
Abstract
During a Escherichia coli O157 prevalence study on cattle farms, 324 E. coli O157 isolates were collected from 68 out of 180 cattle farms. All isolates harbored the eaeA gene and the enterohemolysin (ehxA) gene. The majority of the strains only contained vtx2 (245 isolates), the combination of vtx1 and vtx2 was detected in 50 isolates, and in 29 isolates none of the vtx genes was present. Pulsed-field gel electrophoresis (PFGE) revealed that at a similarity level of 98% the isolates grouped into 83 different genotypes, 76 of which were only detected on one farm. Twenty-two out of the 68 positive farms harbored isolates belonging to more than one PFGE type, with a maximum of four different PFGE types. Minimal inhibitory concentrations of 10 antimicrobial agents were determined on a subset of 116 isolates, that is, one isolate per positive age category per farm. Acquired resistance to at least one antimicrobial agent was detected in 18 isolates and within a farm, only one resistance pattern was observed. All these 18 isolates were resistant toward streptomycin, and 16 of them also showed resistance toward sulfisoxazole. Six isolates were resistant to three or more antimicrobial agents.
Introduction
One of the most contentious areas in the management of E. coli O157 infections in humans lies in the possible effect that antimicrobials may have on the evolution of the infection. Because antimicrobials may cause increased expression of vtx in vivo (Zhang et al., 2000) and/or lysis of the bacterial cell walls, thereby liberating vtx (Walterspiel et al., 1992; Wong et al., 2000), they are not recommended for treating E. coli O157 infections in humans. However, there are studies suggesting that some antimicrobials, if administered early in the course of infection, may prevent disease progression to HUS (Fukushima et al., 1999; Ikeda et al., 1999). Although many E. coli O157 isolates are susceptible to several antimicrobials, recent reports indicate that, like in other foodborne pathogens, antimicrobial resistant strains of E. coli O157 emerge (Farina et al., 1996; Galland et al., 2001; Schroeder et al., 2002). Such strains can reach humans via the consumption of contaminated meat (Sørum and L'Abée-Lund, 2002) and may represent a threat to human health.
Genetic fingerprinting is an epidemiological tool that has often been used to identify probable sources of infections. Additionally, this approach has also been used to understand the ecology of E. coli O157 and might be used as a part of intervention strategies to reduce on-farm pathogens. From various methods available for genetic characterization of bacterial isolates, pulsed-field gel electrophoresis (PFGE) has been used successfully in the past for E. coli O157 and is considered as the gold standard to fingerprint E. coli O157 strains (Swaminathan et al., 2001).
Although PFGE is used worldwide to determine the genetic diversity of animal E. coli O157 strains, the number of isolates typed is often limited or they originated from a limited number of farms. The aim of the present study was to evaluate the genotypic diversity of E. coli O157 isolates within and between cattle farms (dairy, beef, and mixed farms). Additionally, the isolates were further characterized by their virulence genes profile and antimicrobial resistance.
Materials and Methods
Sample collection and microbiological analysis
E. coli O157 isolates (n = 324) were collected during a prevalence study on 180 cattle farms (dairy: n = 49, beef: n = 75, mixed: n = 45 and veal: n = 11) in the northern part of Belgium (Cobbaut et al., 2009). On each of these farms the bedding material in the pens was sampled by the overshoe method (Cobbaut et al., 2008). Within a farm, pens harboring the three following age categories were sampled if present: <8 months, 8–30 months, and >30 months.
E. coli O157 was isolated by enrichment in modified tryptone soya broth (Oxoid, Basingstoke, United Kingdom), supplemented with novobiocin (20 mg/L; Sigma-Aldrich, St. Louis, MO) during 6 hours at 42°C, followed by selective plating on sorbitol MacConky agar (Oxoid) supplemented with cefixime (0.05 mg/L; Invitrogen, Paisley, United Kingdom) and potassium tellurite (2.5 mg/L; Invitrogen) after immunomagnetic separation. After 24 hours of incubation at 42°C, up to three suspected sorbitol-negative colonies were picked from each selective plate and further tested for the presence of the O157 antigen using the E. coli O157 Latex kit from Oxoid. A maximum of nine agglutination-positive isolates per farm were selected (three of each sampled age category) for serotype confirmation by PCR (Cobbaut et al., 2009) and further characterization.
