
Abstract
Methanogenic processes have great potential in the sustainable treatment of organic wastes with the production of methane as a renewable source of energy. However, the broader application of methanogenic processes has been hindered by process instability frequently encountered during fluctuations in operational conditions. The accumulation of organic acids, particularly propionate, is considered to be an important cause of process instability. Therefore, to gain an understanding of microbial responses during process instability, it is imperative to identify microbial populations involved in the utilization of elevated levels of propionate. In this study, microbial community analysis showed that bacterial populations from the orders of Syntrophobacterales and Clostridiales were the primary syntrophic partners in anaerobic conversion of propionate. Archaeal populations associated with Methanosaeta and Methanoculleus dominated the propionate-degrading communities enriched in methanogenic batch bioreactors. The involvement of Methanosaeta and Methanoculleus in anaerobic conversion of propionate was further supported by the close correspondence between elevated propionate and increased population abundance of Methanosaeta and Methanoculleus in continuous anaerobic digesters treating animal waste. Subsequent testing using additional methanogenic batch bioreactors revealed that the dominance of Methanosaeta and Methanoculleus populations was linked to the anaerobic degradation of elevated levels of propionate and butyrate, but not the conversion of formate, acetate, or methanol into methane. The identification of microbial populations specifically linked to anaerobic conversion of elevated propionate in this study provided much needed insight for the understanding of microbial processes relevant to process stability in methanogenic waste treatment.
Introduction
Methanogenic processes are vital in global elemental cycling, particularly given the relevance of methane as a potent greenhouse gas to climate change (Falkowski et al., 2008; Bae et al., 2018). As the end product of methanogenic conversion of organic matter, methane can also serve as a renewable source of energy. Therefore, methanogenic processes have been applied in the anaerobic treatment of diverse categories of organic waste with the simultaneous production of renewable energy, represented by anaerobic digestion as the most implemented technology (Nallathambi Gunaseelan, 1997; Nasir et al., 2012; Hagos et al., 2017). The broader adoption of methanogenic treatment processes, however, has been hindered by the difficulties in maintaining operational stability, especially during episodes of process perturbation (Chen et al., 2008; Chen et al., 2012; Yuan and Zhu, 2016).
Considerable efforts have been made to understand the underlying mechanisms of process instability in methanogenic treatment processes. Previous studies have shown that process perturbations, such as changes in operational conditions and organic loading rates, would result in process imbalance frequently characterized by pH fluctuations due to accumulation of organic acids (Ahring et al., 1995; Leitão et al., 2006; Li et al., 2014). Among the organic acids produced as intermediates during the anaerobic decomposition of organic materials, propionic acid, present as propionate at neutral pH conditions, is found to be the most persistent and inhibitory, likely due to its slower biodegradation kinetics and less favorable thermodynamics in anaerobic biotransformation into methane (Nielsen et al., 2007; Boe et al., 2010).
The anaerobic degradation of propionate under methanogenic conditions requires syntrophic interactions between methanogens and fermentative bacteria partners (Schink, 1997). Fermentative bacteria are known to convert propionate into acetate and H2 in two distinct pathways (Müller et al., 2010). The Smithella pathway, observed in Smithella spp., converts propionate to acetate and H2 by dismutation followed by β-oxidation in the following reactions:
The overall Smithella pathway has the following stoichiometry:
The classic propionate oxidation pathway is found in all other known anaerobic propionate degraders as per the following stoichiometry:
In order for both pathways to be thermodynamically favorable, the products of propionate conversion, that is, H2 and acetate, need to be maintained at low levels, which can be achieved by the utilization of H2 and acetate by methanogens in methanogenic processes. The overall stoichiometry of both anaerobic propionate oxidation pathways is identical following the consumption of H2 and acetate by hydrogenotrophic and acetoclastic methanogenesis, respectively:
Despite the large diversity of bacterial taxa known to participate in syntrophic propionate conversion, the bacterial partners specifically involved in this process have been consistently identified as populations from two bacterial orders—Syntrophobacterales and Clostridiales (Müller et al., 2010; Sieber et al., 2012). In contrast to the demonstrated specificity of these bacterial populations to anaerobic propionate conversion, much less is known about the methanogens that specifically partner with bacteria to convert the products of anaerobic propionate oxidation into methane. Given the thermodynamic necessity of maintaining adequately low H2 partial pressure to enable anaerobic propionate oxidation, hydrogenotrophic methanogens, with the ability to consume H2, have been the focus of previous studies (Stams et al., 1992; Wu et al., 1992; Shigematsu et al., 2006; Narihiro et al., 2015; Li et al., 2018). Diverse populations of methanogens have been found in methanogenic treatment processes. However, unlike the bacterial counterparts where lineages specifically linked to anaerobic propionate conversion have been identified, methanogens specifically involved in the degradation of propionate remain obscure.
With the objective of this study to identify methanogen populations specifically involved in the anaerobic conversion of propionate, methanogenic batch bioreactors were developed with elevated propionate as the sole substrate to enrich propionate-degrading microbial populations, which were profiled by 16S rRNA gene amplicon sequencing and clone library analysis. Subsequently, the roles of methanogen populations identified to specifically involve in anaerobic propionate conversion were further evaluated in continuous anaerobic digesters with episodes of elevated propionate. Findings from this study provide much needed insight into the understanding of microbial responses to elevated propionate during process perturbations in methanogenic waste treatment.
