
Abstract
Finishing pigs infected with Salmonella pose significant food safety risks by carrying the pathogen into abattoirs. This study was conducted to determine the dynamics of Salmonella infection in finishing pigs, and associated immunological, physiological, and behavioral alterations, by longitudinally comparing infected to noninfected pigs during 6 weeks postinfection (p.i.). Bacteriological data revealed that all inoculated pigs started shedding Salmonella within 2 h p.i., and persistently shed the bacteria up to the end of the study. Ileal and cecal contents, as well as mesenteric lymph node samples, were all positive throughout the study, containing 3–4 log10 cfu/g of Salmonella at 24 h p.i., and 4–5 log10 cfu/g of Salmonella up to 4 weeks p.i. Levels of Salmonella dropped markedly (p < 0.05) in all samples at 5 weeks p.i. There was no difference between groups for blood cell counts. Tumor necrosis factor-α was greater (p < 0.05) in infected pigs: (1) in the mesenteric lymph nodes by 48 h p.i.; (2) at 24 h and 3 weeks p.i. in the ileum; and (3) in the cecum and spleen at 3 weeks p.i. Interleukin-12, interleukin-1 and its antagonist, and a porcine-specific antimicrobial peptide RNA expression in tissues changed over time, but were not different between groups. Infected pigs spent more time in ventral recumbency, standing, and sitting than controls (p < 0.01). Infected pigs were also more active (p < 0.01), and approached a novel object more quickly than control pigs (p < 0.05). No treatment differences were detected for rectal temperature or plasma cortisol (p > 0.10). This study shows that finishing pigs can carry high levels of Salmonella for up to 4 weeks p.i. in the gastrointestinal contents and mesenteric lymph nodes, shedding high levels of the bacteria without developing clinical symptoms, but developing an immune response throughout the intestinal tract. Moreover, subtle behavioral changes measured as postures were detected, and therefore warrant additional investigation.
Introduction
Until recently, research on the pathogenesis of Salmonella infection in pigs has been mostly focused in young animals. Researchers have striven to understand how the infection occurs, and how these animals become long-term carriers of the bacteria. Currently, most of the current knowledge is based on studies with young pigs, up to 6–8 weeks of age (Wood et al., 1989; Wood and Rose, 1992; Fedorka-Cray et al., 1994, 1995; Gray et al., 1995, 1996a, 1996b). However, more recently, increased interest has been focused on market weight pigs, with the goal of developing potential intervention strategies to be applied in this specific group of animals. A great deal of research has been conducted to develop knowledge on the preharvest ecology and epidemiology of Salmonella in swine production systems. However, a critical gap of knowledge exists in our understanding on the dynamics of Salmonella infection in finishing pigs, which limits the development of intervention strategies. Therefore, a longitudinal study was conducted with the objective of determining the dynamics of Salmonella enterica infection in finishing pigs, as well as to investigate potential immunological, physiological, and behavioral alterations associated with the infection.
Materials and Methods
Experimental animals and Salmonella challenge
This research was conducted under the approval of the Purdue University Animal Care and Use Committee. A total of 39 finishing pigs (barrows, 14–15 weeks of age) were used in this study. The animals were randomly divided into two groups (challenge or control) housed in individual pens (on concrete floor and with no direct contact with each other) located in separate facilities to avoid cross contamination, and allowed a 1-week adaptation period before starting the study. In the challenge group, 24 pigs (housed in the biosafety level 2 facility of the USDA, ARS, Livestock Behavior Research Unit, in West Lafayette, IN) were individually inoculated with a nalidixic acid-resistant S. enterica serovar Typhimurium (χ4232) strain. A frozen stock culture (maintained at −80°C in glycerol) was used to prepare the inoculum as described by Hurd et al. (2001). Each animal received 2 mL of 108 colony forming units (cfu) per mL of the challenge strain (intranasally; 1 mL in each nostril). Although the inoculation of individual animals does not precisely mimic natural infections, it is necessary to assure that all animals receive the same dose and at the same time. The control group (15 pigs) was not challenged and was maintained as the control for immunological, physiological, and behavioral analyses (until day 21 of the experiment). At days 1, 2, 7, 14, and 21 postinfection (p.i.), three randomly selected pigs per group were euthanized for necropsy and sample collection. Additionally, at days 28, 35, and 42 p.i., three randomly selected pigs from the challenge group were euthanized for necropsy and sample collection. Euthanasia was performed by injection with a solution of pentobarbital sodium and phenytoin sodium (Beuthanasia-D, Schering-Plough Animal Health Corp., Union, NJ), and immediately followed by exsanguination.
Bacteriological analysis
For the bacteriological analysis, individual fecal samples were collected directly from the rectum (using disposable surgical gloves, changed between animals to avoid cross contamination) at 2, 4, 6, 8, 24, and 48 h p.i. At necropsy on days 1, 2, 7, 14, 21, 28, 35, and 42 days p.i., additional samples were aseptically collected (from three pigs per sampling point), including mesenteric lymph node, ileal tissue and contents, cecal tissue and contents, and rectal contents (feces). Samples were immediately transported to the laboratory and processed within 10 min postcollection. In the laboratory, fecal and intestinal content samples were homogenized and 1 g aliquots (wet weight) were used for the isolation and enumeration procedures described below. Tissue samples were immersed in alcohol and flamed to eliminate any potential external contamination, and transferred into plastic bags, where they were smashed with a rubber mallet, and homogenized with a Stomacher 400 Circulator (Seward Limited, Worthing, United Kingdom). Aliquots from the homogenized tissue samples were used for the isolation and enumeration procedures described below.
