
Abstract
Artificial digestion method is widely used for the detection of Trichinella larvae (mainly the mature larvae, e.g., encapsulated larvae in encapsulated Trichinella) in meat. The previous studies demonstrated that Trichinella spiralis pre-encapsulated larvae (PEL) at 14–18 days postinfection (dpi) had the infectivity to new hosts. However, to our knowledge, there is no report on the detection methods of PEL in meat. The purpose of this study was to compare the efficiency of artificial digestion and Baermann's methods for detection of T. spiralis PEL in meat, and to test the factors affecting the sensitivity of the two methods. Forty-five male Kunming mice were randomly divided into 3 groups (15 mice per group), and each group of mice was orally inoculated with 20, 10, or 5 muscle larvae of T. spiralis, respectively. All infected mice were slaughtered at 18 dpi, and the muscles were minced. The digestion method that was recommended by International Commission on Trichinellosis and Baermann's method were used to detect the PEL in the infected mice. The detection rate of PEL in both mice infected with 20 muscle larvae by digestion and Baermann's method was 100% (15/15); the detection rates of PEL in mice infected with 10 larvae by the two methods just mentioned were 93.33% (14/15) and 100% (15/15), respectively; when the mice infected with 5 larvae were tested, the different detection rate of PEL was achieved by using digestion method (63.33%) and Baermann's method (100%). Additionally, the number of PEL collected from the mice infected with 20, 10, or 5 larvae by Baermann's method was greater than that by digestion methods. The mortality of PEL increased along with the prolongation of digestion duration, because the PEL were not resistant to enzymatic digestion. The results revealed that the Baermann's method is superior to the digestion methods for detection of T. spiralis PEL in muscle samples with low-level infections.
Introduction
T
The effective detection of Trichinella larvae in meat is essential for ensuring food safety, and facilitating international trades and regulations. The detection methods include direct (trichinelloscopy and artificial digestion) and indirect (serological) techniques. The trichinelloscopy is a simple method that can be used in any laboratory or field setting. It is, however, a laborious and time-consuming method for inspecting the individual carcasses of food animals. Its sensitivity is lower than that of the artificial digestion method for detecting Trichinella larvae in meat from food animals (Forbes et al., 2003; Beck et al., 2005). Artificial digestion method is a sensitive, efficient, reliable, and cost-effective method. Consequently, it has become the method of choice for routine slaughter inspection in most countries and is recommended for the control of Trichinella in domestic and wild animals intended for human consumption by the International Commission on Trichinellosis (ICT), World Organization for Animal Health (OIE), and the European Union (Gamble et al., 2000; Webster et al., 2006; OIE, 2008).
The theoretical sensitivity of digestion method testing a 1 g sample is 1 larva per gram (lpg) of muscles, the minimum level of infection considered to be of public health concern. However, due to nonuniform larval distribution and technical limitations, the actual sensitivity of this method is 3–5 lpg when examining the prescribed 1 g of meat (Gamble, 1998; Forbes and Gajadhar, 1999; Li et al., 2008). When 10 samples of 100 g of minced pork containing 3–50 larvae were tested by digestion method in 24 laboratories, 37.5% of laboratories failed to detect some positive samples with the percentage of false negatives ranging from 11% to 100% (Marucci et al., 2009). This level of sensitivity could result in false negative results in samples containing sufficient larvae to cause human Trichinella infection. In addition, it is not know whether the digestion method can be used to detect the infective pre-encapsulated larvae (PEL) in meat.
The disadvantage of serological methods (such as enzyme-linked immunosorbent assay [ELISA] using excretory–secretory [ES] antigens) for detecting of Trichinella infection is that they give false negative results during the early stage of infection. In lightly or moderately infected pigs, a 4–7 week time lag occurs between the time when larvae become infective for a new host and positive serology (Gamble et al., 1996; Gamble and Patrascu, 2004; Nockler et al., 2005). This delay in the production of antibodies to Trichinella indicates that animals infected with lower numbers of Trichinella larvae may not be detected for several weeks after exposure, even after some muscle larvae have become infective (Kapel and Gamble, 2000; Wang et al., 2007; Gajadhar et al., 2009). The previous studies demonstrated that the PEL of T. spiralis at 14–18 days postinfection (dpi) had the infectivity to new hosts (Campbell, 1983; Li et al., 2009). Consequently, this window of false negativity of serologic tests prevents their use for the early detection of Trichinella infection.
