
Abstract
The recovery of soil microbial community is important for reclamation success in coal mining subsidence land. This study aims to investigate the relationship between soil microbes and vegetation species in a reclamation area in Panji coal mining subsidence land, and to determine a suitable planting model to improve the soil physiochemical properties. In this experiment, Sesamum indicum and Glycine max, forge crops Medicago sativa and Sorghum sudanense, and Zea mays were planted, and amplicon high-throughput sequencing was applied to determine the soil microbial community structure. The results showed that significant growth-related dynamic changes in microbial community structure were mainly associated with the phylum of Proteobacteria, Actinobacteria, and Firmicutes, which accounted for 29.69%, 13.93%, and 12.51% in the total bacterial sequences, respectively. In addition, the activities of soil sucrase and urease showed a significant increasing with the decrease of pH, from 8.05 to 7.74. Correspondingly, plants had no significant effect on the fungal community composition, which comprised 52.55% of Ascomycota and 25.45% Basidiomycota, respectively. More important, soil pH was a crucial factor influencing the bacterial community structure. The study indicates that vegetation can improve the soil nutrient, enzyme activities, and microorganisms in the surface soil of the reclamation area and remit subsidence area soil alkalinization, which is of great significance for the ecosystem restoration in coal mining subsidence land.
Introduction
Coal is a major natural resource, which accounts for <70% of the energy source in China. However, mining process has its disadvantages in the form of extensive environmental degradation and ecological disturbance in the area, especially Huainan–Huaibei subsidence area in China (Frostegård et al., 1993). Compared with the farmland, the surface soil in the coal mining subsidence area often has adverse effects on plants, such as high compactness, low nutrient levels, and poor water holding capacity (Zhao et al., 2007). Thus, it is an urgent need to adopt effective strategies and rationally utilize natural resources for soil reclamation (Li et al., 2007). The interaction between plant and soil laid the foundation for an effective and sustained restoration of terrestrial ecosystems (Eviner and Hawkes, 2010). Ecosystem development can be driven by reforestation and the spontaneous establishment of certain plant species in coal mining subsidence areas. The future ecological restoration in China's subsidence areas will focus on the establishment of a standardized ecological restoration evaluation index system; the goal of ecological restoration is to meet the needs of modern ecological agriculture (Liu and Lin, 2009). Compared with the farmland cultivated for a long time, the surface soil in the subsidence area of coal mining under artificial reclamation often has adverse effects on the growth of crops (Zhao et al., 2007). Thus, to improve the poor quality of soil compaction and sterility, artificial planting is often adopted to improve soil fertility. Presently, the criteria for successful soil restoration in coal mining subsidence area have largely been narrowed to soil erosion, physicochemical status, and vegetation characteristics (Li et al., 2007; Fujii, 2014). Microbial ecological indicators, including microbial populations, microbial communities, and function diversity potential indicate soil ecosystem reclamation state (Liu and Lin, 2009).
In addition, the microbial community structure closely related to the soil physicochemical properties, accordingly reflect the environmental parameter change and can be used to evaluate soil reclamation. More importantly, soil microbes actively participate in majority of the soil biological and biochemical reactions, and are responsible for many ecological functions, such as stabilizing soil ecological balance, regulating material circulation, converting plant nutrients, and purifying environmental pollution (Brookes and Mcgrath, 1984). Correspondingly, soil microbial community structure is mostly dependent on plant type and species, both of which can affect the soil fertility (Lozano et al., 2014). For example, various chemical compounds released by roots recruit the functional microbes to set up a mutual symbiosis between plants and microbial communities. Moreover, microbes can maintain nutrient levels in plants associated soil to increase the plant productivity in extreme environment (Hause and Schaarschmidt, 2009; Fujii, 2014).
The distribution and regulation of soil microorganisms in coal subsidence area were discussed through different types of vegetation restoration. The knowledge is often limited about the composition of microbial communities associated with reclamation and its interactions affecting ecosystem processes. The study aims to provide new insights on land reclamation and ecological restoration on coal mining subsidence land by studying the effects of different types of plants on the soil environment.
Materials and Methods
General situation of the study area
This study was conducted for Panji mining area (32°46′17″ N, 116°49′58″ E) in Huainan city, Anhui province. This area has warm temperature and monsoon climate all around the year. The annual average temperature is about 15°C and the annual rainfall of about 937 mm. The soil is yellow brown. The subsidence area of coal mining is dominated by dry farming, with wheat/corn rotation as the main planting method, and oil crops, including Glycine max (GM) and Sesamum indicum (SI) were important cash crops.
In this study, a flat and newly reclaimed area was selected as the experimental site in the Panji ecological park. The whole test site is divided into 18 test plots, each test plot covers an area of 15 m2 (length × width = 5 m × 3 m), and 2 m gap was reserved between each plot. Five plants were planted without base fertilizer, oil crops SI and GM, forage crops Medicago sativa (MS) and Sorghum sudanense (SS), and food crops Zea mays (ZM). The row spacing and plant spacing of ZM and MS are 50 cm, SI and GM are 20 cm, and SS are 30 cm. In May 2017, a single plant was uniformly planted in 3 experimental plots as a repeat experiment, and three control check plots without plants (CK).
Sampling and soil physical and chemical properties' analyses
In November 2017, three mixed soil subsamples were collected from each experimental plot using the diagonal distribution method (sampling depth: 0–20 cm), mixed and preserved in an aseptic bag (∼100 g), then stored in prepared ice boxes, transferred to the laboratory. The sample is divided into two parts, one part should be air dried naturally for at least 1 week, sieved through a 0.2 mm mesh, removing the gravel, animal and plant debris on the surface of the soil, before determination of soil properties, and stored in a refrigerator at 4°C before further analyses. The other samples were stored at −20°C before microbial species testing. Soil water content (SWC) was measured by weighing method. Soil organic matter (SOM) was determined by oxidation with potassium dichromate in a heated oil bath. Soil total nitrogen (TN) by the Kjeldahl method, ammonium nitrogen (NH4+-N) by the indophenol blue colorimetric method, and nitrate nitrogen (NO3−-N) by the ultraviolet spectrophotometric method. Soil pH value and electrical conductivity were measured by using pH and conductivity meters after diluting the soils to make the soil to solution ratio of 1:2.5, respectively. Soil urease activity (SUA) was determined by indigo powder blue colorimetry (Tabatabai and Bremner, 1972), expressed as NH4+-N (mg/kg). Soil sucrase activity (SSA) was determined by colorimetric method (Jiang et al., 2000), and expressed as the content of reducing sugar (mg/g).
