
Abstract
Salmonella shedding in many livestock species can increase significantly after transport and lairage. Preprocessing increases in shedding can amplify the amount of Salmonella that enters the processing facility and the likelihood of end-product contamination. We previously produced an anti-Salmonella phage cocktail that reduced colonization in swine when the pigs were exposed to an environment heavily contaminated with Salmonella, similar to what might be seen in a transport trailer or processing facility holding pen. The aim of this study was to increase the efficacy of the phage treatment by (1) expanding the host-range of the cocktail and (2) developing a more cost-effective microencapsulation technique. We collected samples from wastewater treatment facilities and isolated 20 distinct phages belonging to either the Siphoviridae or Myoviridae families. From this library we identified 10 phages that together lysed a mixed culture of Salmonella enterica Typhimurium, Enteriditis, and Kentucky—three serovars commonly associated with meat and poultry products. The phages were microencapsulated using two sodium-alginate-based methods that only reduced the cocktail titer by 1.0–1.5 logs (premicroencapsulation: 10.4 log10 PFU/mL; postmicroencapsulation method one: 9.2 log10 PFU/mL; postmicroencapsulation method two: 8.9 log10 PFU/mL). Microencapsulated phages remained stable at both 4°C and 22°C for up to 14 days with no appreciable drop in titer (mean titer: 8.9 log10 PFU/mL). These data indicate that phage cocktails with wider host ranges are possible and a cost-effective microencapsulation process can protect the phages over an extended period, making simultaneous treatment of large numbers of animals with feed- or water-based delivery possible.
Introduction
S
A growing number of groups are working to develop phage therapy as a food safety intervention strategy. While the majority of such work has focused on poultry (Barrow et al., 1998; Fiorentin et al., 2005; Loc Carillo et al., 2005; Toro et al., 2005; Wagenaar et al., 2005; Hurley et al., 2008; Borie et al., 2009; El-Shibiny et al., 2009), a smaller number of groups have focused on swine (Smith and Huggins, 1983; Lee and Harris, 2001) and cattle (Callaway et al., 2008). The majority of these studies indicate that phage therapy can effectively limit colonization of different foodborne pathogens in livestock.
We previously showed that a cocktail of 15 phages, isolated from wastewater treatment facilities based on their ability to lyse Salmonella enterica Typhimurium, reduced Salmonella shedding in pigs under simulated production settings (Wall et al., 2010). The goal of the current study was to increase the efficacy of our phage treatment by expanding its spectrum to include other S. enterica serovars commonly associated with Salmonella infections in food animals or humans. In addition, we aimed to produce a more practical microencapsulation method to protect the phages from the gastric environment, ensuring that viable phages reach the actual sites of infection. This new microencapsulation method makes the simultaneous treatment of multiple animals through feed or water possible, which will be necessary in successfully transferring this technology to the livestock industry.
Materials and Methods
Salmonella strain
S. enterica Typhimurium γ4232 was isolated as previously described (Wall et al., 2010). S. enterica Enteriditis and S. enterica Kentucky isolates were kindly provided by Dr. Arun Bhunia (Purdue University, West Lafayette, IN).
Wild-type phage isolation
Twenty wild-type phages were isolated from waste water treatment plants located throughout Indiana. The phages were isolated based on their ability to lyse the different Salmonella serovars using a previously described method (Wall et al., 2010). Briefly, wastewater samples were centrifuged (2000 g) for 10 min. The supernatant was filtered using 0.2 μm syringe filters and treated with chloroform (1:100) to kill residual bacteria. Phages were enriched by combining the supernatant with S. enterica Typhimurium grown to log phase in 2× tryptic soy broth and incubated at 37°C overnight. The overnight culture was centrifuged (2000 g) and the supernatant was filtered as before. Filtered samples (100 μL) were added to 100 μL of fresh log-phase Salmonella, 67.5 μL of 1 M CaCl2, and 3 mL nutrient overlay. The phage/nutrient overlay was added to nutrient agar plates and incubated overnight at 37°C. Individual plaques were isolated and plaque purified (3×) and stored at −80°C until further use.
Host-range analysis
Individual phages were tested for their ability to lyse Salmonella serovars Typhimurium, Kentucky, and Enteriditis by standard plaque assay. Each Salmonella strain was grown individually to log phase and combined with the individual phages as described above. The coculture was incubated overnight and individual phage titers were recorded.