The aim of this study was to compare the efficiency of artificial digestion and Baermann's methods for detection of the PEL of T. spiralis in muscles of experimentally infected mice with low-level infections, and test the factors affecting the sensitivity of artificial digestion and Baermann's methods.
Materials and Methods
T. spiralis and experimental animals
T. spiralis isolate (ISS 534) used in this study was obtained from domestic pigs in Nanyang city in Henan Province of China, and maintained by serial passage in male Sprague-Dawley rats at 6- to 8-month intervals. For inoculation of the mice used in the study, we used T. spiralis muscle larvae recovered from the infected rats at 42 dpi by artificial digestion in a solution of 0.33% pepsin (1:31,000 U.S. National Standard Formulary strength; Sigma)—1% HCl at 43°C for 4 hours, as described next (Gamble et al., 2000).
Six-week-old, specific pathogen-free male Kunming mice weighing 20–25 g each were used in the study. The mice were purchased from the Experimental Animal Center of Henan Province and were bred in plastic microisolator cages. Forty-five mice were randomly divided into three groups of 15 mice each. Each group was orally inoculated with 20, 10, or 5 muscle larvae of T. spiralis, respectively (Cui et al., 2008). The infected mice were killed at 18 dpi by exposure to ether and cervical dislocation, and their carcasses were then flayed and eviscerated. The muscles of mice infected with 20, 10, and 5 larvae were tested by both artificial digestion and Baermann's methods, as described next. The muscles of additional fifteen mice infected with 300 larvae were used to test the factors affecting the sensitivity of artificial digestion and Baermann's methods for detecting PEL.
Artificial digestion method
Artificial digestion method for detection of T. spiralis larvae was performed by using the magnetic stirring method recommended by ICT (Gamble et al., 2000). Briefly, the infected mouse carcasses were trimmed free from all fat and fascia, and then blended for several times with 5 second intervals until no visible pieces of muscles remains. Two grams of muscle samples mixed with 60 mL of digestion solution (0.33% pepsin [1:31 000 U.S. National Standard Formulary strength; Sigma]-1.0% HCl) at a 1:30 ratio were digested for 4–5 hours at 43°C. A magnetic spin bar was utilized to provide continual mixing during digestion. At the conclusion of the digestion process, the entire digests from the beaker are poured through an 80 sieve (180 μm mesh) into a 2 L separatory funnel, and settled for 30 minutes. The final 10 mL of fluid recovered from separatory funnels was examined under an Olympus microscope with 40×magnification, and larvae in the fluid were counted.
Baermann's method
The Trichinella larvae in muscle samples minced were directly collected by Baermann's method (John and Petri, 2006) with some modification. Briefly, a 2-L glass separatory funnel with a diameter of 12 cm is set up in a ring stand, with a piece of rubber tubing connected with its stem and pinchcock closing the tube. A 1 mm mesh sieve with a diameter of 9 cm was covered with 4–6 layers of cotton gauze. The funnel was filled with prewarm (37°C) saline solution to a level just covering the gauze, and 2 g of muscle samples minced was placed at the center of the gauze. Prewarm (37°C) saline solution was again added into the funnel, and the samples were covered. All air that may remain inside the funnel and tube should be eliminated. The apparatus was left on a stand at room temperature, but the rubber tubing was submerged in a constant water bath at 37°C. After 0.5–3.5 hours, 40 mL of fluid from the stem of the funnel was drawn off through the tubing into a 50-mL glass centrifuge tube, and settled for another 20 minutes, after which all but 10 mL of the fluid at the bottom of the tube was aspirated out, the 10 mL of fluid remaining in the tube was examined under an Olympus microscope with 40×magnification, and larvae in the fluid were counted.
Comparison of Baermann's and digestion methods for detection of PEL in muscles of mice infected with low doses of T. spiralis
One hundred twenty muscle samples of mice experimentally infected with 20, 10, or 5 larvae of T. spiralis on 18 dpi were used to compare the sensitivity of artificial digestion and Baermann's methods for detection of PEL. The muscles of mice infected with 20 and 10 larvae were equally divided into thirty 2 g samples, and the muscles of mice infected with 5 larvae were equally divided into sixty 2 g samples. All of the infected muscle samples were tested by both artificial digestion and Baermann's methods.
Effects of digestion duration on the viability of PEL
To test the effects of digestion duration on the viability of PEL, 5 mice infected with 300 larvae were sacrificed on 18 dpi; their muscles were minced, mixed, and equally divided into five samples (9.6 g each). The five muscle samples were digested for 1, 2, 3, 4, and 5 hours, respectively (Gamble et al., 2000). The PEL were collected and enumerated. The larval viability was identified by the morphology, activity, and methylene blueeosin-borax staining for 1 minute (Shen et al., 2003). The live larvae are active in wriggling motion and colorless, the dead larvae are not active and stained as reddish or purple.