Materials and Methods
Development of methanogenic batch bioreactors
To enrich for methanogenic microbial populations specifically involved in the anaerobic conversion of propionate and other volatile organic acids (VFAs), methanogenic batch bioreactors were developed using one of the following compounds as the sole substrate: formate, acetate, propionate, butyrate, or methanol. All methanogenic batch bioreactors were set up using previously described protocols (Chen and He, 2015) and the same defined anaerobic medium prepared according to a previously described recipe (He and Sanford, 2002). The medium was autoclaved, cooled, and subsequently aliquoted into 160-mL serum bottles flushed with pure N2 and sealed with butyl rubber stoppers and aluminum caps.
The first-generation methanogenic batch bioreactors were seeded with a 10% (v/v) inoculum using the digestate from previously established continuous anaerobic digesters treating animal waste (Zhang et al., 2011) into 100 mL fresh anaerobic medium. Subsequent to inoculation, each batch bioreactor was fed with one of the following as the sole substrate: formate (50 mM), acetate (20 mM), propionate (20 mM), butyrate (20 mM), or methanol (30 mM). Additional feedings of the substrate at the same concentration were replenished when the substrate was depleted in the batch bioreactors. At the completion of 10 feedings of substrates, second-generation methanogenic batch bioreactors were established by transferring a 10% (v/v) inoculum from first-generation batch bioreactors into fresh medium followed by repeated feedings of the same substrate. At the conclusion of 10 feedings, the second-generation methanogenic batch bioreactors were sampled for microbial community analysis. All batch bioreactors were set up in triplicates and maintained in a shaking incubator at 80 rpm and 35°C with periodic monitoring of methane production and substrate concentration.
Configuration and operation of continuous anaerobic digesters treating animal waste
To identify methanogenic microbial populations specifically involved in anaerobic conversion of propionate, population dynamics of methanogens were monitored during two episodes of elevated propionate in previously established triplicate continuous anaerobic digesters (Chen et al., 2012). Dairy wastewater was used as the regular feedstock for these continuous anaerobic digesters at an organic loading rate of 1.0 g volatile solids (VS)/L/day. The operating temperature was controlled at 35°C with the hydraulic retention time maintained at 20 days throughout the study period. All continuous digesters exhibited stable operation before the two episodes of elevated propionate.
The first episode of elevated propionate was introduced into the anaerobic digesters with the direct addition of propionate without disrupting the regular feeding of dairy wastewater. To maintain propionate at elevated levels, a sodium propionate stock solution (5 M) was used to feed the triplicate continuous anaerobic digesters at a loading rate of 6.0 mM/day for 2 days. Propionate levels in the anaerobic digesters were closely monitored and the propionate loading was increased to 9.0 mM/day in order for the peak propionate level to reach 12.0 mM (Fig. 1). To replicate the microbial responses to elevated propionate, a 2nd episode of elevated propionate was introduced 10 days after the completion of the 1st episode of elevated propionate. Propionate was added at a loading rate of 9.0 mM/day for 8 days, resulting in elevated propionate levels similar to those in the 1st episode of elevated propionate (Fig. 1). The loading rate of dairy wastewater remained unchanged at 1.0 g VS/L/day during the study period with or without the addition of extraneous propionate.
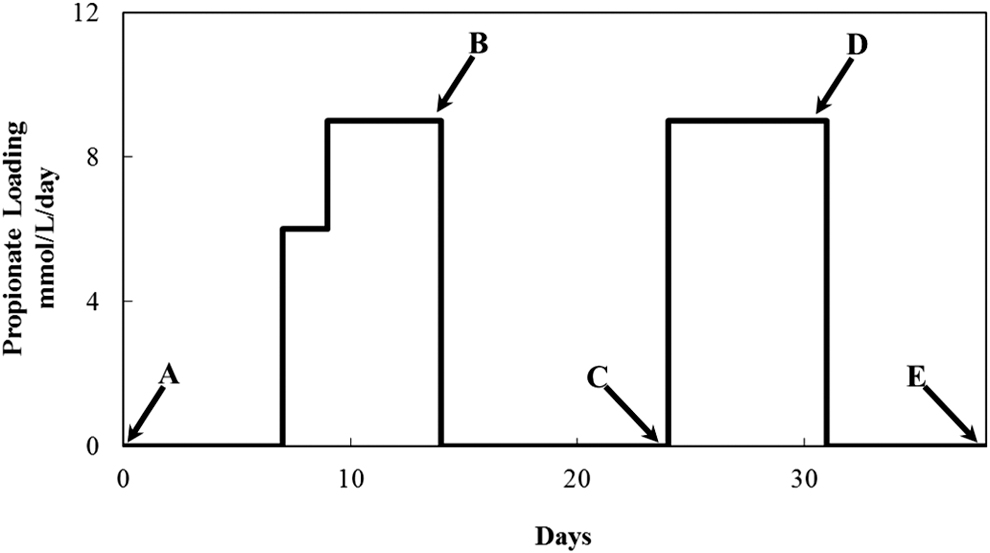
The addition of extraneous propionate to continuous anaerobic digesters during the two episodes of elevated propionate. Biomass sampling at five time points was shown by the arrows labeled with letters A to E. Feeding of dairy wastewater to the anaerobic digesters remained constant at 1.0 g VS/L/day throughout the study period. VS, volatile solids.
Process parameters of the continuous anaerobic digestions, including CH4 production and VFA concentrations, were monitored as described previously (Chen et al., 2012). Biomass samples were taken from the continuous anaerobic digesters at five time points A–E as illustrated in Fig. 1. Samples were pelleted by centrifugation at 14,000 g for 15 min and preserved at −80°C for further processing.