Individual samples were sequentially enriched in Tetrathionate broth (1:10; incubated at 37°C for 24 h; Neogen Corporation, Lansing, MI) and Rappaport-Vassiliadis broth (Neogen Corporation) containing 50 μg/mL of novobiocin and 50 μg/mL of nalidixic acid (incubated at 42°C for 24 h; Sigma-Aldrich Corporation, St. Louis, MO), and streaked on XLT-4 agar (Neogen Corporation) containing nalidixic acid (50 μg/mL) for isolation of the challenge strain (incubated at 37°C for 24 h). Presumptive colonies on XLT-4 agar were individually picked and streaked on Rambach chromogenic agar (incubated at 37°C for 24 h; DRG International, Mountainside, NJ) for identification as Salmonella (Pignato et al., 1995). Additionally, individual samples were 10-fold serially diluted with buffered peptone water (Neogen Corporation), and directly plated (100 μL per plate) on XLT-4 agar containing 50 μg/mL of nalidixic acid (incubated at 37°C for 24 h) for specific enumeration of the challenge strain. Presumptive colonies were selected for identification as described above.
Immunological analysis
Liver, lung, spleen, mesenteric lymph node, jejunal, ileal, and cecal tissue samples were collected at 24 and 48 h p.i., and 1, 2 and 3 weeks p.i. to determine mRNA expression of two pathogen recognition molecules (toll-like receptors [TLRs] 2 and 4), two acute phase cytokines (tumor necrosis factor alpha [TNF-α], and interleukin 1 [IL-1]), IL-1 receptor antagonist (IL-1Ra), a T-cell activation cytokine (IL-12), and an antimicrobial peptide specific to pigs (PR-39). Tissue samples were rinsed with sterile 1 × Hank's Balanced Salt Solution (Invitrogen Corporation, Carlsbad, CA) and placed into 1 mL RNALater solution (Ambion, Austin, TX). Total RNA extraction was performed using QIAmp RNA Mini Kit (Qiagen, Valencia, CA). Extracted RNA was analyzed for quantity and purity using 260 and 280 nm spectrophotometer readings (GeneQuant pro; Biochrom, Cambridge, United Kingdom). Reverse transcription (cDNA synthesis) was carried out using TaqMan® Reverse Transcription Reagents (Product no. N8080234; Applied Biosystems, Foster City, CA). The reagent mixture contained 10 μL TaqMan RT buffer, 22 μL magnesium chloride, 20 μL dNTP, 5 μL random hexamers, 2.5 μL Multiscribe™ reverse transcriptase, and 2 μL RNase inhibitor. Exactly 38.5 μL of sample was added, and then centrifuged briefly to remove air bubbles. Tubes were transferred to a thermal cycler (PCRexpress, Serial no.13330; Hybaid, Basingstok, United Kingdom), and reacted for 60 min. On completion of reverse transcription, samples were stored at −80°C until further analysis. Primer and probe sequences for quantitative real-time polymerase chain reaction (qRT-PCR) were designed using Primer express 1.1 Software (Applied Biosystems) and synthesized by Applied Biosystems (Table 1). Probes were labeled with VIC fluorescent dye. A total of 5 μL of template cDNA was used for RT-PCR for each gene. Each sample was combined with 4.5 μL of forward and reverse primer, 3.25 μL probe, 7.75 μL DEPC water, and 25 μL Master Mix (TaqMan® Universal PCR Master Mix; Applied Biosystems) for the control genes. Bovine 18S ribosomal RNA was selected as a control gene and used for normalization purposes. All qRT-PCRs were performed in duplicate using a template from individual animals in each reaction. A relative standard curve was used as the qRT-PCR quantification method. The standard curve was constructed using the following dilutions of cDNA (in triplicate): 1, 0.5, 0.25, 0.125, 0.0625, and 0.0315. A single control sample was selected to be used as the template for the standard curve. qRT-PCR was performed and analyzed using ABI prism® 7000 sequence detection system (Applied Biosciences, Foster City, CA).
IL, interleukin; IL-1Ra, IL-1 receptor antagonist; TNF-α, tumor necrosis factor alpha; PR-39, an antimicrobial peptide specific to pigs.
Physiological analysis
The health of the animals is a critical component of their overall well-being. As such, infection may constitute an important cause of stress response, leading to physiological and behavioral alterations. With the objective of determining if the infectious challenge applied in this study could cause physiological alterations indicating a stress response, samples were collected for the analyses of stress mediators. Blood samples (10 mL in vacuum tube containing 7.5% K3EDTA) were collected via jugular venipuncture immediately before infection; 8 h, 16 h, 24 h, and 48 h p.i.; 1 week p.i.; and then weekly until 6 weeks p.i. Blood samples were immediately placed in an ice bath until they were centrifuged (650 g for 15 min) at 4°C within 4 h of sampling. Plasma from these samples was stored at −80°C until later analysis for plasma catecholamine and cortisol. For plasma catecholamine analysis, samples were acidified and deproteinized with 4 M perchloric acid, and a commercial alumina-based solid phase extraction kit (ESA, Chelmsford, MA) was used to extract catecholamines from the plasma. After extraction and injection onto the reverse-phase column, norepinephrine and epinephrine were detected using high-performance liquid chromatography with electrochemical detection. All samples were measured in duplicate, and assayed in 7 runs. Interassay coefficient of variation (CV) for norepinephrine, epinephrine, and dopamine were 5.43%, 5.10%, and 5.74%, respectively. Plasma cortisol concentrations (ng/mL) were determined for duplicate samples using standard RIA double-antibody kits (DiaSorin, Stillwater, MN). Samples were re-analyzed if the duplicates differed by more than 5%. These kits were previously validated for swine plasma (Daniel et al., 1999). Cortisol assays were performed in two batches and the intra-assay CV was 8.14% and the inter-assay CV was 10.48%. White blood cell number, differential white blood cell (neutrophils, lymphocytes, monocytes, eosinophils and basophils), red blood cells, hematocrit, hemoglobin, and platelet count were assessed using the Hemavet® HV950FS (Drew Scientific, Waterbury, CT). Rectal body temperature was measured using a standard human digital thermometer immediately before infection, and then every 8 h until 80 h p.i., for a total of 10 measurements p.i.
