
Abstract
Salmonella is a major cause of foodborne diseases worldwide, which has fueled the demand for the development and evaluation of sensitive, specific, and rapid detection methodologies, such as polymerase chain reaction (PCR). In this study, six primer pairs for the detection of Salmonella were evaluated by PCR with isolates of Salmonella spp. (115) and other bacteria (104). The primers designed for the sifB gene provided the best performance regarding specificity and sensitivity (100%). These primers were selected and used to develop a PCR assay for Salmonella detection during the enrichment steps of the conventional detection method in spiked beef samples. The enrichment steps were: buffered peptone water (BPW), Rappaport-Vassiliadis soya broth (RVS) and at the Müller-Kauffmann tetrathionate novobiocin broth (MKTTn), after 18 h (BPW) and 24 h (RVS and MKTTn) of incubation. The initial concentrations of the Salmonella inocula were 101, 102, and 103 colony-forming units/25 g. The protocol was able to detect Salmonella at all concentrations in the enrichment steps, but not in the nonenriched samples. These results indicated that the proposed protocol was suitable to detect Salmonella in beef during the intermediate stages of the conventional isolation protocol, substantially reducing the time required to obtain the final results.
Introduction
S
Conventional methods for Salmonella spp. detection, such as ISO 6579, consist of the steps of pre-enrichment, selective enrichment, selective plating, biochemical tests, and seroagglutination and usually take several days (ISO, 2002). Alternative methods for detection, such as the polymerase chain reaction (PCR), have been developed to reduce the analysis time, obtain higher sensitivity and specificity, improve the flow of sample handling, and enable the automation of laboratory protocols (Jasson et al., 2010; Mandal et al., 2011). Despite being more specific and sensitive, PCR, like any other technique, has its limitations: the components of the food can interfere with the amplification reaction, and the concentration of the microorganism of interest may be below the detection limit of the technique, leading to false-negative results (Wang et al., 1997). Furthermore, the method does not distinguish between live and dead cells. Various procedures, such as the homogenization of samples and DNA extraction, may be used to minimize these problems (Kanki et al., 2009; Li and Mustapha, 2002).
Several protocols for detecting Salmonella in foods include the enrichment of samples prior to PCR (Wang et al., 1997; Gouws et al., 1998; Vázquez-Novelle et al., 2005; Myint et al., 2006; Saroj et al., 2008; Matias et al., 2010). The use of an enrichment step prior to PCR allows the proliferation of microorganisms that were present in low concentrations, increasing the sensitivity of the technique.
Although there are numerous laboratory protocols for the detection of Salmonella spp. in foods products, their detection limits and efficiency vary widely. Furthermore, many of them do not use an internal amplification control (IAC) and therefore provide no guarantee that the negative results are not caused by inhibition of the PCR. For these reasons and the importance of this pathogen, the development and improvement of detection methods are still necessary. This study aimed to evaluate and compare PCR protocols based on six pairs of primers for the detection of Salmonella spp. and to test the most effective primers in assays with artificially contaminated beef.
Materials and Methods
Reference bacterial strains, field isolates, and DNA extraction
Reference strains and field isolates of Salmonella and other bacteria were used in this study (Table 1). The reference strains were obtained from American Type Culture Collection (ATCC, Manassas, VA), Bio Manguinhos Fiocruz (BM, Rio de Janeiro, RJ, Brazil), and National Collection of Type Cultures (NCTC, Salisbury, UK). The field isolates were obtained from Brazilian research labs at Federal University of Viçosa (Viçosa, MG), Federal University of Paraná (Palotina, PR), São Paulo State University (Botucatu, SP) and University of São Paulo (São Paulo, SP). These isolates originated from foods of different animal origin that had been submitted for Salmonella detection using the conventional method and were serotyped at the National Reference Center of Bacterial Intestinal Infections—FIOCRUZ (Manguinhos, Rio de Janeiro, Brazil). The non-Salmonella strains were selected due to their phylogenetic proximity with the genus Salmonella or for being found in the same environment as this pathogen.