In concurrent experiments, the phages were combined together (based on volume) into a cocktail and tested for their ability to lyse a mixed culture of the three test Salmonella serovars together. Here plaque assays were performed as described above using the phage cocktail and a combination (based on volume) of log-phase growth Salmonella serovars Typhimurium, Kentucky, and Enteriditis.
Phage cocktail preparation and microencapsulation
Salmonella Typhimurium was grown to log-phase growth in 100 mL flasks of nutrient broth. The flasks were inoculated with individual phages and incubated overnight at 37°C with shaking. Overnight samples were centrifuged, filtered, and treated with chloroform as described above.
Phages were first microencapsulated using the sodium alginate method as previously described (Wall et al., 2010). Briefly, 3.5 mL (∼10 log10 PFU/mL) of phage preparation was suspended in phosphate-buffered saline and slowly mixed with 14 mL of Span-85 followed by 70 mL of canola oil. The mixture was emulsified at 5500 rpm for 1 min. With continued mixing, 17.5 mL of 0.5% CaCl2 and 0.05% ZnCl2 was added drop wise and mixed for an additional 2 min. The microspheres were pelleted at 1000 g for 15 min. The oil was decanted and the pellets were washed in three volumes water (30 mL) and pelleted again. The pellet was resuspended in one volume water and two volumes 0.2% poly-L-lysine (microencapsulation method one). Microspheres were pelleted once again, resuspended in PBS and stored at 4°C until further use. Phages were also microencapsulated using the above-described protocol excluding poly-L-lysine in attempt to produce a more affordable method (microencapsulation method two). Plaque assays were then performed on the microencapsulated phages (as described above) at 0 h, 24 h, 72 h, 7 days, and 14 days at both 4°C and 22°C to determine viability of the phages over time.
Characterization of phages
Phages were characterized by both light and transmission electron microscopy (TEM). For light microscopy, droplets of the sample were deposited on glass slides and coverslipped before imaging. Images were generated using an Olympus Vanox light microscope using Nomarski differential interference contrast filtering. Initial magnification was with a 20× objective. Images were captured with final horizontal field width of ∼560 μm using a Qimaging Micropublisher 5.0 RTV camera. For TEM, a 300 mesh with a formvar+ carbon film grid was floated for ∼1 min on individual phage samples before being stained with 2% uranyl acetate. The Philips CM-100 TEM was operated at 80 kv, spot 2, 200 μm condenser aperture, and 50 μm objective aperature. Images were magnified at 15,000× and 28,500×.
Results
Phage isolation and characterization
Twenty phages were isolated from wastewater samples obtained from 14 different wastewater treatment plants throughout Indiana. These phages were isolated initially based on their ability to lyse S. enterica Typhimurium. Each individual phage was then screened for its ability to lyse S. enterica serovars Kentucky and Enteriditis as well. From the initial group of 20, 10 phages were identified that lysed the different serovars to varying degrees (Table 1). The 10 phages selected were obtained from 10 individual wastewater treatment plants. All 10 phages effectively lysed S. enterica Typhimurium producing titers ranging from 7.2 to 10.8 log10 PFU/mL. Nine of the 10 phages lysed S. enterica Enteriditis producing titers ranging from 6.1 to 10.8 log10 PFU/mL. Only 5 of the 10 phages lysed S. enterica Kentucky. These phages also produced comparatively lower titers ranging from 6.5 to 9.0 log10 PFU/mL. When combined together, the phage cocktail was highly lytic against a mixed culture of Salmonella Typhimurium, Kentucky, and Enteriditis producing a titer of 10.4 log10 PFU/mL by plaque assay (Table 2).
Each phage was grown individually on each of the different serovars. Plaque assays were performed as described in the Materials and Methods section. Numerical values are phage concentrations (log10 PFU/mL).
The 10 phages were combined into a cocktail (equal parts based on volume) and microencapsulated. Plaque assays were performed on a mixed culture of Salmonella enterica Typhimurium, Enteriditis, and Kentucky as described in the Materials and Methods section.
Plaque and phage morphology
All 10 phages produced similar plaques on Salmonella Typhimurium and Enteriditis (round; between 1 and 2 mm). Plaques formed on Salmonella Kentucky were larger (>3 mm) with more diffuse edges. The 10 phage isolates were observed by TEM. All phages were nonenveloped, tailed viruses but with two distinct morphologies. Phages CHI, NWI, LI, WI, and CV had long, thin (noncontractile) tails and were ∼400 nm in length, indicating that they belonged to the Siphoviridae family of viruses (Ackerman, 2005). Phages VI, DR, BE, BL, and CI had short, thick (contractile) tails and were ∼200 nm in length (Fig. 1), indicating that they belonged to the Myoviridae family of viruses (Ackerman, 2005).