DNA extraction and high-throughput sequencing of 16S rDNA and internal transcribed spacer
Around 0.5 g each of fresh soil sample was weighed to extract the total genomic DNA according to the extraction method of Zhou et al. (1996). Using the cetyltrimethyl ammonium bromide (CTAB) method with minor modifications. Cell lysis was performed using sodium dodecyl sulfate 20% and 20 mg/mL proteinase K, followed by CTAB 1% incubation and extraction with phenol–chloroform (Cerqueira et al., 2011). The extracted DNA was preserved in the refrigeration cabinet at −20°C for further analysis.
The V4V5 region of the 16S rDNA gene was amplified using the bacterial 515F–907R primer set. The primer sequences were: 515F (GTGCCAGCMGCCGCGGTAA) and 907R (CCGTCAATTCMTTTRAGTTT). The internal transcribed spacer (ITS)2 region of the ITS gene was amplified using the fungal ITS3–ITS4 primer set. The primer sequences were: ITS3F (GCATCGATGAAGAACGCAGC) and ITS4R (TCCTCCGCTTATTGATATGC). PCR reactions, containing 25 μL 2 × Premix Taq (TaKaRa Biotechnology, Dalian Co., Ltd, China), 1 μL each primer (10 mM), and 3 μL DNA (20 ng/μL) template in a volume of 50 μL, were amplified by thermocycling: 5 min at 94°C for initialization; 30 cycles of 30 s denaturation at 94°C, 30 s annealing at 52°C, and 30 s extension at 72°C; followed by 10 min final elongation at 72°C. The PCR instrument was Bio-Rad S1000 (Bio-Rad Laboratory, CA).
Sequencing libraries were generated using the NEBNext® Ultra™ DNA Library Prep Kit for Illumina® (New England Biolabs, MA) following the manufacturer's recommendations and index codes were added. The library quality was assessed on the Qubit@ 2.0 Fluorometer (Thermo Fisher Scientific, MA) and Agilent Bioanalyzer 2100 system (Agilent Technologies, Waldbron, Germany). At last, the library was sequenced on an IlluminaHiseq 2500 platform and 250 bp paired-end reads were generated (Guangdong Magigene Biotechnology Co., Ltd., Guangzhou, China).
Statistical analysis
Duncan test with one-way analysis of variance was used to compare the differences of soil physicochemical properties and multiple comparisons (p < 0.05) between different treatments, and bivariate correlation analysis was also used to analyze the correlation between soil nutrients and soil enzyme activities. SPSS 19.0 was used for physical and chemical data and Pearson correlation analysis. Alpha diversity, including the Shannon, and Simpson indices were analyzed using Mothur (version 1.31.2) and R software. The uniformity of bacteria and fungi was expressed by the Pielou index. Mothur was used to calculate the similarity of different operational taxonomic units (OTUs), and sequences with similarity above 97% were clustered into OTUs. Bray–Curtis distances were calculated based on OTU matrix by using R software (version 3.4.0). Species analysis was based on OTU classification. The Curtis distance matrix was used to perform nonmetric multidimensional scaling (NMDS). Canonical correlation analysis (CCA) was performed in the R software to identify the major factors affecting the structure of bacterial and fungal communities. Cluster heat map and network diagram were performed by R software.
Results
Effects of plant growth on physiochemical properties of soil
The soil in the reclamation area of coal mining is slightly alkaline as shown in Table 1. The pH of soil having plants, such as ZM, SI, GM, MS, and SS was lowered as compared with the control. The soil pH value of five crops ranged from 7.74 to 7.76, and pH of control was 8.05. The pH of the control was measured to be higher than those cultivated with plants. Meanwhile, the SOM contents were significantly higher compared with the control group due to the planting of crops (p < 0.05), and it was 2.12–2.85 times as much as the control area. The SWC of MS and SS plants were also significantly higher than the control.
Main Chemical Properties of Soil in Different Planting Areas
ZM, SI, GM, MS, and SS, respectively, represent soil samples from planting areas of Zea mays, Sesamum indicum, Glycine max, Medicago sativa, and Sorghum sudanense, while CK represents soil samples from planting areas of five plants as the control.
Different superscript letters (a, b, c, d) in the same column highlight significant differences that occurred after different treatments (p < 0.05).
NH4+-N, ammonium nitrogen; NO3--N, nitrate nitrogen; SOM, soil organic matter; SSA, soil sucrase activity; SUA, soil urease activity; SWC, soil water content; TN, total nitrogen.
The TN, NH4+-N, and NO3−-N in the soil of SI, GM, MS, and SS planted in the reclamation area of coal mining were all higher compared with the control, the TN and NO3−-N in ZM area were higher than control. However, the TN contents in the surface soil were significantly higher than the control; it was 2.2–3.28 times as much as the control area. Moreover, both leguminous plants (GM, MS) and SS have higher NH4+-N content as compared with ZM and SI.
Nevertheless, the surface SUA of control is 26.67 mg/kg; significantly, soils planted with GM have increased 513% than control. Although the planting of ZM, SI, MS, and SS have increased the SUA, there was no significant difference. The surface SSA of control is 0.70 mg/g, the reclamation area was significantly increased (1611–2393%) by crop planting, with the order of SS > SI > MS > GM > ZM.
Table 2 shows that the correlation analysis of soil nutrients and soil enzyme activities showed that SSA was significantly negatively correlated with pH, but possessed a positive correlation with SOM contents. There was a significant positive correlation between SUA and soil TN content.
Correlation between Soil Nutrients and Enzyme Activities
Indicates that the correlation coefficient is significantly correlated at the level of p < 0.01.
Analysis of soil microbial community structure in different plant-growing areas
The dominant bacteria (the ratio to total above 1%) were Proteobacteria, Actinobacteria, Firmicutes, Bacteroidetes, Acidobacteria, Chloroflexi, Gemmatimonadetes, Cyanobacteria, Verrucomicrobia, and Planctomycetes (Table 3). Among them, the difference from the control of Proteobacteria, Actinobacteria, Verrucomicrobia, and Chloroflexi were the most consistent among the soil bacteria in different planting areas. The content of Proteobacteria in the control group accounted for 51.82% of the total bacteria and that the planting area was significantly lower as compared with control (38.02–49.42%) (p < 0.05). Moreover, the content of Verrucomicrobia in the planting area was higher (72.97–206.31%) than control (p < 0.05). Chloroflexi contents in the planting area was 50.60–75.18% higher compared with control, but there was no significant difference. In addition to SI and GM, the Actinobacteria were significantly increased in ZM, MS, and SS. It was found that Acidobacteria and Gemmatimonadetes contents in planting areas were greatly higher than control. Except for MS and SS, the content of Firmicutes in soil of plant area was lower compared with the control area, and there was no significant difference. Likewise, the Planctomycete contents were slightly higher than the control area, except the SS, which has lower contents than the control area.