Behavioral analysis
Behavior data were collected using digital video recording systems installed on a hard drive connected to high resolution cameras. Both cameras and recording system were capable of capturing 480 frames per second. The activity and position of each pig was measured from the video recording beginning 12 h p.i. and continuing through 60 h p.i. Behavior data were obtained by conducting 10 min scan samples of videotaped behavior. The frequency of each position (ventral recumbency, lateral recumbency, sitting, or standing) and activity (inactive, eating, walking, nosing, or other) was measured in 4 h blocks of time, for a total of 12 observation periods. As a measure of sickness behavior, beginning 22 h preinfection, a novel object test was performed every 6 h for 4 days. Initially, the position and activity of each pig was recorded as described above. Then, each pig was tested individually for response to a novel object, such as bandanna, trash can, boot, knit hat, green rope, chain, plastic organizer, pry bar, tin pan, drainer, green mat, hammer, metal clipboard, flying disc, utility cord holder, and drink cup (items were randomized and never repeated). The designated object was attached to a hook and placed at the center of the front of the pen, and the length of time it took the pig to move to within 0.3 m of the object was recorded (latency to 0.3 m), as well as the time it took to touch the object (latency to touch). At that point the object was removed, disinfected in 10% bleach, and rinsed in water (to prevent cross-contamination) before use for the next pig. If the pig did not touch the object within 2 min, the test was ended. If the pig, during those 2 min, appeared to never notice the object (eyes were closed, or it was facing the other direction the entire test), it was noted and that measurement was removed from the analysis.
Statistical analysis
Data were checked for normality (Shapiro-Wilk test) and appropriately transformed when necessary. Bacteriological data (cfu/g of sample) were subjected to logarithmic transformation (log10 cfu/g of sample). Immunological data are presented as nontransformed values; however, all analyses were performed using the transformed data. Logarithmic transformations (using the Univariate Procedure of the statistical software) were used to normalize IL-1, IL-1Ra, IL-12, TNF, and PR-39 data for all tissues, and was only necessary to normalize TLR4 data for jejunal tissues. All other TLR data did not require transformation. Data were analyzed using SAS software (SAS Institute, Cary, NC) as repeated measures using PROC MIXED, with treatment (control or infected), sampling day, and interactions of treatment with day. Animal was considered the experimental unit. Data collected before infection were used as covariates. The most appropriate covariance structure for each model was determined using the Bayesian Information Criterion. Statistical differences are reported when p-values were <0.05. Tendencies toward significance (0.05 < p < 0.1) are also reported. Immunological data are presented as least squares mean ± standard error, whereas all other data are presented as arithmetic mean ± standard error. Means were separated using the Tukey-Kramer adjustment.
Results
Bacteriological analysis
Each animal was sampled (feces) twice before receiving the Salmonella challenge, and all were negative for the bacteria. All animals in the control group remained negative for Salmonella throughout the study period, demonstrating that no cross contamination occurred.
No inoculated animal showed clinical signs of infection, including no changes in body temperature. All inoculated pigs started shedding Salmonella within 2 h p.i., and persistently shed the bacteria up to the end of the study (i.e., for 6 weeks). The concentrations of Salmonella in the feces were high until week 4, and then reduced markedly (p < 0.05) in weeks 5 and 6 (Fig. 1). Following the same pattern, ileal and cecal contents, as well as mesenteric lymph node samples were all positive throughout the study, containing more than 3 log10 cfu/g of Salmonella at 24 h p.i., and 4–5 log10 cfu/g of Salmonella from 48 h to 4 weeks p.i. Concentrations of Salmonella dropped markedly (p < 0.05) in all samples at 5 weeks p.i., and were detectable only by enrichment (Fig. 1).