Identification of the virulence factor genes
For each isolate identified as E. coli O157, a multiplex PCR was performed using the primers for vtx1, eaeA, and ehxA as described by Fagan et al. (1999) and for vtx2 as described by Paton and Paton (1998). PCR assays were carried out in a 25 μL volume containing 1 μL of the lysates, 1× Taq buffer (20 mM Tris-HCl, pH 8.0, and 50 mM KCl), 0.75 U Taq DNA polymerase, 500 μM dNTPs, 3 mM of MgCl2, and 1.5 μM of the vtx1, eaeA, and ehxA and 0.6 μM of the vtx2 primers.
Temperature conditions consisted of an initial 95°C denaturation step for 3 minutes, followed by 30 cycles at 95°C for 20 seconds, 58°C for 40 seconds, and 72°C for 90 seconds. The last cycle was followed by an 8-minute elongation at 72°C and a final hold at 4°C. PCR amplification products were analyzed by electrophoresis in 1.5% agarose gels.
Pulsed-field gel electrophoresis
PFGE was performed to analyze the genetic relatedness of the studied isolates using the standardized method of the PulseNet (2007). The agarose-embedded DNA was digested in the agarose plugs with XbaI (Invitrogen). The resulting fragments were separated by a ChefMapper (Biorad, Marne La Coquette, France) in a 1% Seakem Gold agarose gel (Biowhittaker Molcular Applications, Rockland, ME). The electrophoresis conditions were as follows: auto algorithm; molecular weight: low, 300 K–high, 6000 K; calibration factor: 1.00; 0.5× TBE, 14°C, 1.0% SeaKem Gold agarose; gradient: 6 V/cm at 14°C; run time: 18 hours; included angle: 120°; initial switch time: 2.2 seconds; final switching time: 54.2 seconds; ramping factor: linear. After electrophoresis, the gel was stained with ethidium bromide and digitally captured under UV light.
PFGE profiles were clustered with GelCompar 3.0 (Applied Maths, Sint-Marten-Latem, Belgium) using the Dice coefficient (1% position tolerance) and the unweighted-pair group method using arithmetic averages algorithm. Each PFGE genotype (defined as a similarity of <98%) was assigned an arbitrary number.