Analysis of Variance Analysis of Relative Percentage Abundance of Bacteria-Dominant Gates in Different Planting Areas
ZM, SI, GM, MS, and SS, respectively, represent soil samples from planting areas of Zea mays, Sesamum indicum, Glycine max, Medicago sativa, and Sorghum sudanense, while CK represents soil samples from planting areas of five plants as the control.
Different superscript letters (a, b, c, d) in the same column highlight significant differences that occurred after different treatments (p < 0.05).
Meanwhile, our results showed that following phyla, Ascomycota, Basidiomycota, Zygomycota, Chytridomycota, Glomeromycota, and Rozellomycota, were most dominant in plant areas (Table 4). Whereas, we have also observed that Ascomycota and Chytridiomycota were lower than the control planting area. However, Basidiomycota was higher than control, but there was no significant difference between them. Except that the content of Glomeromycota in the soil of the SI planting area was 120% higher compared with the control area, the other soil samples were all lower compared with the control area. The content of Zygomycota in the planting area was lower than the control. Except that the content of Rozellomycota in the soil of SS growing area was lower compared with the control area of 55.56%, it was not detected in other soil samples.
Analysis of Variance Analysis of Relative Percentage Abundance of Fungi-Dominant Gates in Different Planting Areas
ZM, SI, GM, MS, and SS, respectively, represent soil samples from planting areas of Zea mays, Sesamum indicum, Glycine max, Medicago sativa, and Sorghum sudanense, while CK represents soil samples from planting areas of five plants as the control.
Different superscript letters (a, b) in the same column highlight significant differences that occurred after different treatments (p < 0.05).
The MiSeq high-throughput sequencing results showed the plant growing area significantly improved the diversity of soil bacteria and fungi as compared with the unplanted control soil. Heatmap was made on the richness of sequences contained in different OTU (different genus) in each sample after clustering.
The heatmap also showed that the following bacteria, such as Lactobacillus, Pseudarthrobacter, Lactococcus, Acinetobacter, Sphingomonas, Vibrio, RB41, Leuconostoc, Pseudomonas, Aeromonas, Nocardioides, and Aridibacter, were more dominant (Fig. 1). In the planting area, the contents of Lactobacillus, Lactococcus, RB41, Nocardioides, Aridibacter, Opitutus, Flavobacterium, Gemmatimonas, Dongia, Gemmatirosa, Haliangium, Caulobacter, Acidibacter, and Bryobacter were higher compared with the control group; but Acinetobacter, Vibrio, Pseudomonas, Aeromonas, Hymenobacter, Staphylococcus, Deinococcus, and Methylobacterium showed lower relative abundance.
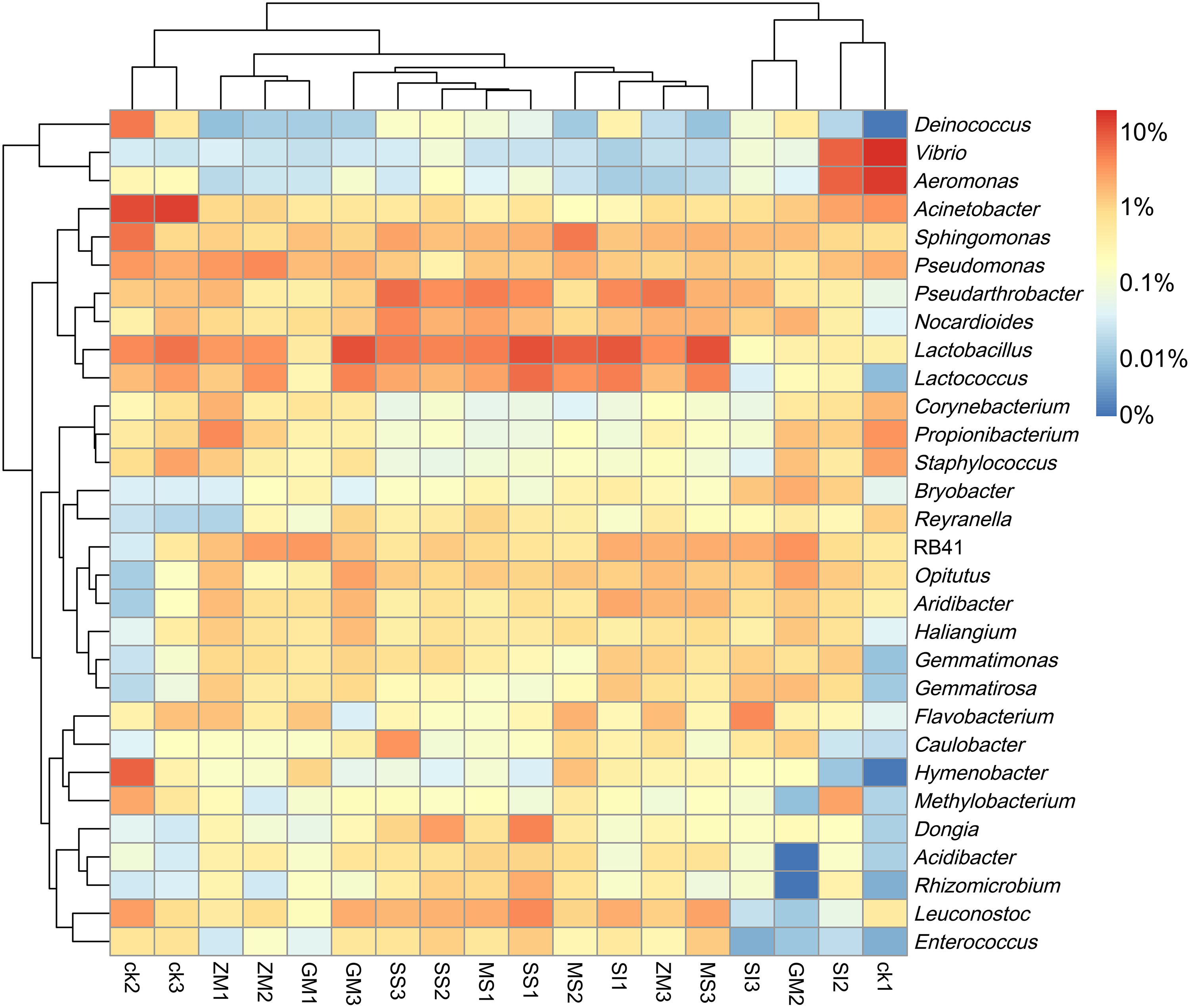
Heatmap was made on the richness of sequences contained in different OTU (different genus) in ZM, SI, GM, MS, SS, and CK groups for the top 30 bacteria genera after clustering. Red and blue indicate higher and lower expression values, respectively. CK, control check group; GM, Glycine max; MS, Medicago sativa; OTU, operational taxonomic unit; SI, Sesamum indicum; SS, Sorghum sudanense; ZM, Zea mays.