Effects of separation time and gauze layers on recovery of PEL by Baermann's method
To determine the effects of separation time (the time when apparatus stands for) and the layers of gauze on the recovery of PEL, nine mixed muscle samples (9.6 g each) from mice infected with 300 larvae on 18 dpi were tested by Baermann's method. The recovery of PEL in each sample was evaluated by using four, five, and six layers of gauze at each 0.5 hour interval during 0–3.5 hours of separation. Three replications were done of each procedure. The PEL in each sample at each 0.5 hour interval were collected and enumerated.
Statistical analysis
All statistical analyses of data were done with SPSS for Windows version 16.0 (SPSS Inc.). The number of PEL recovered in different groups was compared by Wilcoxon rank sum test and Kruskal-Wallis H test. To compare the detection rates of PEL by digestion and Baermann's methods, Chi-square test was used. The Spearman rank correlation was used to find out correlation between the mortality of PEL and the digestion duration. The effect of digestion duration on the viability of PEL was tested by linear trend. The level of significance used was 5% (p<0.05).
Results
The sensitivity of digestion and Baermann's methods for the detection of PEL in mice infected with 20, 10, or 5 larvae of T. spiralis at 18 dpi are shown in Table 1. The detection rates of PEL in mice infected with 20 larvae were 100% by the two methods just mentioned. When the mice infected with 10 larvae were examined by digestion and Baermann's methods, the detection rate of PEL was 93.33% and 100%, respectively (χ 2=1.034, p>0.05). When the mice infected with five larvae were tested, the detection rate of PEL was achieved by using digestion method (63.33%) and Baermann's method (100%) (χ 2=13.469, p<0.05). Additionally, all the number of PEL collected from mice infected with 20, 10, or 5 larvae by Baermann's method was greater than that by digestion methods (Z 20=−4.669, Z 10=−4.674, Z 5=−5.266, p<0.05).