Antimicrobial susceptibility testing
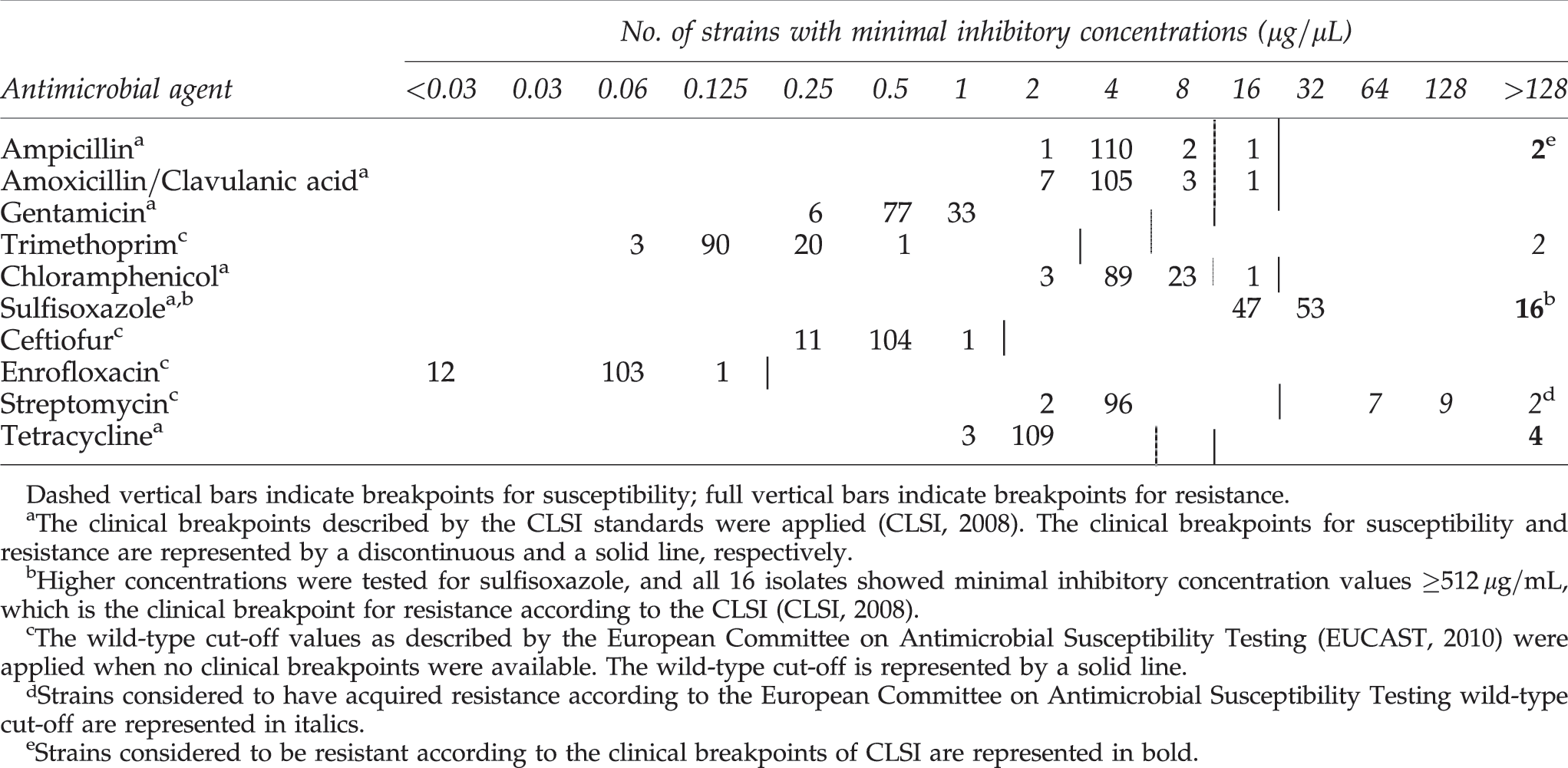
A collection of 116 isolates were analyzed for antimicrobial resistance. From each farm, one isolate from each positive age category was randomly selected. A quantitative agar dilution test on Mueller Hinton II agar (Becton Dickinson Company, Cockeysville, MD) was performed according to Clinical and Laboratory Standards Institute guidelines (CLSI, 2008). Minimal inhibitory concentrations (MIC) of ampicillin, amoxicillin/clavulanic acid, ceftiofur, chloramphenicol, enrofloxacin, gentamicin, streptomycin, sulfisoxazole, trimethoprim, and tetracycline were determined by incorporating twofold dilutions of these antimicrobials in the Mueller Hinton II agar plates. Concentrations ranged from 0.03 to 128 μg/mL except for sulfisoxazole, for which concentrations of 256, 512, and 1024 μg/mL were additionally tested. Overnight cultures were inoculated in Brain Heart Infusion Broth (Oxoid). Suspensions were aerobically incubated overnight at 35°C. Starting from the latter cultures, phosphate buffered saline (PBS) suspensions of 0.5 McFarland were made. Before inoculation on the antimicrobial-containing plates using the Denley Multipoint Inoculator (Mast, Bootle, United Kingdom), 1/10 dilutions in PBS were prepared. E. coli ATCC25922 and Staphylococcus aureus ATCC 29213 were used as control strains. MIC values were recorded after aerobic incubation at 35°C during 16–20 hours.
Results were interpreted by evaluating the distribution of the MIC values of a specific antimicrobial agent for all tested strains. In addition, the clinical breakpoints as recommended by the CLSI or, if not available, the wild-type cut-off values described by the European Committee on Antimicrobial Susceptibility Testing (EUCAST, 2010) were taken into account.