The changes in the fungal diversity and the abundance of different soil samples are shown in the heat map (Fig. 2). These Clonostachys, Fusarium, Hygrocybe, Lophiostoma, Malassezia, Myrothecium, Neurospora, Papiliotrema, Podospora, and Ramularia, and relative abundance of soil fungi in the growing area were higher than the control area. The relative abundance of Cercospora was lower than control.
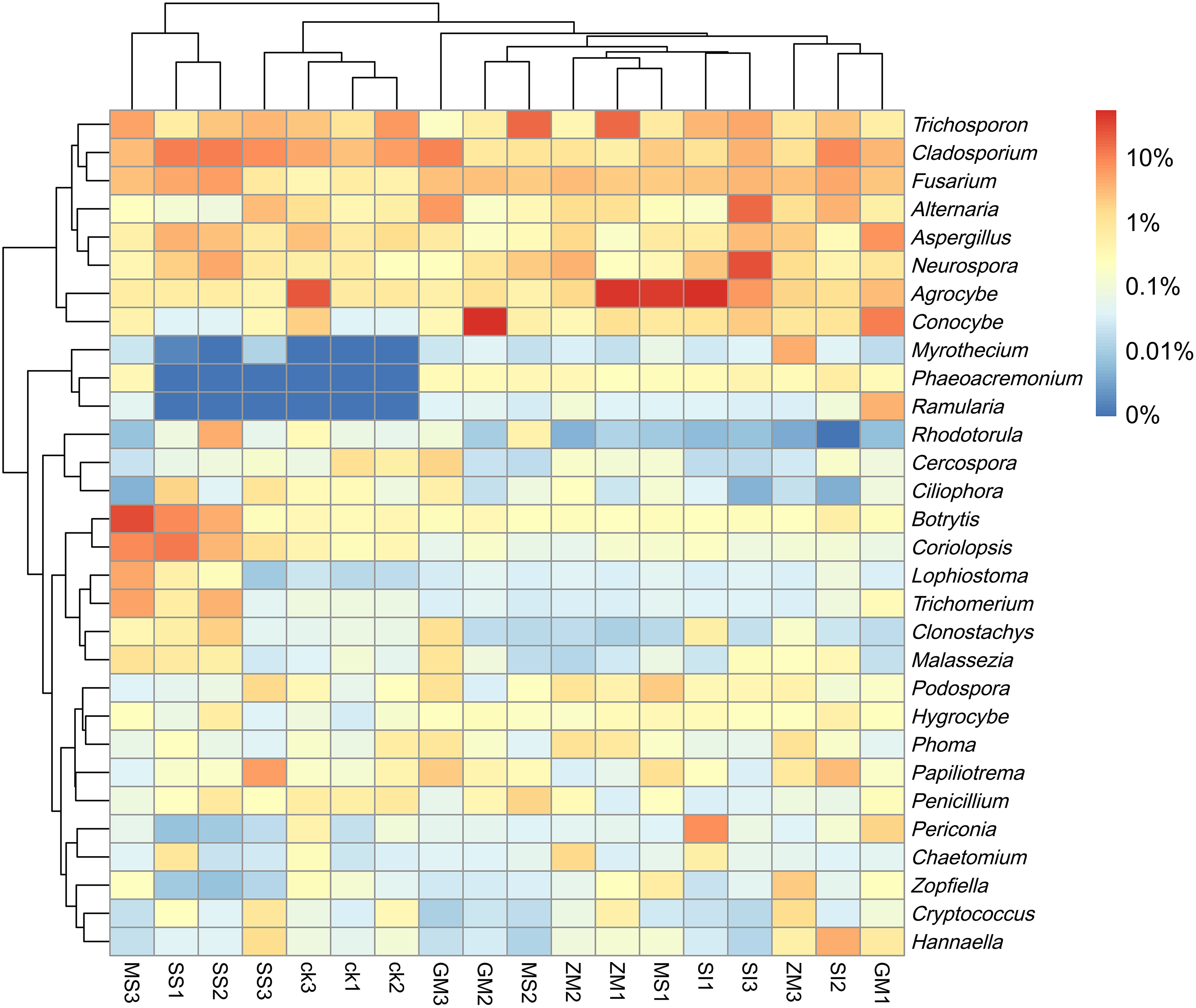
Heatmap was made on the richness of sequences contained in different OTU (different genus) in ZM, SI, GM, MS, SS, and CK groups for the top 30 fungi genera after clustering. Red and blue indicate higher and lower expression values, respectively.
Factors affecting microbial diversity and structure in different plant-growing areas
Chao1 algorithm estimates the index of OTU number in the community (Table 5); the soil bacterial 16S rDNA diversity index and soil physical and chemical properties were analyzed by canonical correspondence analysis (Fig. 3), and it was found that bacterial diversity of soil was affected by following factors such as pH, SSA, SUA, TN, and SOM (p < 0.05), and soil pH was the most profound environmental factor, followed by SSA, SUA, TN, and SOM. Among them, soil bacteria present in the planting area of SS were significantly positively correlated with SSA and SOM, whereas, soil bacteria in GM planting area showed a significantly positive correlation with TN and SUA.
