
Abstract
Background:
A novel pathotype, Shiga toxin–producing Escherichia coli O104:H4, was the cause of a severe outbreak that affected European countries, mainly Germany, in 2011. The effect of different regimens of rifampicin and gentamicin were evaluated to determine possible treatment modes for the novel strain, and to evaluate the SOS response and its effect on toxin release.
Materials and Methods:
Pulsed-field gel electrophoresis (PFGE) was performed on the novel E. coli O104:H4 pathotype and two pre-outbreak E. coli O104:H4 CDC strains. Transcript levels of the stx2 and recA gene (SOS response inducer) were evaluated using quantitative real-time reverse transcriptase polymerase chain reaction (qRT-PCR) in the novel E. coli O104:H4 samples subjected to different regimens of rifampicin and gentamicin. Consequently, reverse passive latex agglutination (RPLA) was used to determine the Stx2 titers in these samples. Western blot was performed to determine the LexA levels (SOS response repressor) in E. coli O104:H4. The efficacy of treatment with antimicrobial agents was assessed in BALB/c mice.
Results:
The outbreak and pre-outbreak strains are closely related as shown by PFGE, which demonstrated slight genomic differences between the three strains. The transcription level of the stx2 gene in the new pathotype was 1.41- and 1.75-fold that of the 2009 EL-2050 and 2009 EL-2071 pre-outbreak strains, respectively. Moreover, the transcription level of the stx2 gene in the new pathotype was substantially decreased as a result of treatment with the different concentrations of the antimicrobial agents, but was enhanced when the antibiotics were administered at two subinhibitory levels. RPLA data were in accordance with the qRT-PCR results. E. coli O104:H4 exposed to gentamicin at both sub–minimum inhibitory concentration (MIC) levels led to high transcription levels of the recA gene and lack of expression of the LexA protein, implying that the SOS response was activated. Rifampicin at both sub-MIC levels resulted in low transcript levels of the recA gene, indicating that the SOS response was not induced. In vivo, the highest survival rate in BALB/c mice was observed in the group that was treated with the minimum bactericidal concentration (MBC) of gentamicin.
Conclusion:
The use of antimicrobial agents in E. coli O104:H4 infection seems to be effective at the MIC and MBC levels. This provides a promising ground for treatment of E. coli O104:H4.
Background
E
Treatment of infection with Shiga toxin–producing E. coli (STEC), including E. coli O104:H4, is mainly supportive (Borgatta et al., 2012). The use of antimicrobial agents in the treatment of STEC is controversial due to its association with an increased risk of HUS development. The use of antimicrobial agents in the treatment of such infections might exacerbate the disease by causing lysis of the bacterial cell, resulting in enhanced release of Stxs that might have been stored in the bacteria. In addition, the increased release of Stxs might also be related to the activation of a DNA repair system in E. coli known as the SOS response, which reduces the efficiency of treatment (Kimmitt et al., 2000). The SOS response system is made up of more than 50 genes; however, its inducer is the RecA protein and its repressor is the LexA protein (Boutry et al., 2013).
Earlier studies conducted on E. coli O157:H7 strains indicated that using the minimal inhibitory concentration (MIC) of rifampicin that inhibits toxin expression, prior to administration of a bactericidal antibiotic, gentamicin, at the minimal bactericidal concentration (MBC), can be an approach for treating EHEC infections, and showed that rifampicin and gentamicin at their subinhibitory concentrations (sub-MICs) did not activate the SOS response in E. coli O157:H7, denoting that these antimicrobial agents are efficient for treatment purposes (Rahal et al., 2011b; Rahal et al., 2012; Nassar et al., 2013). Consequently, this study aimed at determining whether antimicrobial agents can be used in the treatment of the novel Escherichia coli O104:H4 strain infections. Therefore, the effects of rifampicin and gentamicin were evaluated at the levels of MIC and MBC in vitro and in vivo. Antimicrobial agents initially administered at the MIC or supra-MIC dose eventually reach sub-MIC levels in tissues until the next administration (Reaze et al., 2006). Therefore, the effects of rifampicin and gentamicin were evaluated at two sub-MIC levels to assess the SOS response and its effect on toxin release. In addition, the relatedness of the outbreak and two pre-outbreak strains was assessed.
Materials and Methods
Bacterial strains
The outbreak strain D3774/C22711, which was isolated during the 2011 outbreak, was obtained from Statens Serum Institut in Denmark. The two pre-outbreak strains 2009EL-2050 and 2009EL-2071 were acquired from the Centers for Disease Control and Prevention (CDC). These strains were isolated from cases with bloody diarrhea in the Republic of Georgia in 2009.