High-throughput sequencing of 16S rRNA gene amplicon library
The microbial communities in the methanogenic batch bioreactors with propionate as the sole substrate were analyzed by 16S rRNA gene amplicon library sequencing. First, whole community DNA was extracted from the biomass samples and purified as previously described (Zhang et al., 2009). DNA extracts from triplicate batch bioreactors were pooled for polymerase chain reaction (PCR) amplification of the V4 region of the 16S rRNA gene with 515-F and 806-R Golay barcoded primers (Caporaso et al., 2012). PCR amplification and cleaning, amplicon quantification, and paired-end sequencing were completed with previously established protocols (Chen et al., 2017). All sequence reads were analyzed by the Mothur (v.1.35) platform following previously established procedures (Kozich et al., 2013).
After sequence quality processing, including barcode and primer trimming, denoising, and chimera checking, operational taxonomic units (OTUs) were assigned by a 97% identity threshold using previously established procedures (Zhang and He, 2013) followed by assignment of taxonomic ranks with the RDP Classifier at the confidence threshold of 80% as previously described (Cole et al., 2003). Community compositions were estimated according to the taxonomic assignments of all valid sequences.
Clone library analysis of methanogenic batch bioreactors
The methanogen community in the propionate-amended methanogenic batch bioreactors was also characterized by clone library analysis following previously established protocols (Zhang and He, 2013). Clone library analysis was performed to verify results from the high-throughput sequencing of 16S rRNA gene amplicon library and to obtain more complete 16S RNA gene sequences for more accurate taxonomic classification. Briefly, DNA extracts from the second-generation batch bioreactors were used for PCR amplification of the 16S rRNA genes with archaea-specific primers, Arch21F and Arch958R, as previously described (DeLong, 1992). Amplicons were subsequently purified and cloned into plasmid vectors following previously described procedures (Zhang et al., 2011). Approximately 20 cloned plasmid inserts were randomly selected for sequencing.
The resulting 16S rRNA gene sequences were checked for chimeric artifacts using the Chimera Check program in the Ribosomal Database Project II (Cole et al., 2003), with high-quality sequences deposited at GenBank under the following accession numbers: KJ914860—KJ914878. Subsequently, OTUs with 3% difference cutoff were assigned based on the average neighbor clustering algorithm. Phylogenetic analysis was conducted on representative 16S rRNA gene sequences of OTUs classified as Methanosaeta and Methanoculleus with closely related sequences in the NCBI GenBank database using MEGA 4.0 (Tamura et al., 2007).
Quantification of methanogen populations by real-time quantitative PCR
Microbial populations of interest were quantified by real-time quantitative PCR (qPCR) in the methanogenic batch bioreactors and continuous anaerobic digesters during two episodes of elevated propionate. All qPCR assays used the TaqMan chemistry following previously established protocols (Chen and He, 2016). The population-specific TaqMan primer/probe sets used in the qPCR assays included Mst702F-Mst753P-Mst862R (Yu et al., 2005), Mc274F-Mc361P-Mc477R (Chen et al., 2014), and Arc787F-Arc915P- Arc1059R (Yu et al., 2005), for the quantification of Methanosaeta, Methanoculleus, and total archaea, respectively. DNA templates used as the standards for qPCR were partial 16S rRNA genes of representative methanogen populations cloned from the continuous anaerobic digesters in a previous study (Chen et al., 2014), including Methanosaeta (GenBank Accession No. JN052761) and Methanoculleus (GenBank Accession No. JN052756).
Analytical methods
Biogas production from the batch bioreactors or continuous anaerobic digesters was measured with a previously described water-displacement technique (Zhu et al., 2011). CH4 content in biogas was determined with a Hewlett Packard 5890 Series II gas chromatograph equipped with a thermal conductivity detector and a Supelco packing column (60/80 Carbonxen®-1000; Sigma-Aldrich, St Louis, MO) according to a previously described operating procedure (Chen and He, 2016). VFAs were quantified with an Agilent 1200 series High-Performance Liquid Chromatography (Agilent Technologies, Santa Clara, CA) equipped with a Bio-Rad Aminex HPX-87H ion exclusion column (Bio-Rad, Hercules, CA) as previously described (Chen and He, 2016).
Statistical analysis
To evaluate the response of specific methanogen populations to elevated propionate levels, the changes in the abundance of Methanosaeta as well as Methanoculleus during the two episodes of elevated propionate in the continuous anaerobic digesters were analyzed with the one-way analysis of variance (ANOVA) using qPCR results. Significant differences in the abundance of Methanosaeta/Methanoculleus between time points, that is, time points A–E (Fig. 1), with or without elevated propionate, were indicated by a probability value (p) less than 0.05 in ANOVA analysis. Post hoc comparisons were performed using Tukey's test to further identify the time points with significant difference in Methanosaeta/Methanoculleus abundance as indicated by a probability value less than 0.05. In addition, correlations of the abundance between Methanosaeta and Methanoculleus in the continuous anaerobic digesters subjected to two episodes of elevated propionate were evaluated with the Pearson's correlation.
Results and Discussion
Propionate utilization in methanogenic batch bioreactors
High levels of propionate (20 mM) were used as the sole substrate to enrich methanogenic populations involved in anaerobic conversion of propionate. In the second-generation batch bioreactors amended with propionate, the utilization of propionate was accompanied with proportional increases in cumulative methane production (Supplementary Fig. S1). The methane yield from propionate averaged 1.61 mmol CH4/mmol propionate, which was 92% of the theoretical methane yield according to the overall stoichiometry of anaerobic propionate conversion to methane [Eq. (5)].
The agreement between experimental results and theoretical predictions on methane yields from propionate confirmed that the methanogenic populations enriched in the batch bioreactors were indeed involved in the syntrophic conversion of propionate. Therefore, these batch bioreactors were further studied to characterize the methanogen populations grown on propionate as the substrate.