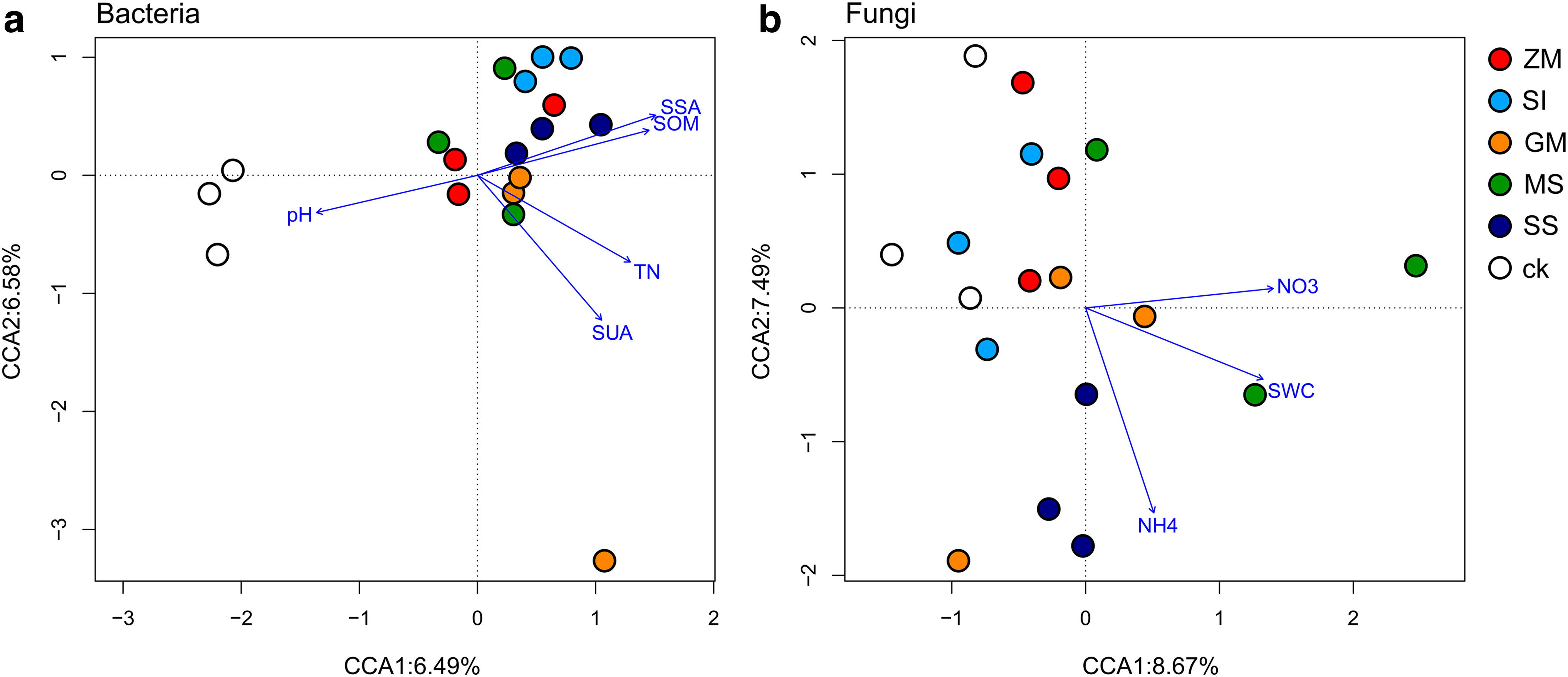
CCA was used to analyze the structure of the bacteria
Analysis of Microbial Diversity Indexes in the Soil of Different Planting Areas
ZM, SI, GM, MS, and SS, respectively, represent soil samples from planting areas of Zea mays, Sesamum indicum, Glycine max, Medicago sativa, and Sorghum sudanense, while CK represents soil samples from planting areas of five plants as the control.
Different superscript letters (a, b) in the same column highlight significant differences that occurred after different treatments (p < 0.05).
NO3−-N, NH4+-N, and SWC were the major factors, which can influence the fungal diversity of soil. Among them, NH4 had a great impact on soil fungal diversity in SS planting areas, and the correlation was positive. Soil fungal diversity in MS growing areas was positively correlated with SWC and NO3−-N.
Through CCA, our study found that soil pH was the key factor affecting soil bacterial diversity, indicating that there is correlation between soil pH and bacteria biota. Except SS and MS growing areas, different physical and chemical properties had no significant influence on the diversity of soil fungi, which may be due to the relatively intensive planting of forage crops, relatively high surface vegetation coverage, high water content, and weak light conditions, providing a unique living environment for fungi.