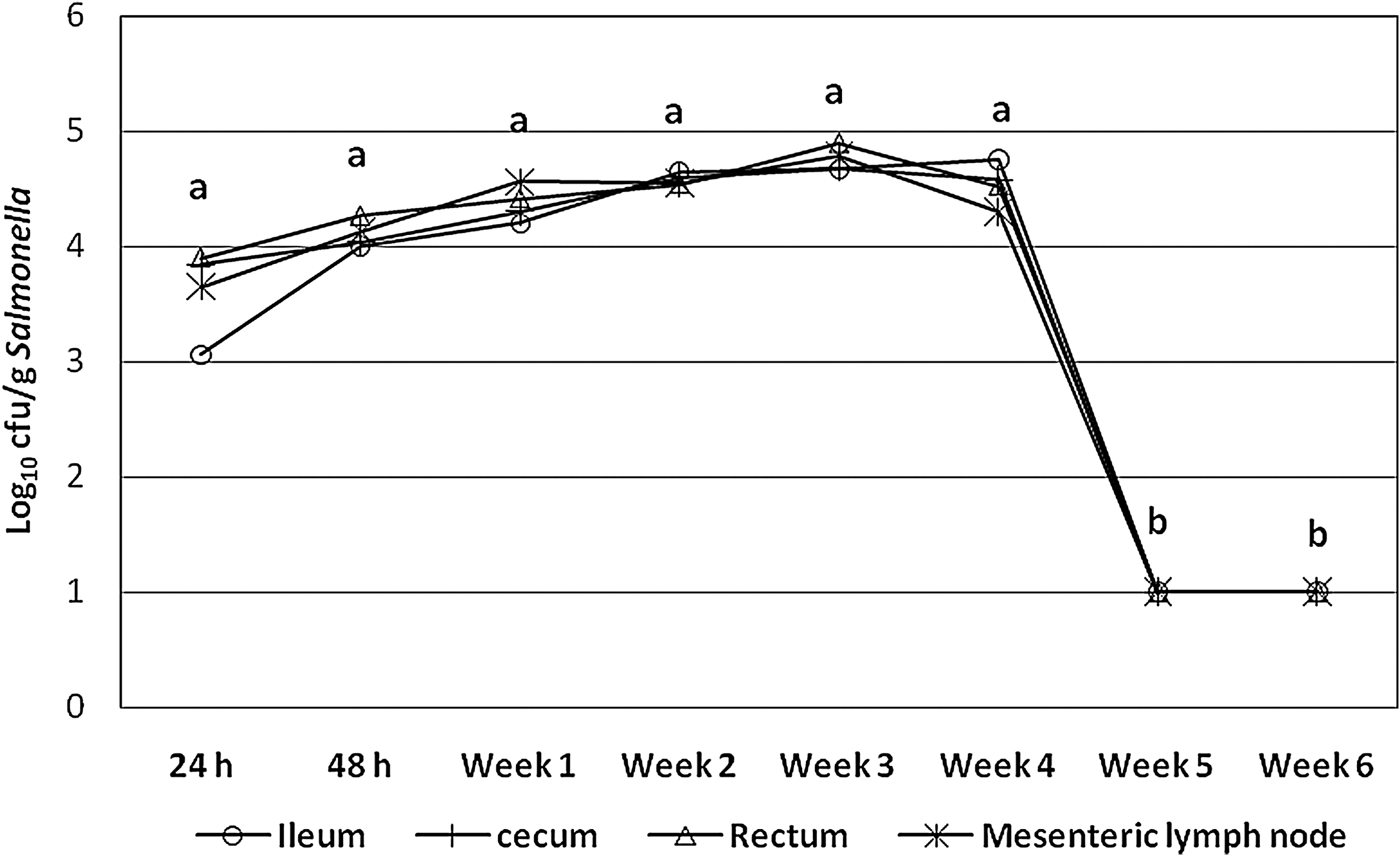
Concentrations of Salmonella enterica serovar Typhimurium nalidixic acid resistant (log10 cfu/g) in ileal and cecal contents, mesenteric lymph nodes, and feces of infected finishing pigs. Concentrations of Salmonella were higher than 3 log10 cfu/g and lower than 5 log10 cfu/g from 24 h to 4 weeks postinfection, dropping markedly (p < 0.05) in all samples at 5 weeks postinfection, becoming detectable only by enrichment. cfu, colony forming units. Different letters (a, b) indicate statistically significant difference (p < 0.05).
Immunological analysis
TLR2 expression was greater (p = 0.006) in spleen tissue of Salmonella infected pigs and tended to be greater (p = 0.06) in lung tissue of infected pigs (Fig. 2). In contrast, TLR4 expression tended (p = 0.06) to be less in cecum and lung tissues of infected pigs. IL-1 expression was not affected by Salmonella infection on any day. An effect of day was evident in the expression of IL-1 in cecum (p = 0.01) and in lung tissues (p = 0.02), with a peak seen on days 7 and 14 p.i., respectively. The expression of the IL-1Ra was greatest for the mesenteric lymph nodes on day 14 (p = 0.05) and the cecum on day 7 (p = 0.01) p.i. TNF-α was the only cytokine that showed significantly increased expression following the Salmonella challenge, with greater expression in the ileum (p = 0.007), mesenteric lymph nodes (p = 0.04), cecum (p = 0.04), and spleen (p = 0.03; Fig. 3). A weak treatment by day effect was found in spleen tissues such that by day 4 p.i., a trend (p = 0.10) for infected pigs to express more TNF-α was detected, and by day 5, the difference was statistically significant (p = 0.03). IL-12 expression was only different in the cecum. A trend (p = 0.10) for treatment by day was detected, such that infected pigs expressed more (p = 0.04) IL-12 on day 7, but less (p = 0.05) on day 15 p.i. than control pigs. Expression of the antimicrobial peptide, PR-39, exhibited an effect of day for cecum tissue (p = 0.01) and lung (p = 0.008) tissue, with a trend (p = 0.09) for a day effect in mesenteric lymph nodes. Blood cell counts were not statistically different between treatments (data not shown).