FI, field isolate; ATTC, American Type Culture Collection (Manassas, VA); BM, Bio Manguinhos (FIOCRUZ, Rio de Janeiro, RJ, Brazil); NCTC, National Collection of Type Cultures (Salisbury, UK).
All strains were maintained in tryptone soya broth (TSB, Oxoid Ltd., Basingstoke, England) added to glycerol at 20% (vol/vol) and −20°C. Before use, the strains were streaked onto tryptone soya agar (TSA, Oxoid) and incubated at 37°C for 24 h, after which isolated colonies were selected and transferred to TSB and incubated at 37°C for 24 h. The resultant cultures were used for DNA extraction and further analysis. DNA was extracted using the Wizard® genomic DNA purification kit (Promega Corp., Madison, WI); after extraction, the DNA was subjected to electrophoresis on 1% agarose gels that were subsequently stained with GelRed™ nucleic acid gel stain (Biotium Inc., Hayward, CA) and observed using a transilluminator.
Primers
The primers used were previously published and are commonly used in various studies for detecting Salmonella, with the exception of the pair designed to detect the sifB gene. These primers were designed using DNAman software (version 4.0; Lynnon Corp., Pointe-Claire, Quebec, Canada) on the sequence of the sifB gene (GenBank: AF236076.1), which is an effective gene in the Salmonella pathogenicity island 2 (SPI-2) that is related to a type III secretion system gene (Miao and Miller, 2000; Freeman et al., 2003). The primer sequences employed in the present study, the amplification target regions, the size of the products, and the references are shown in Table 2.
Efficiency of Salmonella detection using the selected primers
To assess the efficiency of the primers for the detection of Salmonella by PCR, DNA samples of all strains listed in Table 1 were used. Each PCR reaction contained 2 μL of the DNA sample (200 ng/μL), 12.5 μL of GoTaq® Green Master Mix (Promega), 10 mM of each primer, and nuclease-free water in sufficient quantity to bring the final volume to 25 μL. The PCR amplification conditions are shown in Table 3. The products of amplification were subjected to electrophoresis on 2% agarose gels and subsequently stained and observed as described above.
To evaluate the efficiency of the PCR protocols, the sensitivity, specificity, and positive and negative predictive values of each primer pair were calculated (Malorny et al., 2003a).
Sequencing of the PCR products
The PCR products obtained using the primers designed for sifB were purified and sequenced by Macrogen Inc. (Seoul, South Korea). The results were analyzed using the Sequencher™ software (version 4.1.4, Gene Codes Corp., Ann Arbor, MI) and BLAST (
Beef samples and inoculation of Salmonella
Beef cuts (tenderloin) from an inspected slaughterhouse were purchased in local markets. The superficial portions of the cut were aseptically discarded to minimize possible contamination. For analysis, four 25-g units were taken from each cut for analysis and transferred to sterile bags. In each bag, the beef portions were inoculated with 1-mL aliquots of Salmonella at approximate concentrations of 101, 102, and 103 colony-forming units (CFU)/mL (treatments 1, 2, and 3, respectively). For a negative control, the beef portion in the fourth bag was inoculated with 1 mL of sterile solution of 0.85% NaCl (wt/vol) (treatment N). After inoculation, the samples were manually homogenized and incubated at 7°C for 30 min to allow attachment of the bacteria; a similar condition was described by Brichta-Harhay et al. (2007).
Salmonella Typhimurium ATCC 14028 was used for inocula preparation, after incubation in TSB at 37°C for 24 h, dilution in TSB to a turbidity similar to McFarland scale tube 1, and 10-fold dilution in 0.85% NaCl (wt/vol). To confirm the inoculated concentrations, the cultures were pour-plated on TSA (Oxoid) in duplicate, and following incubation at 37°C for 24 h, the colonies were counted (final results are expressed as CFU per 25 g of inoculated beef). The procedure of inoculating beef samples with Salmonella was repeated six times.