The NMDS figure analysis showed that the bacterial community richness of MS and SS region were the most similar and significantly higher than control (Fig. 4). The dynamic changes of soil fungal communities were also studied in different planting areas and control areas. We have found that there was little difference in soil fungal community dynamics between the SS and control soil. However, several other plants have obvious effects on fungal community dynamics.
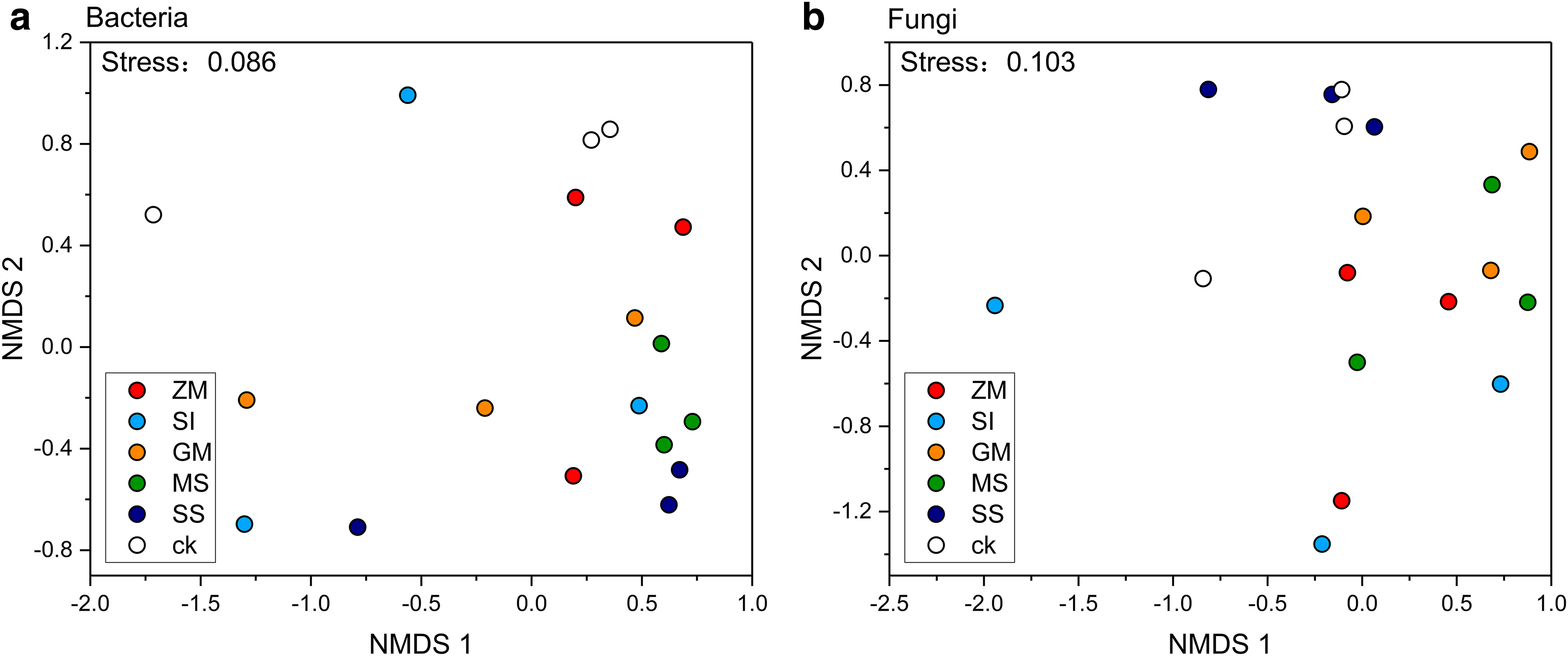
NMDS analysis was performed for the UniFrac distance of microbial community data at the six treatment groups, and analysis of similarity,
The network diagram (Fig. 5) shows that Proteobacteria have the most extensive influence on microbial composition, followed by Ascomycota and Firmicutes. In chemical properties, pH, NO3−-N, SWC, SSA, and SOM had the closest relationship with Proteobacteria, NH4+-N, and SSA had the closest influence on Ascomycota, TN had the most significant influence on Acidobacteria, and NH4+-N and water content had the most significant influence on Basidiomycota. Among them, microbial species are interwoven and closely related.
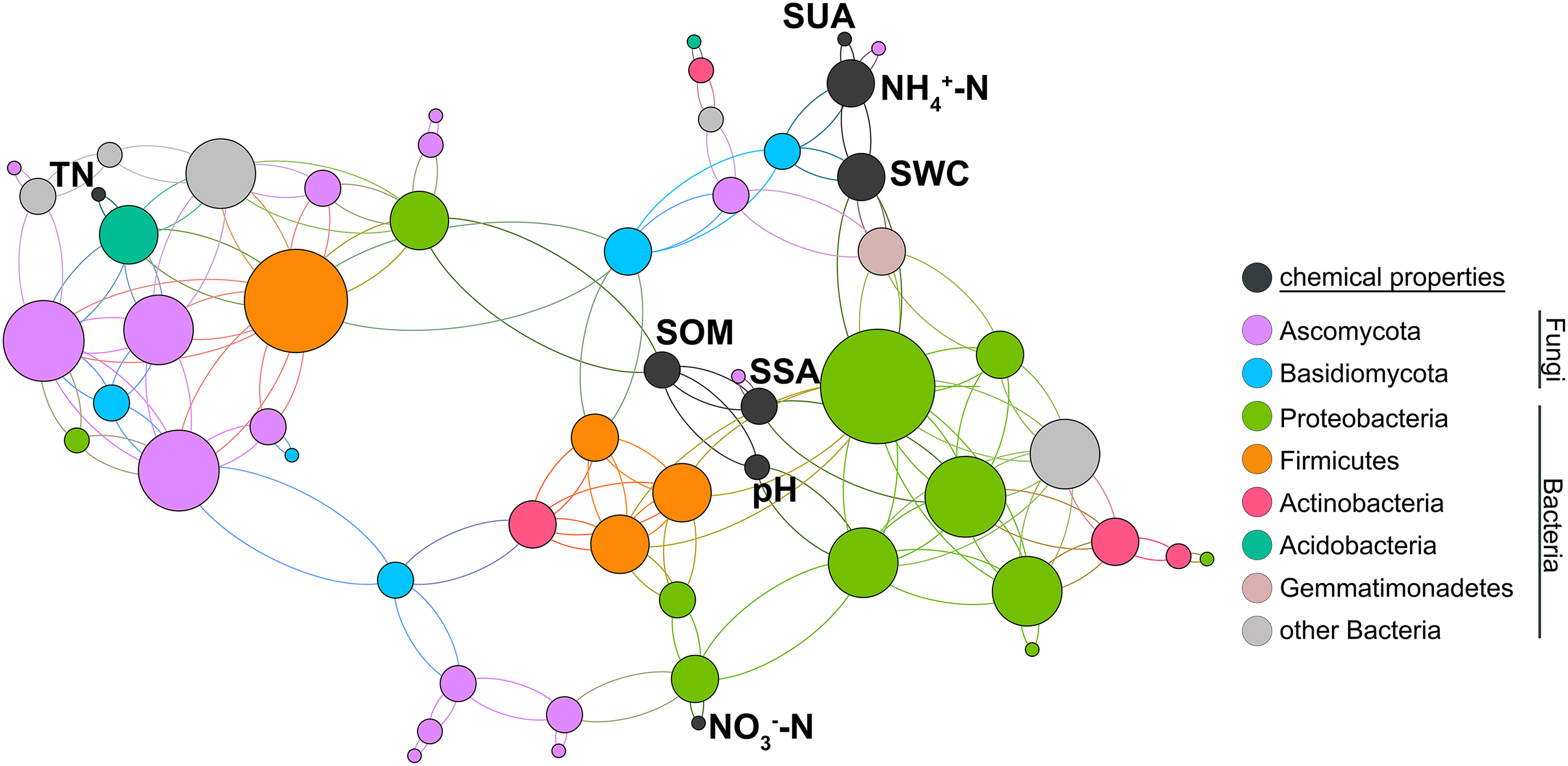
The interaction between soil-dominant bacteria and physical and chemical properties was analyzed in the network diagram. NH4+-N, ammonium nitrogen; NO3--N, nitrate nitrogen; SOM, soil organic matter; SSA, soil sucrase activity; SUA, soil urease activity; SWC, soil water content; TN, total nitrogen.