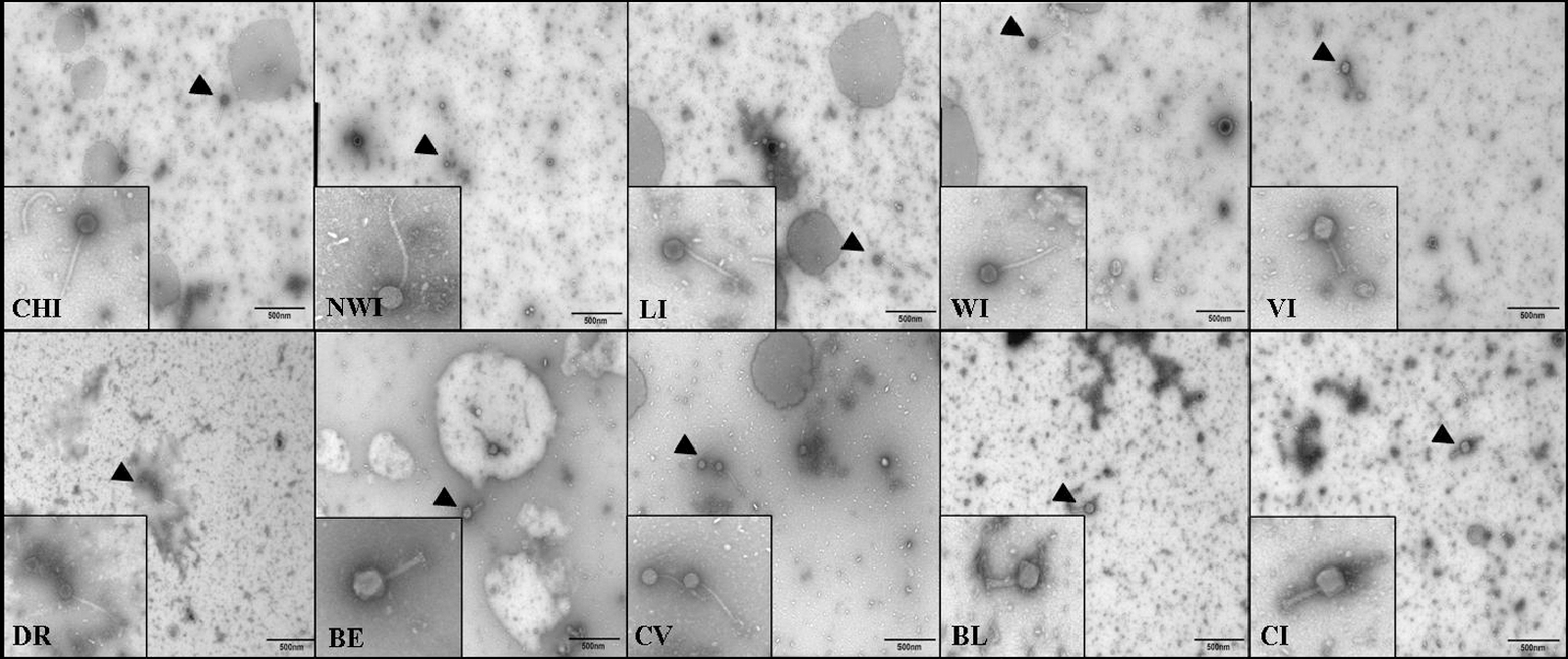
Electron micrographs of phages isolated from wastewater treatment samples. The 10 phages were isolated from 10 wastewater treatment centers. Initials indicate the name of each phage. Black triangles point to 28,500× images of the phages magnified in the smaller squares. Phages CHI, NWI, LI, WI, DR, and CV display morphologies representative of Siphoviridae. Phages VI, BE, BL, and CI display morphologies representative of Myoviridae.
Microencapsulation
Two microencapsulation protocols were tested. Both were sodium-alginate based with one containing poly-L-lysine (method one) and one not containing poly-L-lysine (method two). In each case, the 10 phages were combined into a cocktail (based on volume) before microencapsulation. Microencapsulation produced moderately uniform microspheres as evidenced by light microscopy (Fig. 2). The effects of the different microencapsulation methods on phage viability were tested by coculturing the microencapsulated phage cocktails with a mixed culture of Salmonella Typhimurium, Kentucky, and Enteriditis. Nontreated (naked) phage cocktails produced titers averaging 10.4 log10 PFU/mL. Microencapsulation with method one reduced the phage titer to 9.2 log10 PFU/mL, whereas microencapsulation with method two reduced the phage titer to 8.9 log10 PFU/mL (Table 2).