Conventional detection of Salmonella in artificially contaminated beef samples
Detection of Salmonella in artificially contaminated beef samples was conducted according to the ISO 6579:2002 method (ISO, 2002). An aliquot of 225 mL of buffered peptone water (BPW) was added to each sample, which was incubated at 37°C for 18 h. Then 0.1-mL aliquots were transferred to Rappaport-Vassiliadis soya broth (RVS) and 1-mL aliquots to Müller-Kauffmann tetrathionate novobiocin broth (MKTTn), and they were incubated at 41.5°C and 37°C, respectively, for 24 h. Aliquots of each broth were streaked on xylose lysine deoxycholate agar (XLD) for incubation at 37°C for 24 h and subsequent observation of typical colonies of Salmonella. All of the culture media were purchased from Oxoid.
Detection of Salmonella by PCR in artificially contaminated beef samples
Aliquots of 1 mL were obtained from the following enrichment cultures of the conventional method for Salmonella detection: BPW before incubation, BPW after incubation, RVS after incubation, and MKTTn after incubation. DNA from the aliquots was extracted as described above and amplified by PCR.
PCR was performed using the extracted DNA, the pairs of primers for detection of sifB gene, and primers U1/U2 in the same reaction. The primers U1 (CAGCMGCCGCGGTAATWC) and U2 (CCGTCAATTCMTTTRAGTTT), which are based on a conserved region of the 16S rRNA, were used as the IAC to produce an amplicon of 408 base pairs (bp) (Lane et al., 1985; Border et al., 1990). The amplification conditions were the same as those used previously for the primers SIFBF/SIFBR (Table 3), except that the annealing temperature was changed to 49°C. The products of amplification were subjected to electrophoresis on 2% agarose gels, subsequently stained, and observed as described above.
Results and Discussion
Evaluation of primers for the detection of Salmonella by PCR
In the first stage, some of the amplification conditions were modified from the original protocols to optimize the PCR reactions, allowing the amplification of the appropriate target regions and minimizing the production of nonspecific bands (Table 3). The six primer pairs provided positive results for all of the Salmonella isolates tested. In other studies, PCR assays using primer pair 139/141 were not able to detect some strains of Salmonella Litchfield, Salmonella Senftenberg, Salmonella Pullorum, Salmonella Saintpaul, and Salmonella enterica subsp. arizonae (Rahn et al., 1992; Malorny et al., 2003a; Moganedi et al., 2007). Malorny et al. (2003a) failed to detect the S. Enteritidis and S. bongori strains using the primer pair S18/S19 and failed to detect one strain of Salmonella enterica subsp. arizonae using the primer pair ST11/ST15. In the present study, however, these serovars were not tested, with the exception of Salmonella Enteritidis.
Non-Salmonella isolates were tested for the occurrence of nonspecific amplifications. The primer pairs OMPCF/OMPCR, S18/S19, 139/141, LHNS-531/RHNS-682, and ST11/ST15 produced nonspecific bands from various isolates of Citrobacter freundii, including PCR products with sizes similar to the target region, which were considered false-positive results. In addition, the S18/S19 primers produced false-positive results for Klebsiella spp.
Gooding and Choudary (1999) observed PCR products amplified from Citrobacter freundii strains with the expected size for those of Salmonella spp. using the primers ST11/ST15, and similar results for Pseudomonas aeruginosa and Enterobacter cloacae strains using the primer pair S18/S19. Multiple nonspecific bands were observed when these primer pairs were used with other strains of bacteria, such as Hafnia alvei, Shigella sonnei, and Yersinia enterocolitica. Malorny et al. (2003a) reported similar results, observing nonspecific bands for Citrobacter spp., Klebsiella oxycotan and Yersinia enterocolitica, when the pair ST11/ST15 was used, and nonspecific bands for Proteus vulgaris, Hafnia alvei and other bacteria when the primer pair S18/S19 was used.
