primarily to not having sufficient microorganisms, enzymes and growth factors for organic compound digestion (Hashimoto, 1989; Chynoweth et al., 1993; Zhao et al., 2014).
Effect of I/S VS ratio on nitrogen and phosphorus recovery
AD is a potential method to recover nitrogen and phosphorus from nutrient-rich lipid-extracted microalgae (Zamalloa et al., 2011; Rosch et al., 2012). Nitrogen, primarily in the form of ammonia, is generated in AD reactors from the degradation of proteins, urea, or other nitrogen-rich compounds (McCarty, 1964). Likewise, phosphorus, primarily in the form of phosphate, is generated in AD reactors from the degradation of polyphosphates from microbial cell membranes and nucleic acids (Stabnikov et al., 2004).
AD of lipid-extracted Chlorella and Cyclotella at I/S VS ratios of 5.8–1.9 resulted in an additional total nitrogen loading of 139 ± 6–418 ± 17 mg N/L and 128 ± 8–384 ± 23 mg N/L, respectively, to the AD reactors (Table 2). The loading of lipid-extracted Cyclotella at an I/S VS ratio of 0.97 resulted in an additional 768 ± 46 mg N/L being added to the AD reactors. In both cases, the increase in lipid-extracted microalgae loading resulted in an increased N in the effluent, primarily in the form of ammonia. However, while the lipid-extracted Chlorella and Cyclotella were introduced to the AD reactors at similar loadings, the recovery of N from these two microalgae species was significantly different. Nitrogen recovery percentages from lipid-extracted Chlorella were quite low and ranged from 0% to 12% (Table 2). The ammonium yield from lipid-extracted Chlorella was 5.3 ± 5.8 mg/g-microalgaeVS. Conversely, the nitrogen recovery percentages from lipid-extracted Cyclotella were significantly higher (p < 0.05) and ranged from 35% to 60% (Table 2). The ammonium yield from lipid-extracted Cyclotella was also significantly higher at 28.4 ± 4.2 mg N/g-microalgaeVS. In both cases, the nitrogen recovery percentages and ammonium yields were relatively independent from the initial microalgae loading.
AD of lipid-extracted Chlorella and Cyclotella at I/S VS ratios of 5.8–1.9 resulted in an additional total phosphorus loading of 10 ± 2 to 30 ± 6 mg P/L and 20 ± 2 to 59 ± 5 mg P/L, respectively, to the AD reactors (Table 3). The loading of lipid-extracted Cyclotella at an I/S VS ratio of 0.97 resulted in an additional 120 ± 10 mg P/L being added to the AD reactors. The recovery of P from lipid-extracted Chlorella followed a similar pattern to that of the recovery of N in which P concentrations increased over time and more P was recovered from the highest microalgae loadings than the lowest microalgae loadings (Table 3 and Supplementary Fig. S3A). The average phosphorus yield was 3.8 ± 0.7 mg P/g-microalgaeVS.
Unlike Chlorella, the addition of lipid-extracted Cyclotella to the AD reactors significantly decreased the amount of phosphate recovered compared to that of pure inoculum (Table 3 and Supplementary Fig. S3B). The reduction of phosphate concentration for lipid-extracted Cyclotella was due to the presence of high concentrations of NH4+, Ca2+, and Mg2+ in the AD reactor. All functional AD reactors will contain high concentrations of NH4+, while the high Ca2+ and Mg2+ content were due to the salt water media transfer that occurred during the preparation of lipid-extracted Cyclotella for AD (Guillard, 1975; Harrison et al., 1980). The high NH4+, Ca2+, and Mg2+ concentrations in the presence of phosphate can lead to the formation of many phosphate precipitates, including calcium phosphate, magnesium phosphate, and struvite (Wild et al., 1997; van Langerak et al., 1998; Munch and Barr, 2001). The AD of lipid-extracted Chlorella still had high NH4+ concentrations, but the Ca2+ and Mg2+ concentrations were relatively low due to the freshwater growth medium used to prepare the cells for AD, which led to unfavorable conditions for phosphorus precipitations. Thus, the presence of Ca2+ and Mg2+ in the microalgal cultivation medium must be taken into consideration when working with marine microalgae or else, the soluble phosphate recovery will be significantly less.
Conclusion
Results of this work demonstrated that the AD of whole-cell Cyclotella microalgae produced more biogas per VS than hexane lipid-extracted Cyclotella microalgae, due to the higher carbon content of the hexane-extracted lipids compared to the residual biomass. However, significant biogas quantities were still produced from the AD of both lipid-extracted Cyclotella and Chlorella microalgae, and the 30-day biogas yields were independent of their speciation. Speciation did play a significant role in biogas production rates with lipid-extracted Cyclotella producing 79% ± 1% of its total biogas within the first 5 days of AD, compared to lipid-extracted Chlorella, which only produced 49% ± 1% of its total biogas within the first 5 days of AD. This suggests that full-scale biomass facilities could operate their AD reactors with a much shorter residence time if using Cyclotella compared to Chlorella.
In every case, increasing the microalgae loadings (from I/S ratios of 5.8 to 0.97) increased the overall biogas production and did not affect the 30-day biogas yields. However, in the case of Cyclotella, when the I/S ratio dropped to 0.97, the VS-normalized biogas production rate did decrease significantly compared to the rates observed at I/S ratios from 5.8 to 1.9. This decrease in rate does not appear to be due to inhibition, but rather overwhelming the microbial enzymes with excess substrate. This suggests that AD reactors can operate under relatively high microalgae loading rates, but high biomass populations are required to operate at maximum efficiency.
Unlike biogas production, nutrient recovery was highly dependent on microalgae speciation and recovery yields were more variable. For lipid-extracted Cylcotella, between 35% and 60% (134 ± 15 to 413 ± 9 mg/L N) of the nitrogen from the microalgae solids were mineralized to ammonium. Meanwhile, for lipid-extracted Chlorella, nitrogen recovery ranged from 0% to 12% (0 to 176 ± 57 mg/L N). Further work is needed to determine why the nitrogen recovery was so low in lipid-extracted Chlorella. Phosphorus recovery followed an opposite trend with much higher quantities of phosphorus being recovered from lipid-extracted Chlorella (11 ± 4 to 20 ± 5 mg/L-P) than lipid-extracted Cyclotella (−72 ± 1 to −93 ± 1 mg/L-P). The average phosphorus recovery from lipid-extracted Chlorella was 78% ± 14%. The negative recovery of phosphorus from lipid-extracted Cyclotella is thought to be due to its high salt content, which increased the formation of phosphate-salt precipitates. The results of this work make it clear that microalgae speciation needs to be taken into consideration when assuming nutrient recovery potentials from the AD of lipid-extracted microalgae.
Footnotes
Acknowledgment
This work was supported by the National Science Foundation (NSF)-EFRI-PSBR: The Diatom-Based Photosynthetic Biorefinery (Award No. 1240488).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.