Methanogen populations in methanogenic batch bioreactors
The methanogen community in the propionate-amended batch bioreactors was profiled by 16S rRNA gene amplicon library sequencing. With archaeal sequences accounting for 40.1% of the overall microbial community, populations associated with Methanosaeta and Methanoculleus were found to dominate the archaeal community, with a combined relative abundance of 88% (Fig. 2a). Minor populations of methanogens included Methanospirillum and Methanobacterium. The predominance of Methanosaeta and Methanoculleus suggested that Methanosaeta as acetoclastic methanogens likely utilized acetate as a product of propionate oxidation, while Methanoculleus as hydrogenotrophic methanogens consumed H2 to sufficiently low levels to sustain anaerobic biodegradation of elevated propionate.

Composition of the archaeal community in methanogenic batch bioreactors with propionate as the sole substrate. The relative abundance of methanogen taxa is shown as percentage values determined by 16S rRNA gene amplicon library sequencing
To achieve more accurate phylogenetic classification of Methanosaeta and Methanoculleus populations identified by 16S rRNA gene amplicon library sequencing, the methanogen community was further characterized with clone library analysis in the propionate-amended batch bioreactors. As expected, Methanosaeta and Methanoculleus were again found to be the dominant methanogen populations (Fig. 2b). Combined, these two populations accounted for 95% of the archaeal community, which was consistent with the results from 16S rRNA gene amplicon library sequencing. The nearly exclusive dominance of Methanosaeta and Methanoculleus in the methanogen community further suggested the superior competitiveness and specific involvement of both methanogen populations in the conversion of elevated propionate.
Phylogenetic analysis of representative clones of 16S rRNA genes indicated that all clones of Methanosaeta were closely related to Methanosaeta concilii (Fig. 3), which has been identified as a primary acetoclastic methanogen population involved in the degradation of propionate in anaerobic digestion processes (Shigematsu et al., 2006; Ban et al., 2013; Kim et al., 2015; Zhang et al., 2018). The majority of Methanoculleus clones had Methanoculleus receptaculi as the closest relative (Fig. 3). M. receptaculi is a strictly hydrogenotrophic methanogen (Cheng et al., 2008) and has been found to be important members of propionate-degrading consortia (Ban et al., 2013; Ahlert et al., 2016). These observations corroborated the potential competitiveness and specific involvement of Methanosaeta and Methanoculleus in anaerobic propionate conversion.
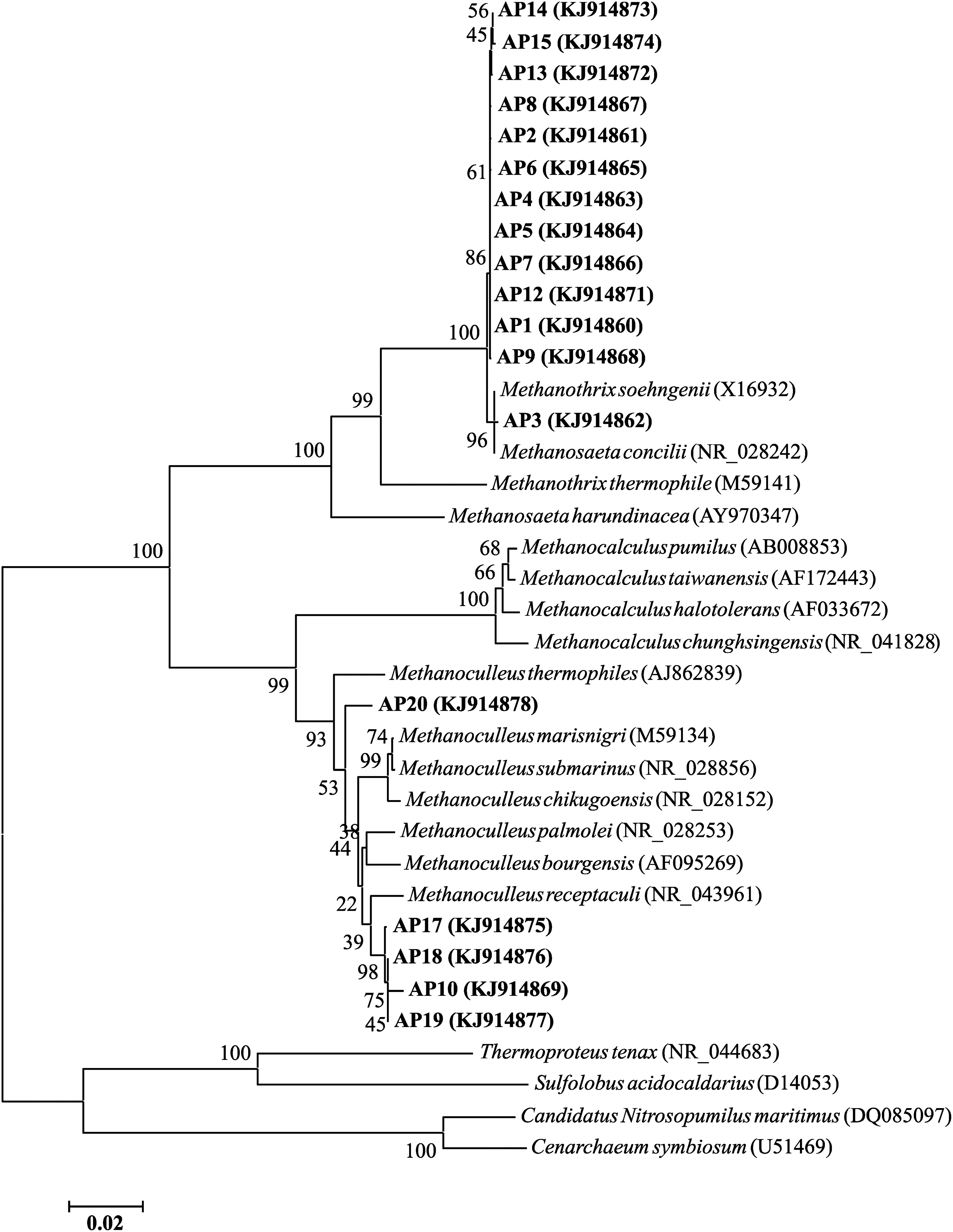
Neighbor-joining phylogenetic tree showing relationships of representative partial 16S rRNA gene sequences cloned from methanogenic batch bioreactors with propionate as the sole substrate to close relatives. Clones from this study are in bold. GenBank accession numbers of the 16S rRNA gene sequences are indicated in the parentheses. The numerical values at branch nodes indicate bootstrap values per 1,000 re-samplings. The scale bar represents the number of substitutions per sequence position.