In this study, the primer pair 139/141 produced nonspecific bands for isolates of Campylobacter coli, Raoutella planticula, Y. enterocolitica, Proteus mirabilis, and Listeria innocua, but they had a higher molecular weight and were distinguishable from the expected 284-bp band. Other studies using these same primers reported similar results. Rahn et al. (1992) observed nonspecific bands from different non-Salmonella strains, including Citrobacter freundii, P. mirabilis, Klebsiella pneumoniae, Hafnia alvei, Pseudomonas sp., Escherichia coli, and Enterobacter aerogenes. Moganedi et al. (2007) obtained nonspecific bands from isolates of Citrobacter freundii, Klebsiella pneumoniae, and Pseudomonas aeruginosa. Löfström et al. (2004) observed similar results with C. freundii, E. coli, E. hermanii, Kluyvera ascorbata, Providencia rettgeri, and Serratia rubidea isolates.
The sensitivity, specificity, positive predictive value, and negative predictive value of each primer pair are shown in Table 4. Because all of the Salmonella isolates were correctly identified by PCR with all of the tested primers, the sensitivities and negative predictive values were 100%. The primer pair SIFBF/SIFBR was the only one that did not produce nonspecific bands or false-positive results, and therefore it had the highest specificity and positive predictive value. The sequences of the PCR products of primers SIFBF/SIFBR confirmed that the target region of the sifB gene of Salmonella spp. was properly amplified. Therefore, this primer pair exhibited the best performance at this stage of the study.
The sifB gene encodes the protein SifB, which is an effector translocated by the type 3 secretion system of SPI-2 (Miao and Miller, 2000). SifB locates to the Salmonella-containing vacuole and Salmonella-induced filaments, both related to intracellular replication of the pathogen (Freeman et al., 2003). Studies indicate that sifB is not required for SIF formation, but deletion of this gene causes a significant reduction in Salmonella replication in macrophages (Freeman et al., 2003; Buckner et al., 2011). Further studies are necessary to fully elucidate the functions and impact of sifB on the virulence of this pathogen (Figueira and Holden, 2012). There are no previous reports of sifB being used in methods for detection of Salmonella, and it was selected in this study for its potential role in Salmonella infections.
Salmonella detection by PCR in enriched cultures of artificially contaminated beef
The frequencies of detection of Salmonella by the conventional method and by PCR using the artificially contaminated beef are shown in Table 5. An agarose gel containing the PCR products of a sample is shown in Figure 1. Nonspecific bands were not observed when the primers SIFBF/SIFBR were used for the detection of pure cultures in the first stage of this study, but when these primers were combined with the IAC primers for PCR with artificially contaminated beef samples, faint nonspecific bands over 800 bp in size were observed in some reaction products. The negative controls, for which no Salmonella was added to the meat, led to negative results for both the conventional detection by the ISO 6579:2002 method and by PCR, confirming the previous absence of Salmonella spp. in the portions of beef used in the inoculation assay.