Discussion
Coal mining has caused serious damage to the soil ecological environment in the mining area, and five plants cultivated in this field can improve the soil enzyme activities and nutrient in the surface soil of the reclamation area. Soil enzymes mainly come from the microbes, animals, and plant roots, and the increase of their activity is an important indication of improvement in soil qualities such as physical and chemical properties, increased activities of microorganisms, and soil enzymes can be used as important biological indicators to measure the restoration of damaged ecosystem (Liu et al., 2014). In this study, vegetation restoration led to the increase of microbial diversity and enzyme activity, which is mainly due to the win/win cooperation between microorganisms and plants. Soil enzymes are active participants, all biochemical processes in the soil, and they are shown to be sensitive and reliable indicators of soil biological activity and soil fertility. The sucrase breaks down sucrose molecules in the soil into fructose and glucose, which is conducive to the uptake and utilization by plant and microbe. The activity of the enzyme may reflect the accumulation and decomposition of soil organic carbon (Bi and Zhang, 2014). The surface SSA of control is 0.70 mg/g, the reclamation area was significantly increased (1611–2393%) by crop planting. SSA is an important enzyme widely existing in soil, which plays an important role in increasing soluble nutrients in soil. Bacteria, which can hydrolyse urea are common in soils (Lloyd and Sheaffe, 1973). Urease can hydrolyze organic urea compounds into NH4+-N, making them available to plants. The activity of the enzyme has a close relationship with nitrogen. The surface SUA of control is 26.67 mg/kg, significantly, soils planted with GM have increased by 513% than control. Urease in soil enzyme system research is relatively in-depth, its enzymatic reaction product ammonia is one of the nitrogen sources of plant; its activity reflects the soil organic nitrogen to effective nitrogen conversion capacity and soil inorganic nitrogen supply capacity that can play a key role in the growth of plants. The results clearly showed that the cultivation of plants can greatly enhance the activity of metabolic enzymes in the soil, and are very effective at improving soil fertility.
In this study, there was a certain difference in soil microbial diversity index (observed species) under different plant cultivation in half a year of reclamation, which was reflected in the following order: (Bacteria) SS > SI > ZM > MS > GM > CK; (Fungi) ZM > GM > MS > SI > CK > SS (Table 5). In addition, a total of 10 dominant bacterial phyla (Proteobacteria, Actinobacteria, Firmicutes, Bacteroidetes, Acidobacteria, Chloroflexi, Gemmatimonadetes, Cyanobacteria, Verrucomicrobia, and Planctomycetes) were detected in the fields. Among the detected bacteria, Proteobacteria were more dominated among all the samples, followed by Actinobacteria, Firmicutes, and Bacteroidetes. The results is the same as the bacterial community diversity in the reclamation soil in Xuzhou coal mining subsidence area (Li et al., 2014). Gemmatimonadetes and Nitrosospira were most widely distributed in the Xuzhou reclamation area; Acidobacteria, Actinobacteria, and Bacteroidetes were most widely distributed in common agricultural soil (nonsubsidence soil), and Proteobacteria was most widely distributed in the subsidence area (Cohen et al., 2017). Plants can effectively improve the soil in the reclamation area, making it tend to the common agricultural soil, and the soil microbial composition gradually tends to the common agricultural soil, which effectively promotes the metabolic activity in the soil enzymes and material circulation. In sum, microbial composition, like that of larger organisms, can affect ecosystem functioning (Reed and Martiny, 2007). After several crops were planted in the reclamation area, the abundance of bacteria was increased, including genus Lactobacillus, Lactococcus, RB41, Nocardioides, Aridibacter, Opitutus, Flavobacterium, Gemmatimonas, Dongia, Gemmatirosa, Haliangium, Caulobacter, Acidibacter, and Bryobacter. Lactobacillus can decompose organic matter (such as sugar), produce hydrogen peroxide, and organic acid that can reduce environmental pH value, and improve plant disease resistance. Lactococcus is one of the most abundant phylotypes in each soil sample, based on metagenomic data (Ren et al., 2018). Lactococcus is a typical plant-related functional bacterium that modifies the activity of plant cell walls. Lactococcus has a constitutively expressed transporter protein, which has significant quantitative and physiological effects on NO3-N budding transport under salinity conditions (Taochy et al., 2015). Nocardioides is a kind of Actinomycetes, which can degrade cellulose. Some studies have shown that Nocardioides, Lactobacillus, and Bacillus as a biological agent in proportion, can promote the indigenous microorganisms in the original soil, thereby, improve the biological population of the soil, gradually reduce the pH of the soil, mitigate the toxicity of the heavy metals, thus restore and improve the soil vitality. Nitrification can be performed by Aridibacter, members of the class Blastocatellia, which are frequently found in soils with a neutral and (slightly) basic pH where they constitute an important fraction of the microbial community (Huber et al., 2017). Gemmatimonas is a Gram-negative, aerobic, polyphosphate-accumulating microorganism (Zhang et al., 2003). Similarly, Reyranella can reduce nitrate to nitrite and secretes cellulose (Pagnier et al., 2012; Kim et al., 2013; Lee et al., 2017). In addition, Opitutus bacteria genera belonging to Reyranella, having fermentation substrates, including monosaccharides, disaccharides, and polysaccharides, the main end products of fermentation such as propionate and acetate, and a small amount of succinate and lactic acid ethanol, etc., which can decrease the soil pH, generate propionic acid salt, and lactic acid salt that can also further degrade methane and carbon dioxide, promoted the carbon cycle in the soil. At the same time, it can also take nitrate as electron acceptor and react with glucose to produce nitrite, which promotes the nitrogen cycle process of soil (Chin et al., 2001; Chin and Janssen, 2002; Park et al., 2017). Caulobacter can promote the transport of quaternary ammonium compounds and cations, such as Ca2+and Mg2+ in soil, actively taking up most of the nutrients from dilute environments (Benz et al., 2015). Bryobacter belonging to the phylum Acidobacteria, is an aciduric bacteria that can grow at pH 4.5–7.2, cells show alkaline and acid phosphatase enzyme activities and are able to utilize the following nitrogen sources: NO3−-N, and NH4+-N (Kulichevskaya et al., 2010). Rhizomicrobium can degrade cellulose, ferment a variety of sugars, produce lactic acid, acetic acid, and fumarate, and can use iron, nitrate, and oxygen as electron receptors to reduce nitrate to nitrite (Nie et al., 2017).