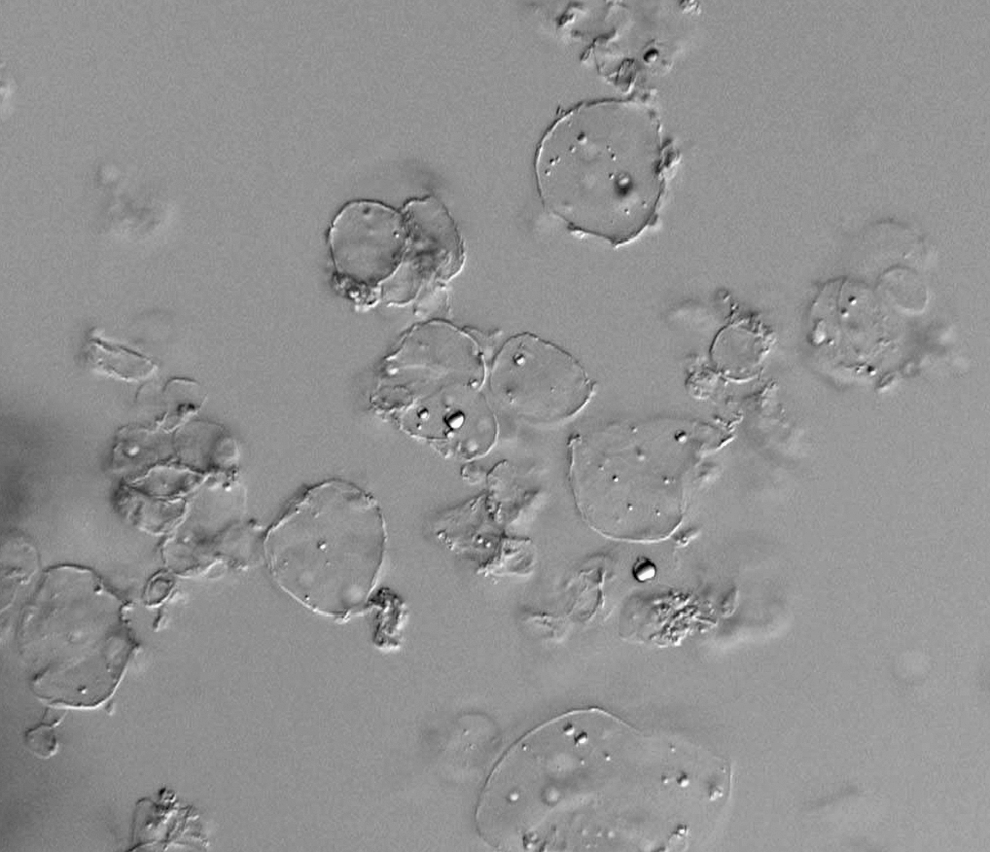
Images of microspheres. Phages were microencapsulated using method two and examined by light microscopy.
Phage viability (over time)
The effect of microencapsulation on phage viability over time was also tested. Each microencapsulated cocktail was incubated at 4°C and 22°C and phage concentrations were measured at 0 h, 24 h, 78 h, 7 days, and 14 days by standard plaque assay. There were no appreciable reductions in phage viability over time (up to 14 days) with either microencapsulation method regardless of incubation temperature. Phage cocktails microencapsulated with method one had starting concentrations of 9.2 log10 PFU/mL. The same cocktails had concentrations of 9.6 log10 PFU/mL (4°C) and 9.2 log10 PFU/mL (22°C) at 14 days. Phage cocktails microencapsulated with method two had starting concentrations of 8.9 log10 PFU/mL. The same cocktails had concentrations of 9.3 log10 PFU/mL (4°C) and 9.3 log10 PFU/mL (22°C) at 14 days (Table 3).
The 10 phages were combined into a cocktail (equal parts based on volume) and microencapsulated using method one or method two as described in the Materials and Methods section. Microencapsulated cocktails were incubated at 4°C and 22°C. Phage viability was measured over 14 days by plaque assay on a mixed culture of Salmonella enterica Typhimurium, Enteriditis, and Kentucky as described in the Materials and Methods section.