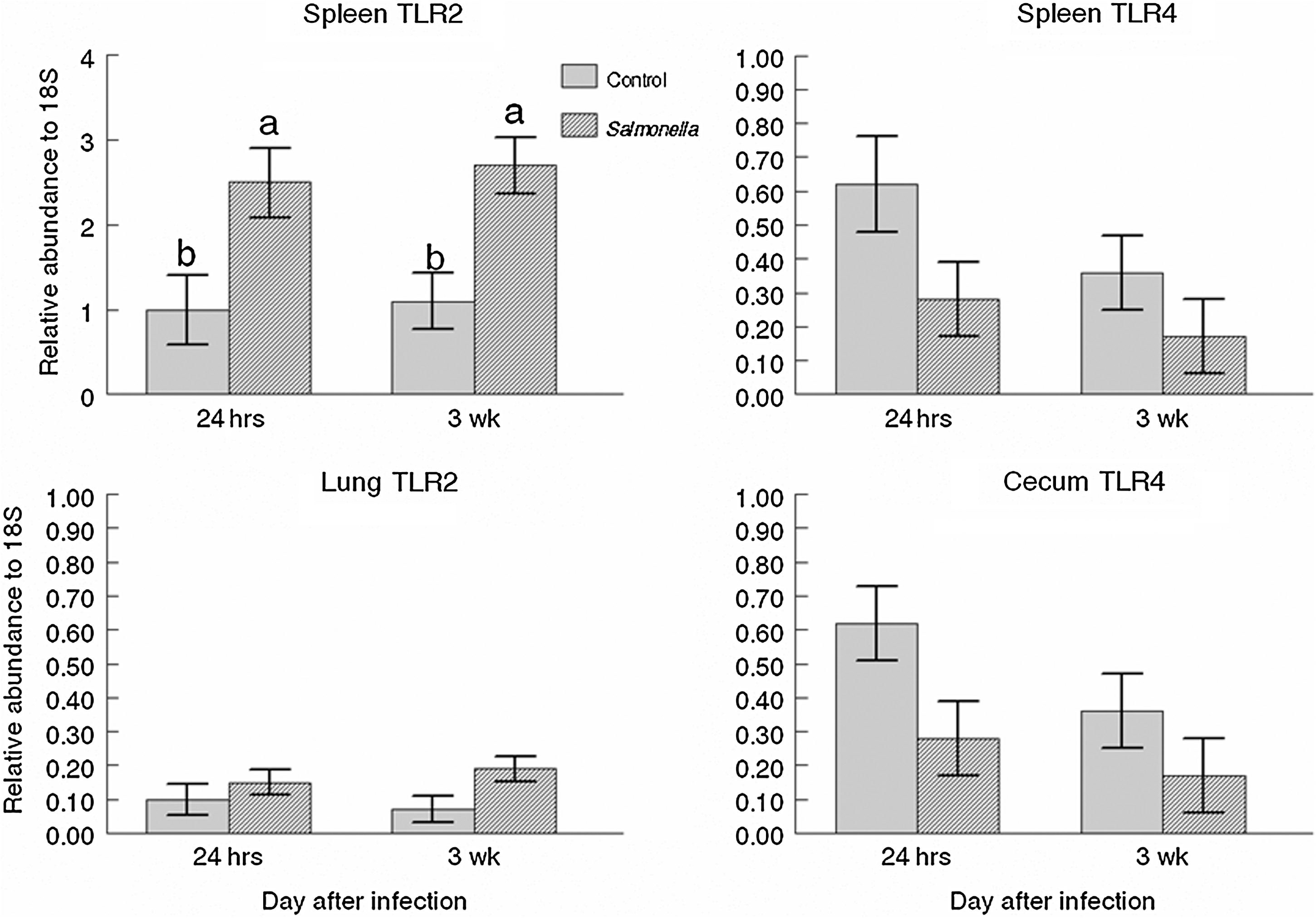
TLR expression in the cecum, lung, and spleen tissues (least square means ± standard error) collected from finishing pigs infected with S. enterica serovar Typhimurium or control (noninfected). Main effects of treatment were: spleen TLR2 (p = 0.006), and spleen and cecum TLR4 (p = 0.06), and lung TLR2 (p = 0.06). TLR, toll-like receptor. Different letters (a, b) indicate statistically significant difference (p < 0.05).
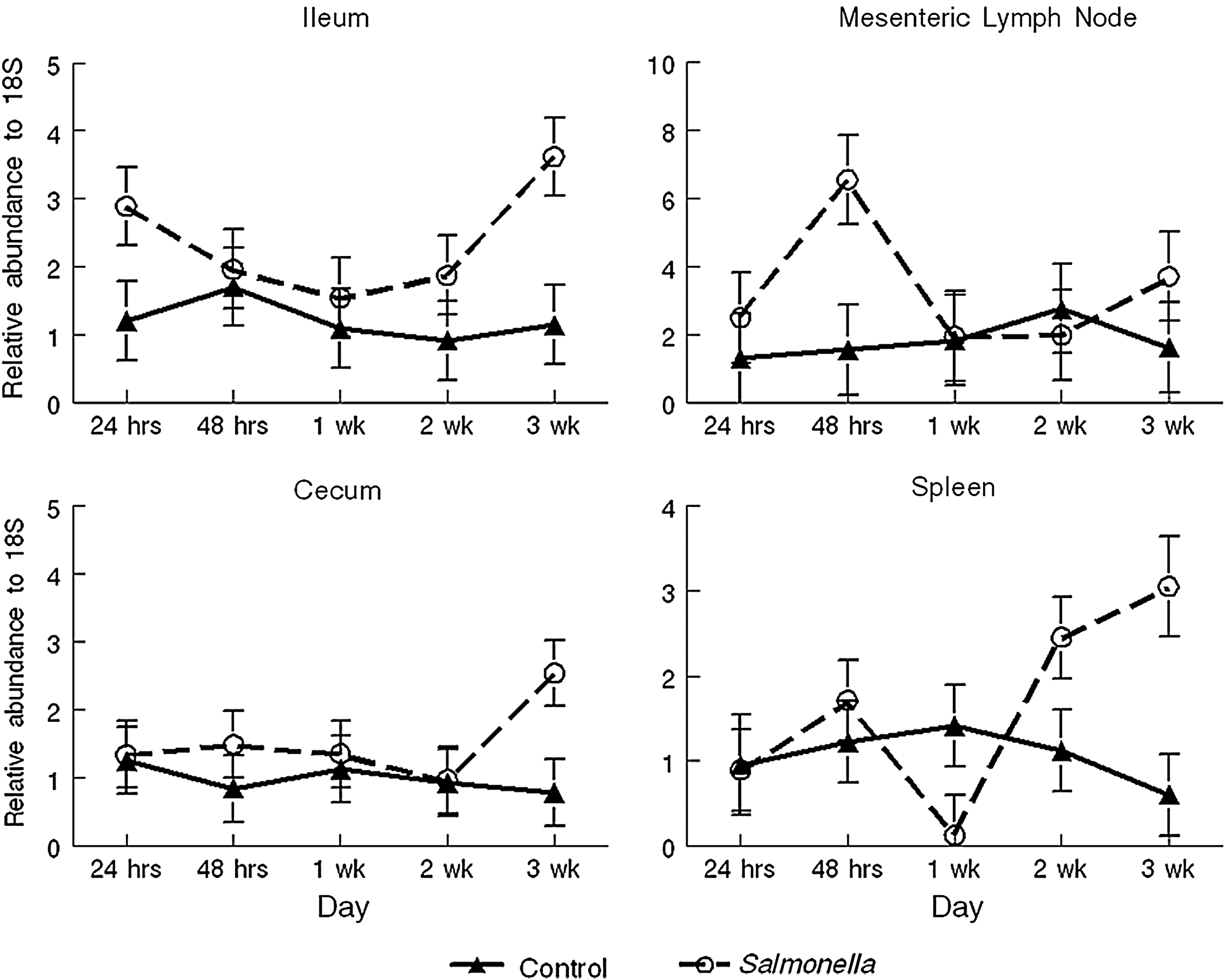
Tumor necrosis factor-α RNA expression for ileal, mesenteric lymph node, cecum, and spleen tissues (least square means ± standard error) collected from finishing pigs infected with S. enterica serovar Typhimurium or control (noninfected). Main effects of treatment: tumor necrosis factor expression in the ileum (p = 0.007), mesenteric lymph node (p = 0.04), cecum (p = 0.04), and spleen (p = 0.04).
Physiological analysis
No significant differences were detected in rectal temperature (39.3°C ± 0.03°C vs. 39.25°C ± 0.02°C), cortisol (20.83 ±1.18 ng/mL vs. 20.08 ± 0.95 ng/mL), epinephrine (1056 ± 108 pg/mL vs. 929 ± 65 pg/mL), norepinephrine (126 ± 14 pg/mL vs. 117 ± 10 pg/mL), or dopamine (167 ± 29 pg/mL vs. 145 ± 17 pg/mL) between control and infected pigs (respectively). Infection with Salmonella did not cause a febrile response (p = 0.29), with pigs experiencing a body temperature of 39.23°C ± 0.03°C preinfection, 39.47°C ± 0.04°C at 16 h p.i., and 39.22°C ± 0.02°C at 56 h p.i. Plasma cortisol stayed at a baseline concentration of ∼20 ng/mL for pigs in both treatments throughout the study. Epinephrine and dopamine decreased (p < 0.0001) over the study period with epinephrine concentrations starting at 140 pg/mL, and ending at 96 pg/mL. Dopamine concentrations started at 163 pg/mL, and ended at 86 pg/mL. Norepinephrine stayed at a steady concentration of ∼990 pg/mL.
Behavioral analysis
During the first 3 days p.i., infected pigs spent more time in ventral recumbency, and tended to stand more than control pigs (p < 0.006 and p < 0.06, respectively). Infected pigs also performed more nosing (p < 0.03), which was defined to include behaviors such as touching their nose to the feeder, water, pen, and floor without conducting any consumptive acts. Control pigs spent more time in lateral recumbency (p < 0.005). During the novel object tests, infected pigs approached objects more quickly than noninfected controls (p < 0.01; Fig. 4), but they did not differ in their latency to touch the object (p < 0.46).