Bacterial populations in methanogenic batch bioreactors
Bacterial sequences accounted for 59.9% of the overall microbial community in the propionate-amended batch bioreactors as profiled by 16S rRNA gene amplicon library sequencing. Populations associated with Syntrophobacter and Peptococcaceae represented the majority of the bacterial community, with a combined relative abundance of 57% (Supplementary Fig. S2). It should be noted that Syntrophobacter and Peptococcaceae belong to the bacterial orders of Syntrophobacterales and Clostridiales, respectively, which are known to be involved in anaerobic propionate conversion as syntrophic partners (Müller et al., 2010; Sieber et al., 2012).
Other bacterial populations contributing more than 3% to the bacterial community participating in anaerobic propionate conversion included sequences classified as Thermovirga and Thermotogaceae (Supplementary Fig. S2), which have been frequently identified in methanogenic processes, however, with specific involvement in propionate conversion remained to be validated (Amin et al., 2021). Analysis of the bacterial populations in the propionate-amended batch bioreactors confirmed the abundance and importance of Syntrophobacterales and Clostridiales in syntrophic propionate degradation. However, further efforts are needed to understand the roles of other minor, but significant bacterial populations such as Thermovirga in anaerobic propionate conversion.
Propionate utilization in continuous anaerobic digesters treating animal wastewater
The continuous anaerobic digesters used animal wastewater as the substrate. As a result, the microbial community in the anaerobic digesters was much more complex than that in the batch bioreactors (Chen et al., 2012), providing ideal process conditions to further validate the competitiveness and specific involvement of Methanosaeta and Methanoculleus in anaerobic conversion of elevated propionate.
To identify methanogen populations specifically involved in the biodegradation of high concentrations of propionate, two episodes of elevated propionate was introduced to the continuous anaerobic digesters by the addition of extraneous propionate (Fig. 1). During both episodes of elevated propionate, the additions of propionate led to rapid increases in propionate concentration, peaking at about 12 mM (Fig. 4a). The concentration of propionate declined immediately following the cessation of propionate feeding, to levels before propionate addition. Corresponding