Pulsed-field gel electrophoresis (PFGE)
PFGE was carried out using XbaI enzyme on the outbreak and the two pre-outbreak E. coli O104:H4 strains to determine their genomic relatedness using the standard operating procedure for Pulsenet PFGE of E. coli O157:H7, E. coli non-O157 (STEC), Salmonella serotypes, Shigella sonnei, and S. flexneri (Ribot et al., 2006).
MIC and MBC
The MIC and MBC of rifampicin and gentamicin for the outbreak strain D3774/C22711 was determined using the broth dilution method according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI) (CLSI, 2009).
Quantitative real-time reverse-transcription polymerase chain reaction (qRT-PCR)
Three different protocols were followed to prepare the bacterial suspensions from which RNA was extracted.
1. First protocol: To assess the difference in the stx2 gene transcription levels, a bacterial suspension of 1×106 colony-forming units (CFU)/mL of each of the outbreak and the two pre-outbreak strains were inoculated into 1-mL Mueller Hinton II Broth and the samples were incubated at 37°C for 16 h.
2. Second protocol: In order to study the effects of rifampicin, gentamicin, or both at the MIC and MBC levels, on the outbreak E. coli O104:H4 strain, a number of samples (samples 1–8) were prepared as shown in Table 1.
3. Third protocol: To determine the effects of subinhibitory concentrations of rifampicin and gentamicin on the outbreak E. coli O104:H4 strain, another set of samples was prepared (samples 9–13 in Table 1). All incubations took place in a shaker incubator at 37°C for 2 h only with vigorous agitation, since the SOS response remains activated for about an hour following its induction, after which it terminates (Janion, 2008).
CFU, colony-forming units; MIC, minimum inhibitory concentration; MBC, minimum bactericidal concentration.
RNA was extracted from the following: (1) 106 CFU of each of the samples in the first and second protocols; and (2) 108 CFU of each of the samples in protocol 3 using the IIustra RNAspin Mini RNA Isolation Kit (GE, Amersham Place, United Kingdom) according to the manufacturer's specifications for bacterial cells. The QuantiTect® Reverse Transcription Kit (Qiagen, Hilden, Germany) was then used to synthesize cDNA following the manufacturer specifications from the extracted RNA adjusted to have a concentration of 10 μg/mL.
Successively, RT-PCR for (1) the stx2 gene in all the samples from the three protocols and (2) the recA gene in the samples in protocol 3, using the QuantiFast™ SYBER® green PCR kit (Qiagen, Germany), was performed. The primers were obtained from Thermo Scientific (Ulm, Germany), and were described in previous studies (Jinneman et al., 2003; Shilpakala et al., 2009). Reaction mixtures of 20 μL for each sample were prepared and each sample was run in triplicates for the gene in inquiry (stx2 or recA) and the housekeeping gene (rpoB). The housekeeping gene was rpoB; this has been used in several studies to assess the transcription levels of stx2 and resulted in reliable data (Rahal et al., 2011b; Chen et al., 2013; Nassar et al., 2013). Each reaction mixture contained the following: ×1 QuantiFast SYBR Green PCR master mix, 5 μM of the forward and reverse stx2 primers, and 1100-ng/μL cDNA samples. Incubation conditions of a single cycle of 95°C for 15 min, 45 cycles of: 95°C for 10 s, Ta for 30 s, and 72°C for 20 s were achieved using BioRad CFX96 Real Time System, C1000 Thermal Cycler (Munich, Germany). The transcription level of the gene in question was calculated using the BioRad CFX manager software in either the outbreak strain sample compared with the pre-outbreak strain samples in the first protocol or samples treated with antimicrobial agents compared to samples without antimicrobial agent in the second and third protocols, employing the rpoB gene as a standard.
Reverse passive latex agglutination (RPLA)
RPLA was carried out on two different sets of samples. The first set included the outbreak E. coli O104:H4 strain exposed to rifampicin, gentamicin, or broth at the MIC and MBC levels as shown in Table 2 (samples 1–8). The growth inhibition in the samples treated with the MIC or MBC doses of the antimicrobial agents was taken into consideration by making sure that the bacterial density of the drug-free sample was equal to that of the samples treated with the antimicrobial agents. The second set comprised the outbreak E. coli O104:H4 strain subjected to two sub-MICs of rifampicin and gentamicin, as shown in Table 2 (samples 9–13). RPLA was performed using VTEC-RPLA kit (Oxoid, Tokyo, Japan) in order to compare the differential production of Stx2 in these samples.