Discussion
There are close to 40,000 culture-confirmed cases of salmonellosis each year in the United States. No longer are human cases of salmonellosis linked exclusively to contaminated animal products as spinach, tomatoes, lettuce, peppers, and even more processed foods such as peanut butter have been implicated in large outbreaks (CDC, 2010). The ubiquity of Salmonella not only in livestock but also in the general environment indicates that multiple intervention strategies are needed to effectively reduce both contamination and infections.
Our previous studies demonstrated that oral administration of an anti-Salmonella phage cocktail could reduce preprocessing Salmonella shedding in swine (Wall et al., 2010). There remain a few challenges, however, to effectively transferring phage-based technologies to the livestock industry. Foremost is specificity. Many phages only infect certain bacterial species or serovars (Ackerman, 2005) and a large amount of diagnostic information is usually needed for narrow spectrum antibacterial therapies to be effective. With this in mind, we sought to create a broader spectrum phage cocktail. We chose S. enterica Typhimurium, Kentucky, and Enteriditis as these serovars are regularly among the most frequently isolated serovars involved in Salmonella infections in food animals and/or contamination of meat and poultry products (USDA, 2008). After extensive screening of wastewater samples, we produced a 10-phage cocktail that produced titers in excess of 10 log10 PFU/mL on a mixed culture of Salmonella Typhimurium, Enteriditis, and Kentucky.
In the United States, livestock producers often market hundreds and even thousands of animals at a time. Therefore, another challenge to implementing phage therapy in food animal production is the need to simultaneously and affordably treat multiple animals. If given as an oral treatment to limit bacterial colonization in the intestine, adequate numbers of phages need to by-pass the harsh environment of the stomach to reach the actual sites of infection. Microencapsulation is used with numerous oral biological treatments (Nechaeva, 2002; Ma et al., 2008) and we were previously successful in microencapsulating phage cocktails. In our previous live animal studies, microencapsulated phages were delivered to pigs by gavage and isolated in high concentrations from the ileum and cecum (Wall et al., 2010). Here we modified this protocol to eliminate some of the more expensive ingredients and found that the new method produced microspheres similar to the previous method. The microencapsulated phage cocktails remained viable for up to 14 days at room temperature. These results indicate that oral treatment of large numbers of animals through feed or water is possible. This is a focus of our current studies. Those experiments will also determine whether the new microencapsulation protocol is as successful at protecting the phage from the gastric environment of the stomach. The greater challenge in direct feeding the phages may be in assuring that each animal receives an adequate enough dose of the phage cocktail to significantly reduce or limit shedding.
It should be noted that contamination of carcasses is often attributed to contaminated hides (Bacon et al., 2000; Arthur et al., 2008). In some cases, animals that are not actively shedding Salmonella can enter the processor with Salmonella positive hides. This cross-contamination can, in turn, lead to carcass contamination. Therefore, future studies may want to focus on whether spraying animals with phage preparations before processing, perhaps in concert with oral administration, can further reduce end product contamination rates.
It will also be necessary to determine if phage therapy remains effective over longer periods. The development of resistance in the host could reduce efficacy; although in the field, phage resistance may be rarer than in the laboratory (Hurley et al., 2008; El-Shibiny et al., 2009). Likewise, phage populations are dependent upon host populations. Therefore, when host populations decrease, so can phage populations, which could lead to rebound infections that could be problematic for long-term use. While the phage cocktail described here has increased host range, there may be Salmonella serovars unaffected by the treatment, which could result in increased colonization by those serovars. Similarly, future live animal studies will want to examine whether phage treatment has any impact on the normal microbiota of the intestine. Each of these issues will have to be addressed to ensure that phage-based treatments are viable biocontrol measures over time.
Bacteriophages have great potential in numerous other areas outside of preharvest food safety. However, there is currently very little basic information regarding the actual viruses that have been used most effectively in different studies. Based upon the preliminary examination of morphology, the phages in our library belong to one of two families of tailed phages: Siphoviridae or Myoviridae (Ackerman, 2005). Our laboratory is currently focused on more thoroughly characterizing the different viruses in terms of genomics, growth kinetics, life cycles, and survival characteristics. This information should be useful in predicting where else these technologies could be used effectively and appropriately.
Footnotes
Acknowledgments
This study was supported in part by National Pork Board Grant No. 09-131 to P.D.E.
Disclosure Statement
No competing financial interests exist.