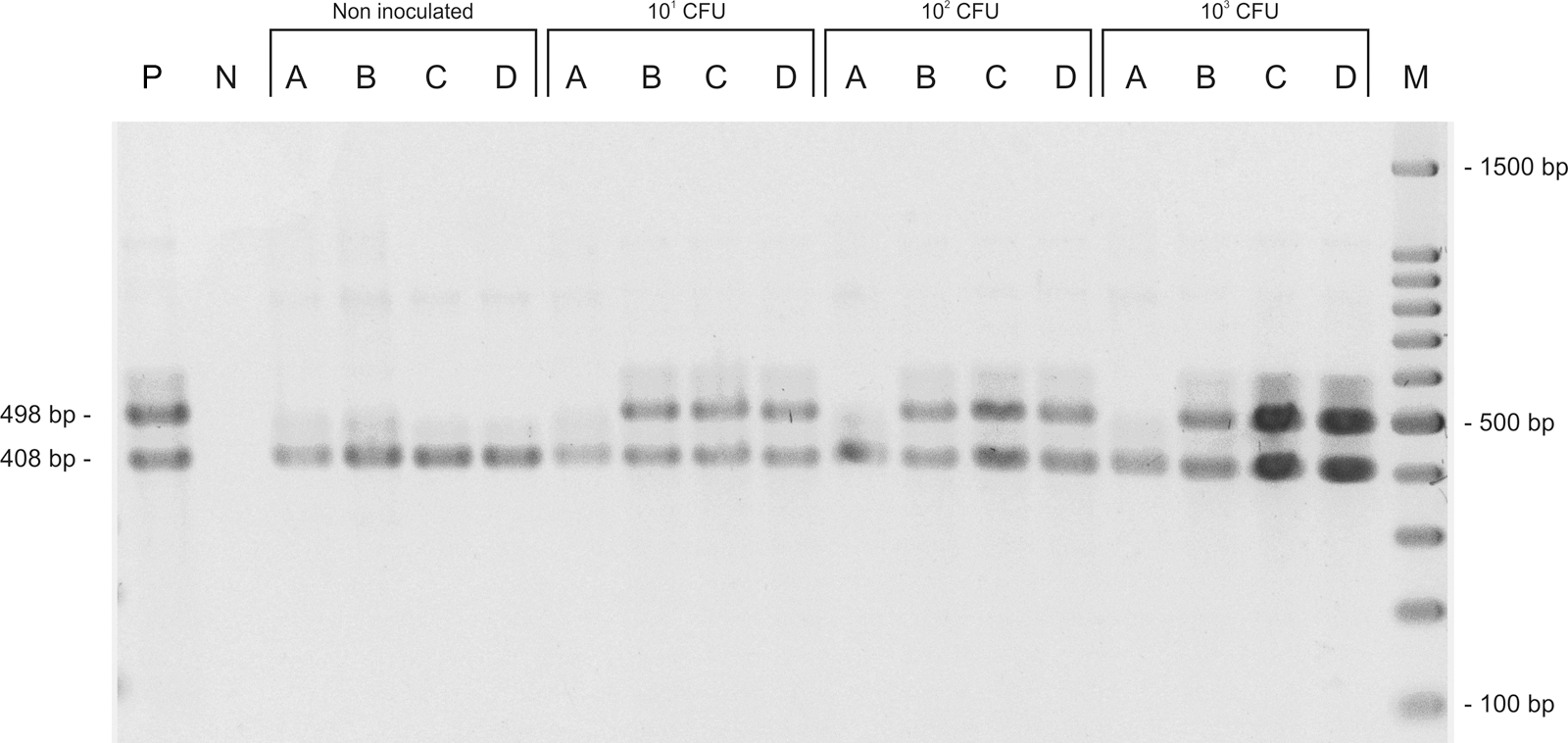
Agarose gel electrophoresis of polymerase chain reaction (PCR) products obtained from PCR reactions using SIFBF and SIFBR primers for Salmonella detection in beef samples artificially contaminated with Salmonella Typhimurium at concentrations 101, 102, and 103 colony-forming units (CFU) per 25 g, and noninoculated. Amplification products of SIFBF/SIFBR (498 base pairs [bp]) and U1/U2 (408 bp, used as internal amplification control). P, Salmonella Typhimurium ATCC 14028 (positive control); N, water (negative control); A, buffered peptone water before incubation; B, BPW after incubation; C, Rappaport-Vassiliadis Soya Broth; D, Müller-Kauffmann Tetrathionate Novobiocin Broth; M, 100-bp molecular weight marker.
NC, negative control (sterile saline); T1, 101 colony-forming units (CFU)/mL; T2, 102 CFU/mL; T3, 103 CFU/mL.
Mean and standard deviation (except for NC) in log CFU/mL of inoculated Salmonella sp.
BPW-0, Buffered Peptone Water prior to incubation; BPW-18, BPW after incubation for 18 h.
RVS, Rappaport-Vassiliadis soya broth; MKTTn, Müller-Kauffmann tetrathionate novobiocin broth.