Results
As already mentioned in the article of Cobbaut et al. (2009), the 324 tested E. coli O157 isolates contained both the eaeA gene and the ehxA gene. Twenty-nine isolates, originating from eight farms, did not possess the vtx1 or the vtx2 genes. Of the 295 isolates that had at least one of the vtx genes, 245 had only vtx2 and 50 had both vtx1 and vtx2.
All isolates were typeable by PFGE and could be classified into 83 distinguishable PFGE types. The majority of the PFGE types (n = 76) were farm specific. Seven genotypes were detected on more than one farm. Table 1 shows the distribution of those PFGE types found on different farms. On farms with more than one PFGE type (n = 22), a maximum of four PFGE types were detected (Table 2). On half of these farms, one PFGE type predominated.
vtx gene present in isolates from farm 3.
vtx genes present in isolates from farm 9.
vtx gene present in isolates from farm 137.
vtx genes present in isolates from farm 146.
PFGE, pulsed-field gel electrophoresis.
Number of purchases within the year before the sampling day.
The vtx-negative isolates originating from eight farms belonged to four different PFGE types, and one type (P53) was shared by five farms (Table 1). In most cases, the isolates originating from one farm harbored the same type of vtx genes and had indistinguishable PFGE type. Nevertheless 17 farms had isolates with different PFGE types although they had the same vtx genes. On farm 9 one strain possessed the vtx1 and vtx2 genes, whereas the other strains had only vtx2, but they all belonged to the same PFGE type (P20).
The distribution of MIC values for the tested antimicrobial agents is presented in Table 3. From the 116 isolates tested for antimicrobial resistance, susceptibility to all the antimicrobial agents was observed in 98 (84.5%) of the E. coli O157 isolates. The remaining E. coli O157 isolates (n = 18), originating from 12 different cattle farms, were all resistant to streptomycin and 16 also showed resistance to sulfisoxazole (Table 4). Six isolates originating from three farms (21, 73, and 172) were multiresistant (=resistant to three or more antimicrobial agents): in addition to streptomycin and sulfisoxazole, resistance was observed to tetracycline (n = 4) and ampicillin and trimethoprim (n = 2). One streptomycin-resistant isolate should be considered as intermediately resistant to ampicillin, chloramphenicol, and the combination amoxicillin–clavulanic acid based on the CLSI breakpoints. Isolates showing antimicrobial resistance and originating from the same farm all had the same antimicrobial resistance profile (Table 4). On three farms (11, 80, and 95) both susceptible and resistant strains were isolated. Antimicrobial resistance was not detected in the vtx-negative isolates.
A: <8 months old; B: 8–30 months old; C: >30 months old.
Underlined = antimicrobial resistance tested.
Bold = resistant isolate.
Intermediate resistance for the antibiotics mentioned between parentheses.
St, streptomycin; Su, sulfisoxazole; Tet, tetracycline; Tri, trimethoprim; Amp, ampicillin; Chl, chloramphenicol; Amo/Cl, amoxicillin + clavulanic acid.
The 18 antimicrobial-resistant strains showed 15 distinguishable PFGE types. The sensitive and resistant strains originating from the same farm displayed discernable PFGE types (farm 11, 80, and 95). From farms 21, 68 and 174 strains were isolated belonging to at least two different PFGE types without difference in antimicrobial resistance pattern (Table 4). Two strains, each originating from a different farm (174 and 176), shared the same PFGE type (P9), antimicrobial resistance profile, and vtx genes.
Discussion
Identification of virulence factor genes demonstrated that 11.7% of the E. coli O157 isolates did not possess vtx genes, although the other virulence genes tested were present. The prevalence of vtx-negative E. coli O157 in cattle was similar to data from other studies. Chapman et al. (1993) found 7% vtx-negative E. coli O157 in fecal samples from cattle, and, more recently, 8.9% of the E. coli O157 were vtx-negative in a Danish study (Nielsen and Scheutz, 2002). It is unclear whether these strains are evolutionary precursors or mutations of the predominant vtx-positive E. coli O157 strains.