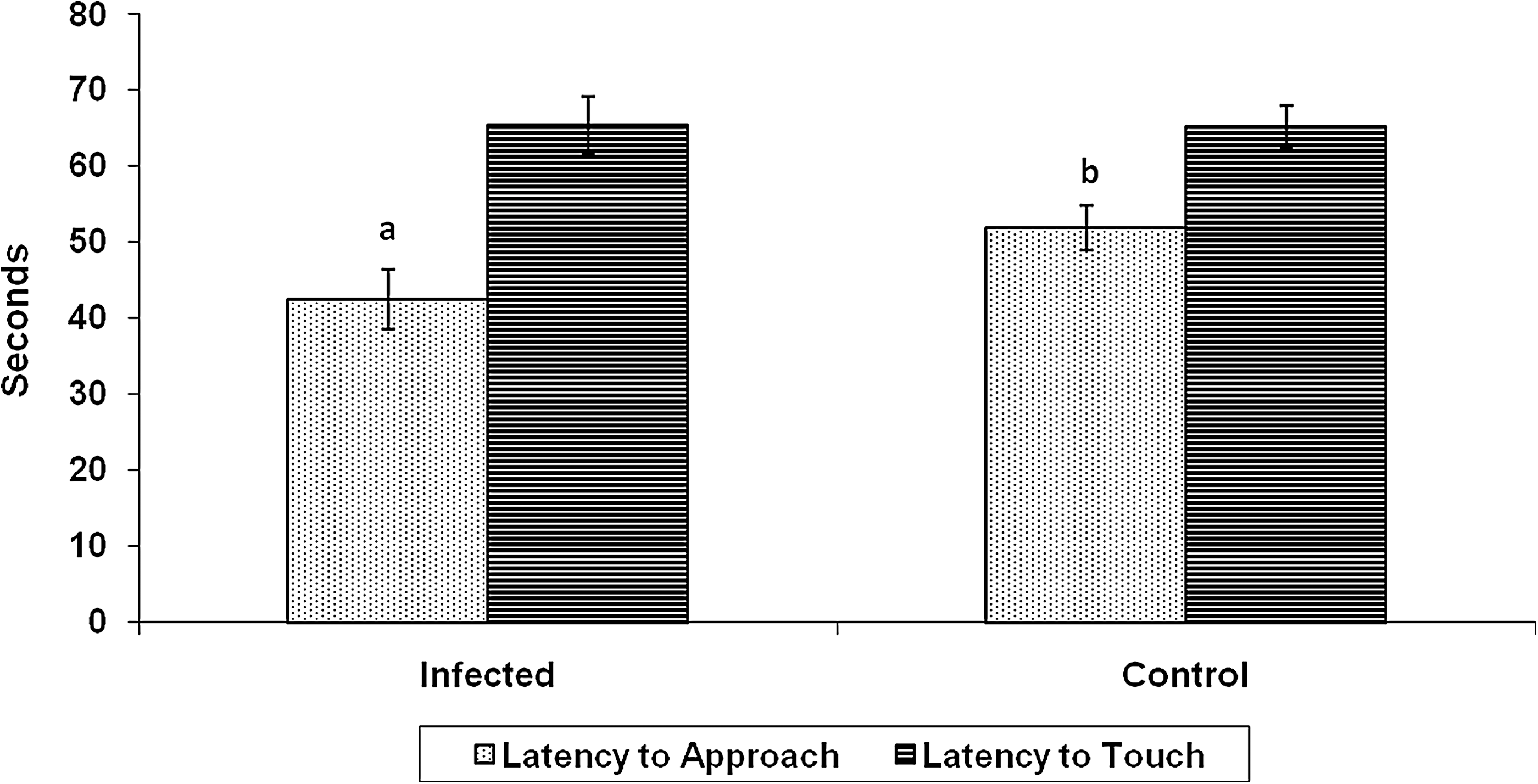
Mean response time to approach the novel object within 30.5 cm and touch the object, over 3 days p.i. with S. enterica serovar Typhimurium. Infected pigs approached the object quicker than noninfected controls (p < 0.01), but did not differ in their latency to touch the object (p < 0.46). Different letters (a, b) indicate statistically significant difference (p < 0.05).
Discussion
Finishing pigs infected with Salmonella can contaminate the lairage area at abattoirs, and be a source of meat contamination during slaughter and processing. However, little is known about the pathogenesis of the infection in animals of this age group. Most research on the pathogenesis of Salmonella infection in pigs has been conducted with young animals (Wood et al., 1989; Wood and Rose, 1992; Fedorka-Cray et al., 1994, 1995; Gray et al., 1995, 1996a, 1996b). This study demonstrates that Salmonella is able to rapidly spread through the gastrointestinal tract of finishing pigs, being shed in the feces within only 2 h p.i. This observation is in agreement with reports by Hurd et al. (2001) and Boughton et al. (2007). Also, Fedorka-Cray et al. (1995) reported that Salmonella could be recovered from the cecum of esophagotomized young pigs (6–8 weeks old) within 3 h of nasal inoculation, and suggested that the tonsils and lungs may be important sites for the rapid invasion and dissemination of the bacteria. However, to date, the mechanism(s) involved in the rapid dissemination of Salmonella in pigs is not known.
While infection with very large numbers of Salmonella may be required to initiate clinical disease (i.e., salmonellosis), Fedorka-Cray et al. (1994) demonstrated that pigs infected with only 104 colony-forming units of Salmonella Typhimurium develop a short-term carrier state, and Gray et al. (1995) demonstrated that a dose of 108 colony forming units of Salmonella Choleraesuis (a swine adapted or specific serovar) results in persistent infection for at least 12 weeks, but with intermittent shedding. It has been demonstrated that young pigs can shed Salmonella intermittently for several months after an experimentally induced infection with doses of 108 to 1010 colony forming units (Wilcock and Olander, 1978; Wood et al., 1989; Nielsen et al., 1995). Here, we demonstrate that finishing pigs (14-15 weeks of age) can excrete high concentrations of Salmonella in feces continuously for up to 4 weeks p.i. This observation is significant as it demonstrates that pigs infected during the finishing production stage (i.e., just few weeks before market) are capable of contaminating the facilities with high quantities of Salmonella, therefore favoring the transmission of the bacteria to other pigs within the same lot and, potentially, to subsequent lots housed in the same facility, if cleaning and disinfection are not properly conducted. Transmission studies exposing finishing pigs to Salmonella-contaminated environments have shown that low concentrations of the bacteria in the environment (102–103 cfu) are enough to infect the exposed animals (Hurd et al., 2001; Boughton et al., 2007). Based on recently published studies, it seems that both infectious dose, route of infection, and serovar are important determining factors for the amount and pattern (continuous or intermittent) of bacteria shedding (Gray et al., 1995; Osterberg and Wallgren, 2008; Osterberg et al., 2009). However, further research is required to more definitively demonstrate these effects.