Our study results also revealed that in the five different plant growing areas and the soil of the control group, the dominant phyla of fungi were Ascomycota, Basidiomycota, Zygomycota, Chytridomycota, Glomeromycota, and Rozellomycota. Compared with the blank control area, the diversity index of fungi in soil under different crop vegetations increased. Except the SS growing area, the abundance of Phaeoacremonium, Myrothecium, and Ramularia in other soil samples have increased significantly, especially Phaeoacremonium. However, the fungi of these three genera were hardly found in the soil of SS and control group. Phaeoacremonium is a vascular plant pathogen that causes woody plants to wilt and die (Lizel et al., 2005). Probably because SS is an annual grass, and its stems are thin, there is almost no Phaeoacremonium in the soil under vegetation. Myrothecium is a kind of hypocotyl fungi. It is a highly efficient cellulose-decomposing fungus, a common plant pathogen, with antifungal activity. Its dead cells and secretions can be used as a biological insecticide to specifically kill nematodes that attack plants through roots (Turhan and Grossmann, 1994; Baldrian et al., 2011). The Ramularia belong to the Ascomycota and also a plant pathogen. Ramularia had almost no presence in the control and SS growing areas, followed by the MS growing areas, but had high concentrations in the other plant growing areas. One study has also observed that the increase in plant secondary metabolism and lignification does not protect plants, but rather enhances the infectivity of fungi (Schützendübel et al., 2008).
The correlation analysis of soil nutrients and soil enzyme activities showed that SUA was significantly negatively correlated with pH, but possessed a positive correlation with SOM contents. There was a significant positive correlation between SUA and soil TN content. SSA/SUA/SOM/TN were positively correlated with soil bacteria, and soil pH was negatively correlated. The positive correlation factors of fungi were NH4+-N, NO3−-N, and SWC. Vegetation restoration has a significant effect on the diversity of bacteria and fungi in soil. Perhaps this is because SOM is the primary source of carbon and nitrogen for microorganisms, thus SOM affects the microbial community diversity (Liu et al., 2019). CCA showed that SOM was one of the main factors influencing bacterial community in our study. It was found that there was a significant positive correlation between bacterial diversity index and soil pH in the Antaibao open pit mining area (He et al., 2017), which was contrary to our results. In the study on the relationship between microbial population and soil fertility under different land use modes (Zhang and Liu, 2002), microbial population increased with the increase of pH value, indicating that the more acidic the soil is, the less conducive to the growth of soil microorganisms, and that most microorganisms are more suitable for neutral or slightly alkaline environment. From the relationship between the main soil nutrient index and the number of soil microorganisms, there is a positive correlation between the total number of soil microorganisms and the content of most soil nutrients, which is the result of the synergistic development of soil fertility, soil environmental state, and soil microorganisms. Healthy soil with high organic matter content and high fertility can promote the healthy growth of microorganisms (Bi and Zhang, 2014). Meanwhile, a large number of soil microorganisms, in turn, can promote the improvement of soil structure and the accumulation, transformation, and maintenance of nutrients. However, soil sucrase and urease activities increased the available carbon and nitrogen content of soil microorganisms, provided nutrients for plant growth, as a result promote the growth and reproduction of rhizosphere microorganisms (Liu et al., 2014). Through the analysis of microbial diversity, it was found that soil microbial community composition in crop-growing areas was more complex compared with crop-growing areas without crop control. There were significant differences in soil microbial community composition between planting different crops and that in crop-growing areas without crop control, but there were no significant differences in the effects of five plants on microorganisms.
All five different plants can improve soil nutrients, from soil nitrogen/pH/enzyme activity/microorganisms. The nitrogen-fixing plant GM is undoubtedly the best choice to improve soil nitrogen. Soil pH is decreased and tends to be neutral, which is more beneficial to the growth of microorganisms and plants. The rhizosphere is the region where plant roots affect the soil through the root-like deposition of exudates, mucus, and exfoliated cells. Root secretion is not only a key determinant of the structure of rhizosphere microbial community, but they also contain a variety of compounds, mainly organic acids and vitamins, as well as fatty acids, acidic amino acids, and vitamins. These compounds change the chemical properties of the soil, such as reducing the alkalinity of the soil in reclaimed areas, while providing a source of nutrients for the rhizosphere microorganisms (Turner et al., 2013; Lareen et al., 2016; Ahkami et al., 2017). These findings indicate that the release of root secretions is development specific, thereby establishing microbiota activity that enhances plant adaptability. In addition to rhizosphere secretions, microorganisms such as Nocardioides, Lactobacillus, and nitrifying bacteria also produce acids (Karatas, 2008). The interaction between plants and the associated soil microbial communities is so intimate that they can be seen as a whole, responding to environmental conditions together, and making choices (Turner et al., 2013; Lareen et al., 2016; Ahkami et al., 2017; Hortal et al., 2017). The pH value of soil is alkaline in Huainan and Huaibei regions, so it will relatively increase the pH value of soil compared with some acidic regions, because the optimal living conditions of most soil microorganisms are neutral (Marschner et al., 2011; Battini et al., 2017). Gradients of soil properties, such as soil pH, are powerful drivers of soil microbial diversity. In soil with a near-neutral pH, microorganisms grow more efficiently and have greater potential to store carbon (Hortal et al., 2017; Malik et al., 2018). Consistent with the other report (Sun et al., 2019), soil pH value and SOM are important factors that control microbial community. In alkaline soil, there was a negative correlation between SOM and pH value. The soil microbial diversity was highest when the pH value was about 7. Soil pH was the key control factor and the best predictor of soil bacterial community composition. The diversity index of bacterial community was positively correlated with the content of SOM and TN, which might be caused by restoration of vegetation in the mining subsidence area. Soil structure, organic matter, and nutrient content are the main factors affecting soil quality in mining areas. Soil microbial community plays an important role in the successful reclamation of mining areas (Thavamani et al., 2017).
Conclusions
In conclusion, this study showed that coal mining has caused serious damage to soil ecological environment in the mining area; five cultivated plants of ZM, SI, GM, MS, and SS can improve the soil nutrient, enzyme activities, and microorganisms in the surface soil of the reclamation area. Plants and microorganisms complement each other and work together to provide a more favorable soil environment for their own survival. All of these effects play a key role in remitting Panji subsidence area soil alkalinization. Finally, reasonable planting has good ecological benefits to the reclamation of tailings, and the best choice is to plant GM in a short time. Among the five plants, GM planting can improve the soil of coal mining collapse reclamation area to the greatest extent in a short time and improve the microbial community diversity.
Footnotes
Acknowledgments
The authors thank Dandan Zhao and Umar Daraz for their suggestions on this article.
Author Disclosure Statement
The authors declare that they have no conflicts of interest.
Funding Information
This work was supported by the National Key Research and Development Program of China (2018YFD0800301-01).