to the increases in propionate concentration, methane production increased and subsequently declined when propionate concentration decreased (Fig. 4b), indicating the rapid conversion of propionate into methane. Notably, there was no indication of acetate accumulation in the anaerobic digesters in response to propionate addition (Fig. 4a), further demonstrating the effectiveness of the methanogen community in the utilization of intermediates from anaerobic propionate oxidation.
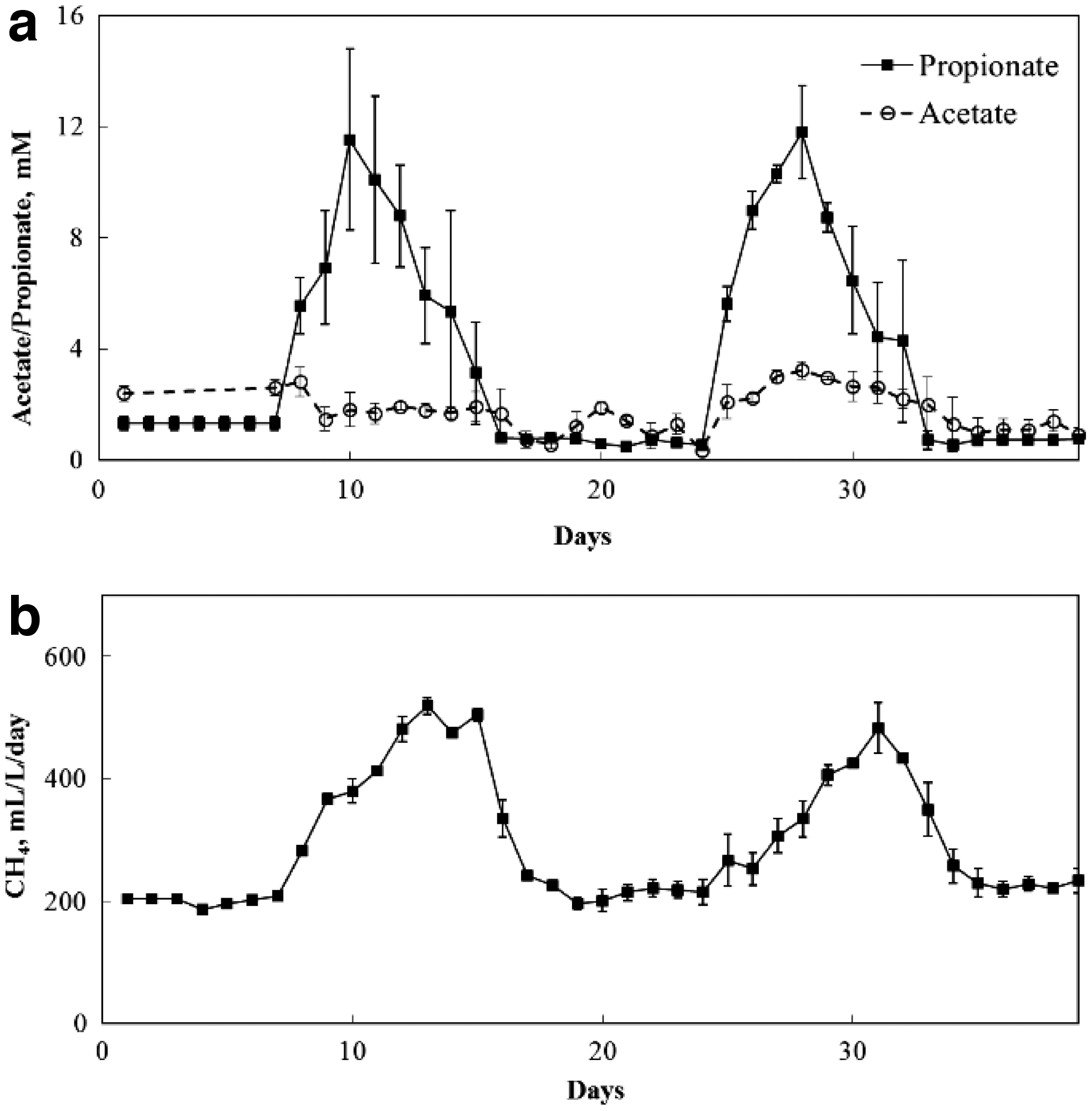
Performance of continuous anaerobic digesters during two episodes of elevated propionate:
Dynamics of Methanosaeta and Methanoculleus in continuous anaerobic digesters in response to elevated propionate
Microbial community analysis of propionate-amended methanogenic batch bioreactors suggested the competitiveness and specific involvement of Methanosaeta and Methanoculleus in the biodegradation of high levels of propionate (Fig. 2a, b). It should be pointed out that in treatment processes using complex waste materials as the substrate, Methanosaeta and Methanoculleus might not be the dominant methanogens even with propionate accumulation, due to competitions from other methanogen populations that utilize various intermediates other than propionate. Thus, the continuous anaerobic digesters using animal wastewater as the substrate provided a relevant model process to simulate this scenario.
The specific responses of Methanosaeta and Methanoculleus populations to elevated propionate were monitored by qPCR in the continuous anaerobic digesters, where effective propionate conversion into methane was observed (Fig. 4). Methanosaeta- and Methanoculleus-specific qPCR assays showed that both methanogen populations responded positively to the addition of extraneous propionate. When the anaerobic digesters experienced the first episode of elevated propionate with an increase in propionate concentration from time point A to B (Fig. 1), the abundance of both Methanosaeta and Methanoculleus increased significantly (Fig. 5a, b). Afterward, the pause in propionate addition at time point B (Fig. 1) and subsequent drop in the propionate concentration (Fig. 4a) led to the significant decline in the abundance of both Methanosaeta and Methanoculleus from time point B to C (Fig. 5a, b). The same population dynamics were again demonstrated during the second episode of elevated propionate from time point C to E (Fig. 5a, b), suggesting the specific involvement of Methanosaeta and Methanoculleus in the conversion of elevated propionate to methane.