Studies including the challenge of young pigs with Salmonella often report the occurrence of clinical symptoms (Wood et al., 1989; Wood and Rose, 1992; Fedorka-Cray et al., 1994; Casey et al., 2007; Scherer et al., 2008; Szabo et al., 2009), such as increased body temperature and diarrhea. In this study, no clinical symptoms of the infection were observed in older pigs (i.e., finishers), although the same strain have been used in previous studies (Wood et al., 1989; Fedorka-Cray et al., 1994). Unfortunately, feed consumption and growth performance were not monitored, and therefore, we were not able to determine these effects. This is an interesting area to be explored, particularly as a recently published study shows that inoculation of pigs with Salmonella Typhimurium or Choleraesuis promotes divergent responses in their somatotropic axis (Davis et al., 2010).
Several immune response changes were observed in the Salmonella-challenged pigs. TNF-α, IL-12, and TLR2 were all altered by the challenge. Curiously, no differences in RNA expression were found for IL-1 or PR-39. However, IL-18 predominates in respiratory and intestinal mucosa (Foss et al., 2001) and may be important in the pathogenesis of bacterial infections, including Salmonella.
The cecum appeared to be the quickest responder with enhanced IL-12 by day 7 p.i., which would be produced by macrophages and dendritic cells interacting with a pathogen. IL-12 would be the initiator of a more organized response and has synergistic effects with IL-2 and IL-18 (Pintaric, 2008). It stimulates growth and function of T-cells and is important in the activation of NK-cells, which are important innate immune responders in pigs.
Curiously, TNF did not increase in the cecum until day 21 p.i. TNF-α is a tightly regulated cytokine as is IL-1, needing multiple stimulants or greater concentrations or length of exposure, because it can be detrimental to the host as well as to the pathogen. TNF-α message in mesenteric lymph node increased only at day 2, and again at day 21 p.i. There are several possibilities for this phenomenon, including (1) the response to the challenge appears to be bi-phasic, with an initial quick response and a second one after Salmonella have multiplied and their own secondary infection ensues, and (2) the infection left the pigs susceptible to a secondary infection with other pathogen(s) in the environment. This is consistent with the prolonged detection of Salmonella in the ileal, cecal, and rectal contents, and mesenteric lymph nodes. Ileal tissue appears to have a more constant stimulation, which enhanced TNF-alpha throughout the challenge. Because of the altered expression, later, in the spleen, it is evident the Salmonella infection created a systemic response.
TLR2 increased in the spleen and tended to increase in lung at day 1, and again on day 21 p.i. However, TLR4 and IL-1, both of which typically increase in response to Gram-negative bacteria, such as Salmonella, did not change with the challenge in any of the tissues. TLRs and interferon-γ have a complex relationship with intracellular bacteria such as Salmonella (Wong et al., 2009). A main effect of day was detected for the expression of those genes and the anti-microbial peptide, PR-39. It appears that the response to this infection shifted toward an IL-12, interferon, and NK-cell-type response.
The behavioral measures collected in this study were used to determine to what extent Salmonella-infected pigs reacted to the infection, and to determine if differences in behavior could be used diagnostically. During acute illness, the general response of animals is to decrease activity, decrease feed intake, posture in protective positions, and increase rest (Hart, 1988). These changes in behavior are known to be controlled by cytokines, released from activated immune cells and centrally located glia cells (Dantzer, 2004). The cytokine response acts quickly, but persists in the hypothalamic-pituitary-adrenal system for up to 48 h after an immune challenge (Deak et al., 2005), and thus could have long-term effects in response to a Salmonella infection. As previously mentioned, pigs in this study did not exhibit any clinical signs due to the Salmonella challenge. Further, they did not show any physiologic changes consistent with activation of the stress response. Thus, they did exhibit subtle behavioral changes relative to postural movements and investigative behavior. The fact that infected pigs spent more time in ventral recumbency could indicate that they were trying to conserve body heat by lying on their feet and not exposing their underside, although we did not see an affect of infection on rectal temperature. Escobar et al. (2007) infected pigs with the porcine reproductive and respiratory syndrome (PRRS) virus and also found that infected pigs spent more time in ventral recumbency. Their results do differ from our in that their pigs also had a febrile response. Their pigs also showed a decrease in activity that was not shown in the current study. Possibly, infection with the PRRS virus induces a more pronounced feeling of malaise as compared to a bacterial infection as induced in this study. Indeed, using a model of sickness behavior through infection with the protozoa, Sarcocystis miescheriana, Reiner et al. (2009) also found decreases in activity; however, the clinical signs of infection were much more severe than those seen in our Salmonella challenge model. The greater time spent standing and investigating objects with their nose could indicate that the infected pigs were restless in response to the infection. In fact, the greater time spent in lateral recumbency by the control pigs is indicative of a relaxed posture. The infected pigs approached the novel object more quickly. This is the opposite response the authors expected, as we hypothesized that infection would cause the pigs to feel sick, and thus be less willing to express investigative behavior. The greater degree of investigative behavior in the novel object test may be consistent with a general increase in activity.
Conclusions
It is concluded that finishing pigs did not develop clinical symptoms after infection with high doses of S. enterica serovar Typhimurium, although they did exhibit subtle behavioral changes relative to postural movements and investigative behavior. Finishing pigs can carry high concentrations of Salmonella for up to 4 weeks p.i. in the gastrointestinal contents and mesenteric lymph nodes. Moreover, finishing pigs can shed high levels of Salmonella for up to 4 weeks p.i. contaminating the environment, and potentially transmitting the bacteria to other pigs.
Footnotes
Acknowledgments
The authors thank Gary Nowling, Kimberly McMunn, Rita Lockridge, Jessica Heinke, and Sophia Wilcox for excellent technical support.
Disclaimer
Mention of trade names or commercial products in this article is solely for the purpose of providing specific information and does not imply recommendation or endorsement of the U.S. Department of Agriculture.
Disclosure Statement
No competing financial interests exist.