MIC, minimum inhibitory concentration; MBC, minimum bactericidal concentration; TSB, trypticase soy broth.
BALB/c mice
Adult, male and female, 4–8-week-old mice weighing 19–41 g were obtained from the Animal Care Facility at the American University of Beirut. The mice were cared for and handled according to “Guide for the Care and Use of Laboratory Animals” (National Research Council Committee, 2011). Although the preferable route of mouse inoculation is oral, intraperitoneal infection was used since BALB/c mice are resistant to STEC infection via the oral route as was done in previous studies (Rahal et al., 2011a, 2011b, 2012). STEC do not colonize the mouse gastric system unless oral antimicrobial agents are given. The use of additional antimicrobials would bias the results since the purpose of the study is to assess the effects of particular antimicrobial agents by themselves in a controlled experimental setting. The LD50 of E. coli O104:H4 in BALB/c mice was identified using the method described by Nowotny (Nowotny, 1979). Subsequently, the efficacy of using rifampicin, gentamicin, or both at the MIC and MBC concentrations to treat BALB/c mice infected with E. coli O104:H4 was assessed. As a result, 80 mice were divided into 10 groups, as shown in Table 3. The in vivo MIC and MBC equivalent doses were determined as described in the previous studies. A control group mock treated with sterile broth and a drug control group that received sterile free broth and antimicrobial agents were included. The injections were prepared in a way such that the maximum total volume to be administered per mouse would not exceed 0.5 mL. The first injections were administered at the beginning of the experiment (hour 0), the second injections were given 1 h later (hour 1), and the third injections were administered 16 h after the second injections (hour 17). The mice were monitored for a period of 2 weeks for weight loss and survival.
MIC, minimum inhibitory concentration; MBC, minimum bactericidal concentration; TSB, trypticase soy broth.
Western blotting
Protein assay
Protein was extracted from samples of the outbreak strain grown in antimicrobial-free broth or subjected to sub-MIC 1 and 2 of rifampicin and gentamicin using bacterial lysis buffer prepared as previously described (Nassar et al., 2013). Bradford assay was then carried out to determine the concentration of the extracted proteins.
Western blot
Western blotting was done to study the expression of the LexA protein and L9 ribosomal protein (loading/ housekeeping protein control) levels using previously described antibodies (Nassar et al., 2013).
Statistical analysis
Statistical analysis was done for the transcription levels of the stx2 and recA genes using unpaired t-test using the GraphPad t-test calculator. In addition, the Kaplan–Meier method was used to estimate the survival rates for each group and the log rank (Mantel–Cox) test was used to compare them. All p-values<0.05 were considered to be statistically significant.
Results
The DNA banding patterns revealed by PFGE analysis of the outbreak strain and the pre-outbreak strains 2009EL-2050 and 2009EL-2071 indicated genomic relatedness of 86.4% and 83.7%, respectively (Fig. 1). The pulsetype profiles obtained using the outbreak and pre-outbreak Georgian strains 2009EL-2071 and 2009EL-2050 are similar to those reported by Scheutz et al. (Scheutz et al., 2011). However, the codes provided in Scheutz et al. are different from those provided by the Statens Serum Institut in Denmark for the outbreak strain and the CDC for the pre-outbreak strains.

Dendrogram of the pulsed-field gel electrophoresis gel for the outbreak Escherichia coli O104:H4 strain D3774/C22711 and the two pre-outbreak E. coli O104:H4 strains 2009EL-2050 and 2009EL-2071.
qRT-PCR
The qRT-PCR analysis indicated that the stx2 gene was expressed in the outbreak and pre-outbreak strains. In addition, the stx2 gene transcript level in the outbreak strain was 1.41 times (p-value: 0.0001) and 1.75 times (p-value: 0.0001) that of the 2009 EL-2050 and 2009 EL-2071 pre-outbreak strains, respectively (Fig. 2). Furthermore, qRT-PCR analysis showed that the treatment of the outbreak strain with the MIC of rifampicin (16 μg/mL) caused an 80% decrease in the transcript levels of the stx2 gene in comparison to the control (E. coli O104:H4 without antimicrobial agent) (p-value: 0.0448). Conversely, complete inhibition of the transcription of the stx2 gene was observed upon treatment of the outbreak strain with the following: the MBC of rifampicin (32 μg/mL) (p-value: 0.009), the MBC of gentamicin (4 μg/mL) (p value: 0.0230), the MIC of rifampicin followed by its MBC (p-value: 0.0137), the MIC of rifampicin followed by the MBC of gentamicin (p-value: 0.0138), and the MIC of gentamicin followed by its MBC (p-value: 0.009). The least decrease in the stx2 transcript level was observed in the sample of the outbreak E. coli O104:H4 treated with the MIC of gentamicin (1 μg/mL) (p-value: 0.0128) (Fig. 3).