PEL, pre-encapsulated larvae.
The effects of digestion duration on the viability of PEL were shown in Table 2 and Figure 1. Some of the PEL were died and broken after digestion for 4 hours (Fig. 1C). It was difficult to recover the dead larvae when the digestion mixtures passed through the sieve and settled. There was a significant positive correlation between the mortality of PEL and the digestion duration (r=0.528, p<0.05). The mortality of PEL showed an increasing trend with the prolongation of digestion time (χ 2=920.772, p<0.05).
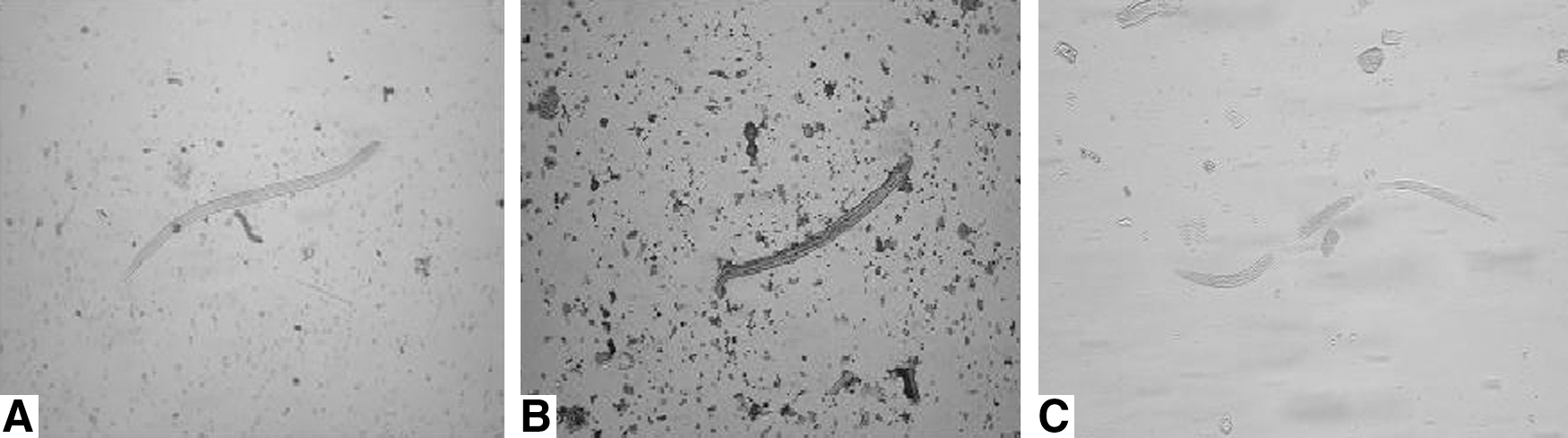
The viability of Trichinella spiralis pre-encapsulated larvae (PEL) at 18 days postinfection (dpi) after digestion for 3 hours
The effects of separation time and the layers of gauze used in Baermann's method on the recovery of PEL at 18 dpi were shown in Figure 2. In this study, the differences among the tests with different layers of gauze were not significant at different separation time (χ 2=0.022, p>0.05). However, the solution after filtration using four layers of gauze was turbid, and the larvae were not easily observed, because the sediment contained muscle fibers. When six layers of gauze were used, the solution after filtration was clear, the larvae could be easily observed and enumerated. The number of PEL recovered by Baermann's method showed a decreasing trend with a prolongation of separation time. The number of PEL recovered during the first time range (0–0.5 hour) was higher than that during the second time range (0.5–1 hour) (Z=−1.933, p<0.05). The PEL recovered during 0–2.5 hours account for the 95% of total PEL collected during 0–3.5 hours.
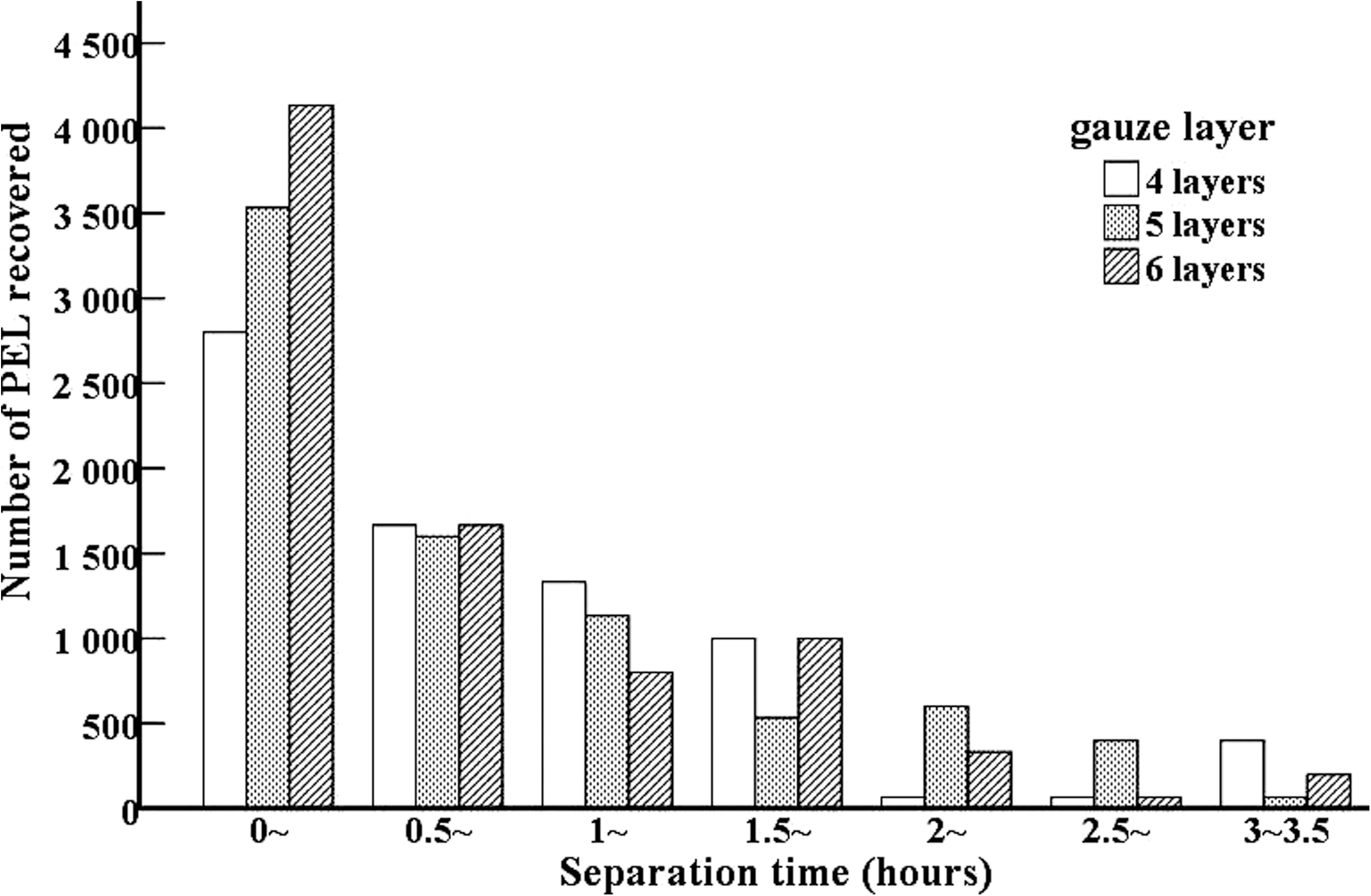
Effects of separation time and gauze layers on recovery of T. spiralis PEL at 18 dpi by Baermann's method.