All of the PCR products amplified using the U1/U2 primers contained a 408-bp band. These primers were used as the IAC, as proposed by Border et al. (1990). The use of an IAC eliminated the possibility that negative results were due to inhibition of the amplification reaction (Hoorfar et al., 2003). Among the causes of inhibition in PCR reactions are substances present in the media that was used in the enrichment steps (Rossen et al., 1992). Hyeon et al. (2010) evaluated the inhibitory effects of BPW, RVS, and MKTTn in a real-time PCR assay for the detection of Salmonella: The results of the addition of BPW to the PCR were similar to those of the control, whereas adding RVS and MKTTn had considerably higher inhibitory effects. In the presence of high concentrations of MKTTn, Salmonella spp. could not be detected. These findings are similar to those of Stone et al. (1994), who failed to detect Salmonella spp. when using Rappaport-Vassiliadis (RV) and tetrathionate (TT) broths for the enrichment of samples prior to PCR. The RVS and MKTTn broths are modifications of RV and TT, respectively, with essentially the same composition as the original medium. The inhibitory effect found in these studies may be due to the substances that make these media selective, such as bile salts and magnesium chloride (Rossen et al., 1992; Stone et al., 1994; Hyeon et al., 2010). In the present study, however, there was no evidence of inhibition of amplification caused by the enrichment media.
In the present study, Salmonella was detected in samples produced with all of the inoculated concentrations, including 101 CFU/25 g, by the conventional method (ISO, 2002) and the PCR technique (Table 5). Using PCR, however, detection occurred only after incubation in enrichment media and was not possible in samples diluted in BPW but not incubated. These results were consistent with those of other studies and demonstrated the need for the pathogen to multiply during the enrichment steps to reach concentrations that allow their accurate detection by PCR (Gouws et al., 1998; Myint et al., 2006). Additional advantages of combining enrichment steps and PCR are the reduced chance of PCR inhibition due to the dilution of inhibitory substances present in food and the reduced risk of amplifying DNA from dead cells (Aabo et al., 1993; Cocolin et al., 1998; Sharma and Carlson, 2000).
In the PCR protocol developed in this work, the evaluated incubation time of BPW was 18 h because it is used in the conventional method (ISO, 2002). In future studies using this PCR assay, shorter time of incubation should be tested because in other studies the optimal incubation period varied from 5 h to 18 h (Trkov et al., 1999; Yeh et al., 2002; Croci et al., 2004; Myint et al., 2006). Agarwal et al. (2002) tested various foodstuffs and obtained different detection limits for different incubation times, and suggested the use of an enrichment period of 6 h for foods in general. This period is the same as that suggested by Ferretti et al. (2001).
Conclusions
The PCR protocol proposed in this study, which used the SIFBF and SIFBR primers to detect Salmonella in beef samples, can only be validated after assays with naturally contaminated samples, because the performance of PCR using these types of samples may differ from that using spiked ones (Candrian, 1995; Gouws et al., 1998; Waage et al., 1999). It is also essential to conduct interlaboratory studies to assess the reproducibility and robustness of the technique (Malorny et al., 2003b; Hoorfar et al., 2004). This PCR protocol, after validation, may be implemented as a routine test in food analysis laboratories and industries, being valuable even to detect Salmonella in early stages of the conventional protocol by testing RVS and MKTTn after incubation and saving time. It may also become the basis of the development of other molecular methods for the detection of Salmonella, such as nested PCR and real-time PCR.
Footnotes
Acknowledgments
Research labs at Federal University of Paraná (Palotina, PR), São Paulo State University (Botucatu, SP) and University of São Paulo (São Paulo, SP) have provided bacterial strains used in this study. Serotyping of samples was performed by the National Reference Center of Bacterial Intestinal Infections—FIOCRUZ (Manguinhos, Rio de Janeiro, Brazil). The authors are thankful to CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico), CAPES (Coordenação de Aperfeiçoamento Pessoal de Nível Superior), and FAPEMIG (Fundação de Amparo a Pesquisa do Estado de Minas Gerais).
Disclosure Statement
No competing financial interests exist.