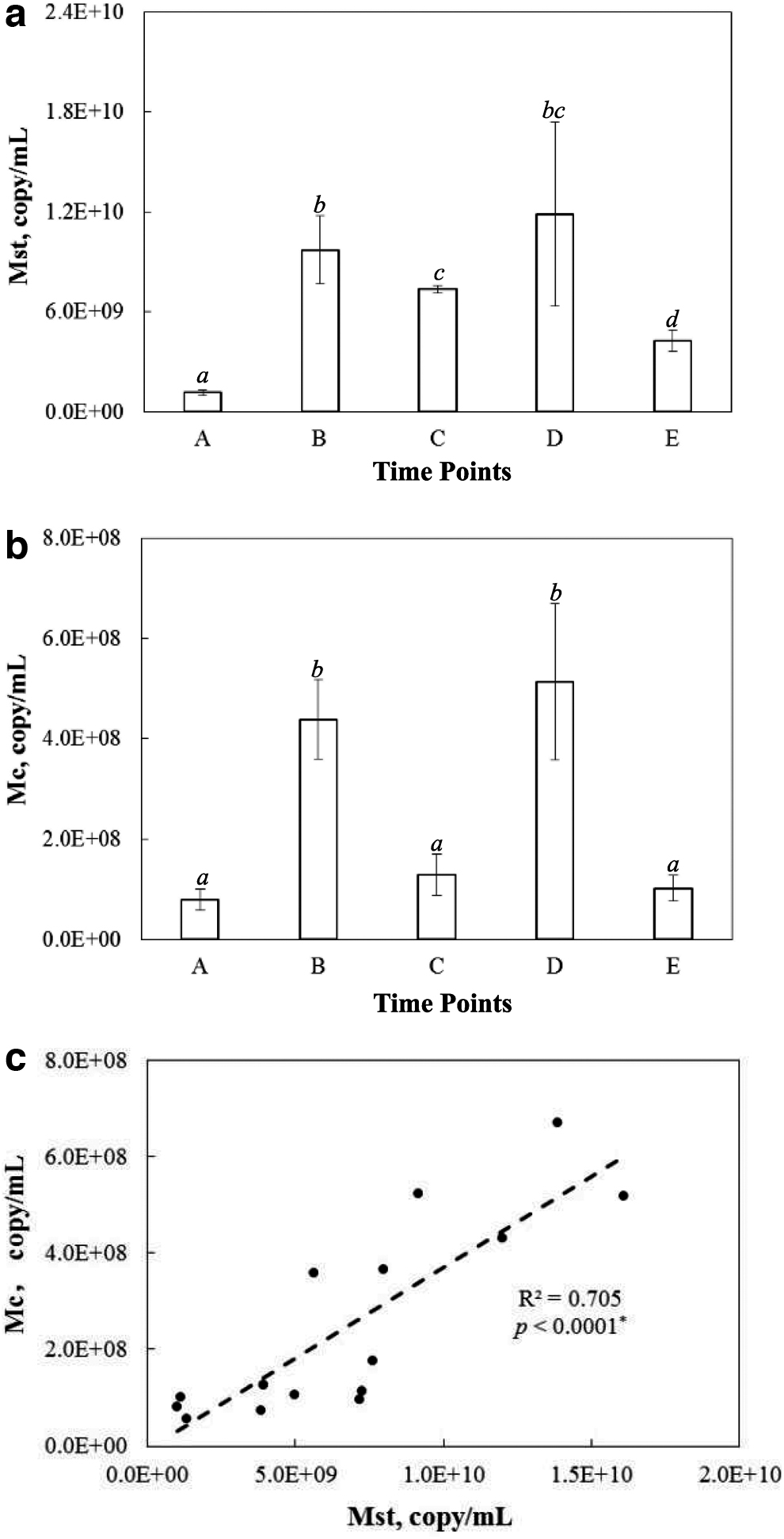
Abundance of
Moreover, the population dynamics of Methanosaeta and Methanoculleus were correlated with a statistically significant correlation coefficient of 0.7, demonstrating the coordinated responses to elevated propionate between Methanosaeta and Methanoculleus in the continuous anaerobic digesters (Fig. 5c). The close coordination between Methanosaeta and Methanoculleus might explain the combined dominance of Methanosaeta and Methanoculleus in the propionate-degrading microbial communities observed in the methanogenic batch bioreactors with propionate as the sole substrate (Fig. 2).
Specificity of Methanosaeta and Methanoculleus to propionate conversion
The involvement of Methanosaeta and Methanoculleus in the conversion of propionate to methane was demonstrated by microbial community analysis of propionate-amended batch bioreactors by 16S rRNA gene sequencing and continuous anaerobic digesters by qPCR (Figs. 2 and 5). It remained to be verified whether the involvement of Methanosaeta and Methanoculleus was specific to propionate. Therefore, additional methanogenic batch bioreactors were set up using other organic compounds, including formate, acetate, butyrate, and methanol. The abundance of Methanosaeta and Methanoculleus was determined by qPCR and compared between batch bioreactors using different compounds as the sole substrate.
As expected, qPCR analysis revealed that Methanosaeta and Methanoculleus populations dominated in the propionate-amended batch bioreactors, accounting for a combined 87% of the archaeal community (Fig. 6), consistent with results from 16S rRNA gene amplicon library sequencing and clone library analysis (Fig. 2). Interestingly, the dominance of Methanosaeta and Methanoculleus was also found in the butyrate-amended batch bioreactors (Fig. 6). Previous studies have shown that anaerobic oxidation of butyrate uses β-oxidation to convert butyrate to acetate and H2 (Schink, 1997) with the following stoichiometry:
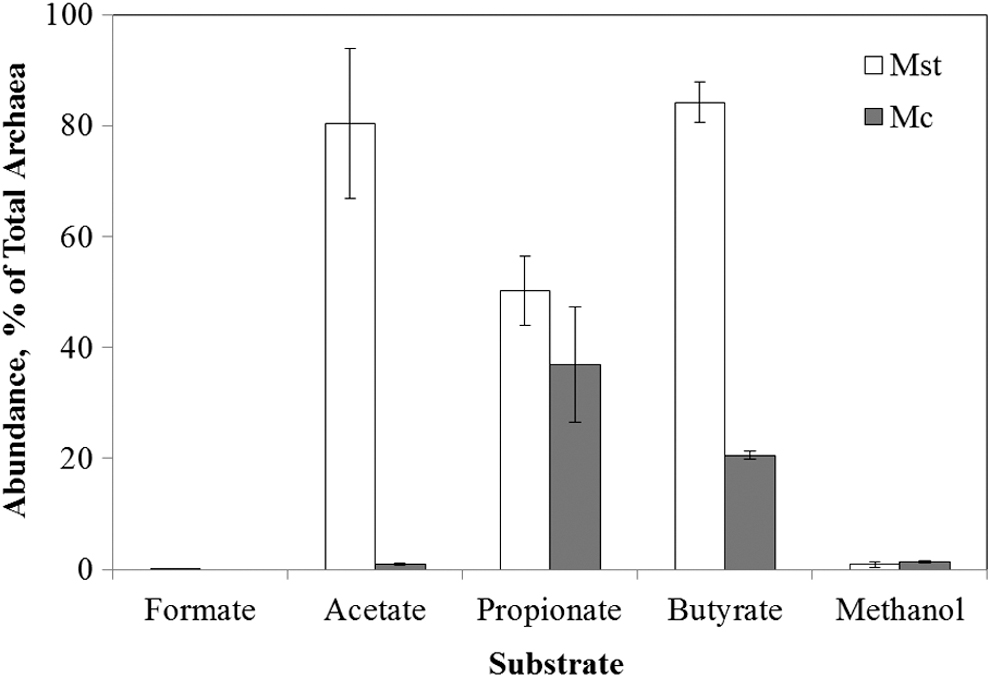
Abundance of Methanosaeta (Mst) and Methanoculleus (Mc) as a percentage of total archaea in methanogenic batch bioreactors amended by various organic acids or methanol as the sole substrate. Microbial abundance was determined as the copies of population-specific 16S rRNA genes with qPCR. Results were means of triplicates with the error bars showing standard deviations.