Discussion
Trichinellosis not only is a public health hazard by affecting patients but also represents an economic problem in porcine animal production and food safety. Due to the predominant zoonotic importance of infection, the main efforts in many countries have focused on the control of Trichinella or the elimination of Trichinella from the food chain. There are increasing global requirements for reliable diagnostic and control methods for Trichinella in food animals to ensure meat safety and to facilitate international trade (Gottstein et al., 2009).
Artificial digestion method is the method of choice for routine slaughter inspection recommended by ICT, and has been widely used in many countries. The principle of digestion method is the digestion of muscle tissue with an acidified pepsin solution to release the live Trichinella larvae from muscle and the direct microscopy of the recovered larvae, but it is mainly applied to the examination of the mature (e.g., the encapsulated) larvae in muscles. To assure the integrity of the test for digestion, digestion duration for 4–5 hours was necessary. However, in this study, we found that there was a significant positive correlation between the mortality of PEL and digestion duration, and the mortality of PEL was increased along with prolongation of digestion duration and the PEL are not resistant to enzymatic digestion for 4–5 hours. The death of PEL after enzymatic digestion might be related with the absence of complete developed cuticles before encapsulation. It was difficult to recover the dead larvae when the digestion mixtures passed through the sieve and settled, which can yield false-negative results (Li et al., 2010). The results indicated that the PEL were not resistant to enzymatic digestion, and the digestion method was unsuitable to detect T. spiralis PEL in meat with low-level infection. Some of the outbreaks of human trichinellosis are probably caused by the infective PEL in inspected meats except that meat samples containing low numbers of encapsulated larvae were not detected, which is enough to cause clinical trichinellosis (Djordjevic et al., 2005; Teunis et al., 2011).
The Baermann's method is a classical parasitological technique mainly used to separate nematode larvae from fecal and soil material. The method is based on the active migration or movement of larvae. Feces or soil samples are suspended in water with several layers of cotton gauze, and larvae will move into the warm water (37°C) after passing through the gauze. After settling, the larvae are collected for identification. The PEL of T. spiralis also have this characteristic of migration in warm water (Ko and Wong, 1992; Zhang et al., 1998). In this study, we found that the PEL after filtration with 6 layers of gauze could be easily observed and the PEL recovered during 0–2.5 hour account for the 95% of total PEL collected during 0–3.5 hours. Hence, the optimization of Baermann's method for collecting PEL is the combination of using six layers of gauze and separating for 2.5 hours. The muscle samples containing PEL at 18 dpi were tested by digestion and Baermann's methods. When the muscle of mice infected with five larvae were tested, the detection rate of PEL by Baermann's method (100%) was obviously higher than that by digestion method (63.33%). Additionally, the number of PEL collected from muscle samples of mice infected with 20, 10, or 5 larvae by Baermann's method was also higher than that by digestion method. The results revealed that the Baermann's method is superior to the digestion method for detection of T. spiralis PEL in meat with low-level infection (five larvae). To ensure the elimination of infective Trichinella PEL from animal meat and food safety, it is suggested that when one pig was Trichinella-positive on routine slaughter inspection, all pigs in the farm tested negative with digestion methods should be re-tested with Baermann's method. However, further validation of Baermann's method is required with the use of larger samples of infected muscles from species such as pigs that are routinely tested for T. spiralis for the purpose of food safety.
Footnotes
Acknowledgments
This work was supported by the National Basic Research Program of China (No.2010CB530000), the National Natural Science Foundation of China (No. 30972492, 30972579).
Disclosure Statement
No competing financial interests exist.