The presence of only one or a small number of different PFGE types on a farm is remarkable, since E. coli O157 can be introduced into cattle farms by multiple sources. If incoming cattle were the major source of E. coli O157, one might have expected much more diversity among E. coli O157 isolates. Possibly, certain E. coli O157 strain types are better adapted for survival and persistence in the environment. Likewise, E. coli O157 isolates of a single PFGE type have been observed to persist on farms from months to years (Shere et al., 1998; Rice et al., 1999; Renter et al., 2003; Cobbaut et al., 2008). Persistent infection and transmission between animals within a farm are an alternative hypothesis to explain strain type stability. The fact that on most farms only a single PFGE type of E. coli O157 could be isolated indicates that introduction and establishment of a new E. coli O157 strain on cattle farms is uncommon—either because an established farm strain is hard to replace by new strains or because new strains are rarely introduced on a farm.
A maximum of four different PFGE types were found in a minority of the farms. Higher diversity has been described in a study done in the United States, where up to seven different subtypes were found in dairy and feedlot cattle farms (Rice et al., 1999). In that study, open versus closed herd management practices had no effect on the number of subtypes present on dairy farms. Similarly, as no relationship was found between the diversity of E. coli O157 subtypes and the number of purchased animals in the present study, it can be suggested that avoiding the entrance of new animals will not prevent the introduction of E. coli O157 into the herd. Instead, nonbovine sources such as wild animals, flies, and human and vehicle movement may act as more important vehicles in the introduction of E. coli O157.
Although most PFGE types are specific to one farm, some strains appear to be more widespread, as indicated by the presence of seven indistinguishable subtypes of E. coli O157 on different farms. Type P53 was found on five farms of which three were in proximity. Two out of those farms belonged to the same owner. Farms located in proximity might share many common denominators, such as the bulk milk transport, human travel, natural water systems, feed sources, and nonbovine residing animals such as cats and dogs (Shere et al., 1998), insects (Hancock et al., 1998), and direct contact with wild animals (Wallace et al., 1997; Cízek et al., 1999; Sargeant et al., 1999; Renter et al., 2004).
In the present study, 15.5% of the E. coli O157 isolates showed resistance to at least one of the tested antimicrobial agents. Galland et al. (2001) found a third of the bovine E. coli O157 strains isolated in Kansas to be resistant to one or more antimicrobial agents, whereas Meng et al. (1998) reported an antimicrobial resistance prevalence of 24% for isolates of bovine origin. Antimicrobial resistance of E. coli O157 was most frequently detected to streptomycin (n = 18) and sulfisoxazole (n = 16), followed by tetracycline (n = 4). This is in accordance with other studies (Willshaw, 1996; Meng et al., 1998; Schroeder et al., 2002; Cho et al., 2006). Age has been described as a significant factor for gut colonization with antimicrobial-resistant E. coli, with a higher prevalence of resistance demonstrated in younger animals in surveys of cattle (DeFrancesco et al., 2004; Khachatryan et al., 2004). The data indicate that beside the age of the animals, also the PFGE, the vtx gene, and the farm type have no effect on the antimicrobial resistance of E. coli O157 strains.
Veterinary use of antimicrobial agents may favor spread of antimicrobial resistance in zoonotic bacteria and may pose a human health hazard (Mølbak, 2006). Resistant bacteria from food animals may enter the human population through the food chain, and when causing illness, antimicrobial treatment may not be successful. Although the role of early antimicrobial therapy in the prevention of HUS is still unclear (Griffin, 1995), surveillance of emerging antimicrobial resistance in enterohemorrhagic E. coli is an important tool for future disease management programs ensuring public health.
Conclusion
The present study demonstrated that all E. coli O157 strains isolated from cattle farms possess the eaeA and ehxA gene, but somelacked vtx genes. PFGE typing showed that in a large number of the farms, only one PFGE type was present. However, some PFGE types seem to be widespread. Antibiotic resistance in E. coli O157 is still low and mainly due to resistance to streptomycin and sulfisoxazole. Antibiotic resistance was not related to certain PFGE types, the age of animals, and the farm type.
Footnotes
Disclosure Statement
No competing financial interests exist.