The products of anaerobic butyrate oxidation, that is, acetate and H2, were the same as those of propionate oxidation [Eqs. (3) and (4)]. Thus, this similarity between the conversions of propionate and butyrate likely contributed to the dominance of Methanosaeta and Methanoculleus in both propionate- and butyrate-amended batch bioreactors. These observations pointed to the possibility that Methanosaeta and Methanoculleus were specifically linked to the anaerobic conversion of organic compounds with acetate and H2 as the primary intermediate products.
It was observed that the relative abundance of acetoclastic Methanosaeta was higher in butyrate-amended batch bioreactors compared with the propionate-amended batch bioreactors (Fig. 6). In comparison, hydrogenotrophic Methanoculleus exhibited the opposite trend (Fig. 6). According to Equation (6), anaerobic conversion of butyrate would set the molar ratio of acetate/H2 in the product mix at 1.00. In contrast, anaerobic conversion of propionate following the classic pathway [Eq. (4)] would yield an acetate/H2 molar ratio of 0.33. The molar ratio of acetate/H2 represents the distribution of substrates between acetoclastic methanogens (i.e., Methanosaeta) and hydrogenotrophic methanogens (i.e., Methanoculleus). Thus, the higher acetate/H2 ratio from anaerobic conversion of butyrate than propionate is consistent with the greater relative abundance of Methanosaeta observed in butyrate- than propionate-amended batch bioreactors (Fig. 6).
It should be noted that anaerobic conversion of propionate could follow the Smithella pathway [Eq. (3)], yielding an acetate/H2 molar ratio of 1.50, which is higher compared with anaerobic butyrate conversion [Eq. (6)]. Thus, the prevalence of the Smithella pathway would have resulted in greater relative abundance of Methanosaeta in propionate- rather than butyrate-amended batch bioreactors, which, however, was not the case (Fig. 6). It could be deduced that the contribution of the Smithella pathway to anaerobic propionate conversion was insignificant in propionate-amended batch bioreactors, which is further corroborated by the lack of detection of Smithella in propionate-amended bioreactors (Supplementary Fig. S2).
In the acetate-amended batch bioreactors, qPCR results showed that Methanosaeta represented more than 80% of the archaeal community, while the abundance of Methanoculleus was negligible (Fig. 6). These results were expected as acetate could be converted directly into methane by Methanosaeta populations alone as obligatory acetoclastic methanogens (Smith and Ingram-Smith, 2007). The absence of Methanosaeta and Methanoculleus was expected in the methanol-amended batch bioreactors, which was consistent with the inability to perform methylotrophic methanogenesis by either Methanosaeta or Methanoculleus (Maestrojuan et al., 1990; Smith and Ingram-Smith, 2007).
The absence of Methanoculleus in the formate-amended batch bioreactors (Fig. 6), however, was unexpected. With rare exceptions, the ability to utilize formate for methanogenesis has been found in almost all Methanoculleus species (Maestrojuan et al., 1990), including M. receptaculi (Cheng et al., 2008), with which many of the clones from the propionate-amended batch bioreactors were closely associated (Fig. 3). It is evident that Methanoculleus populations in the batch bioreactors were not able to compete for formate with other hydrogenotrophic methanogens in this study. Instead, it is possible that these Methanoculleus populations might be specifically competitive in the utilization of H2 at very low partial pressure associated with the syntrophic oxidation of propionate or butyrate, as evidenced by the dominance of Methanoculleus as the hydrogenotrophic methanogens in propionate- and butyrate-amended batch bioreactors. Indeed, Methanoculleus populations have been shown to have high affinity to H2 in previous studies (Hori et al., 2006; Sakai et al., 2009). Notably, Methanosaeta populations are also known to have high affinity to acetate and outcompete other acetoclastic methanogens at low acetate levels (Conklin et al., 2006). Thus, the involvement of Methanosaeta and Methanoculleus in the anaerobic conversion of propionate or butyrate could likely be attributed to the ability of these methanogens to utilize H2 and acetate at low concentrations, a thermodynamic requirement for anaerobic oxidation of VFAs.
Conclusions
In this study, Methanosaeta and Methanoculleus were identified as the key acetoclastic and hydrogenotrophic methanogens, respectively, participating in the anaerobic degradation of propionate as a central intermediate in methanogenic processes. Bacterial populations from the orders of Syntrophobacterales and Clostridiales were confirmed as the primary syntrophic partners in anaerobic conversion of propionate utilizing the classic anaerobic propionate oxidation pathway. Findings from this study provide much needed insight into the understanding of microbial responses to elevated propionate, which is recognized as an inhibitory intermediate frequently contributing to process perturbations in methanogenic waste treatment. The identification of microbial populations specifically involved in anaerobic propionate degradation suggests the potential of exploiting microbial populations such as Methanosaeta and Methanoculleus in enhancing the stability of methanogenic treatment processes.
Footnotes
Authors' Contributions
Q.H. and S.C. conceived and designed research. S.C., Y.W., and H.C. conducted experiments. S.C. and C.H. analyzed data. Q.H. and T.C.H. wrote the article. All authors read and approved the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported, in part, by U.S. National Science Foundation (NSF) award 2025339. S.C. was partly supported by the Institute for a Secure and Sustainable Environment at the University of Tennessee, Knoxville. The opinions, findings, recommendations, and conclusions, if any, in this article are those of the authors, and do not necessarily reflect the views of NSF or University of Tennessee, Knoxville.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.