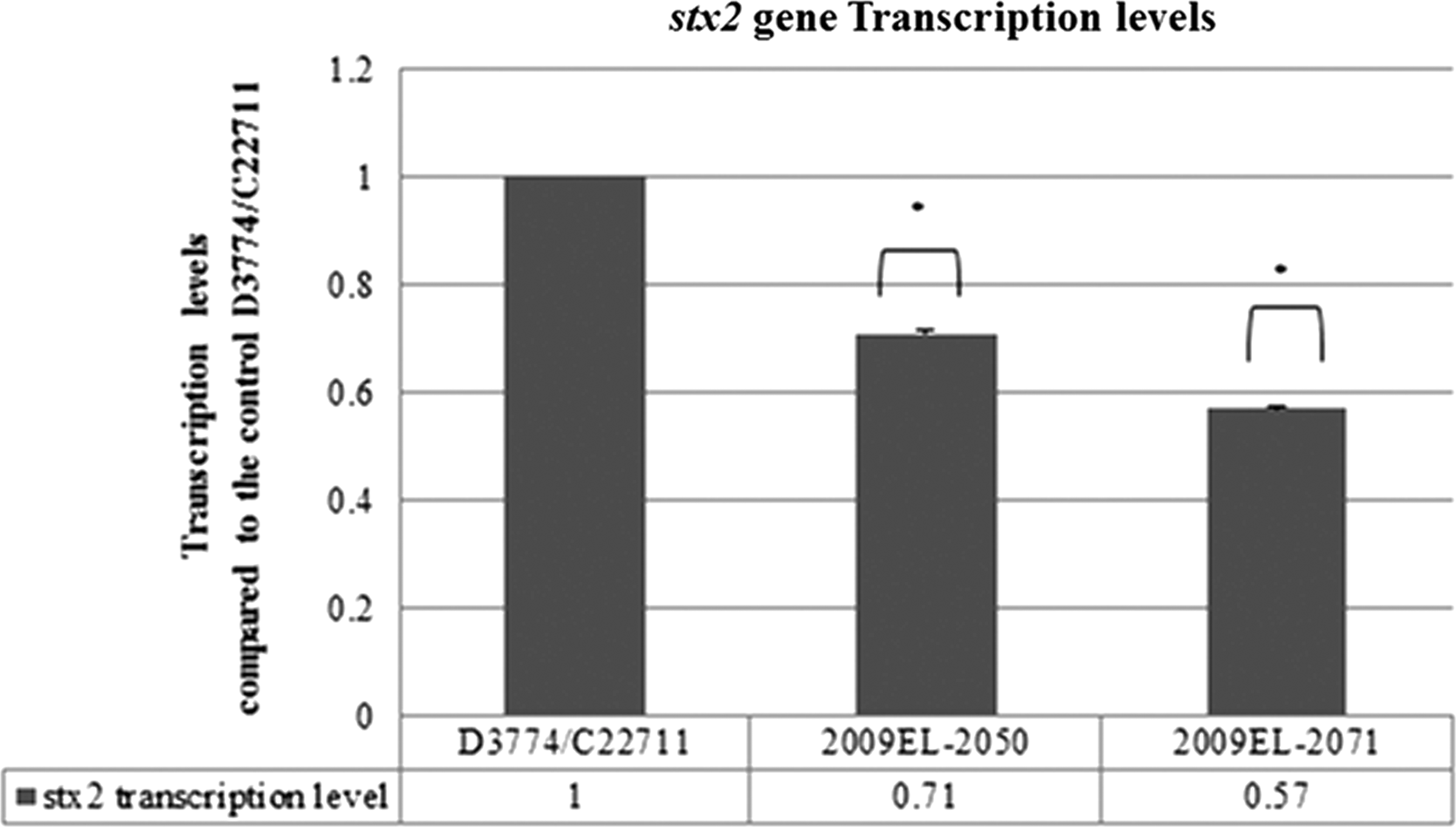
Transcription levels of the stx2 gene in the outbreak Escherichia coli O104:H4 strain D3774/C22711 and the two pre-outbreak E. coli O104:H4 strains 2009EL-2050 and 2009EL-2071. Standard error of the mean values: D3774/C22711: 0.0110, 2009EL-2050: 0.00396, and 2009EL-2071: 0.00576. *p-value<0.05.
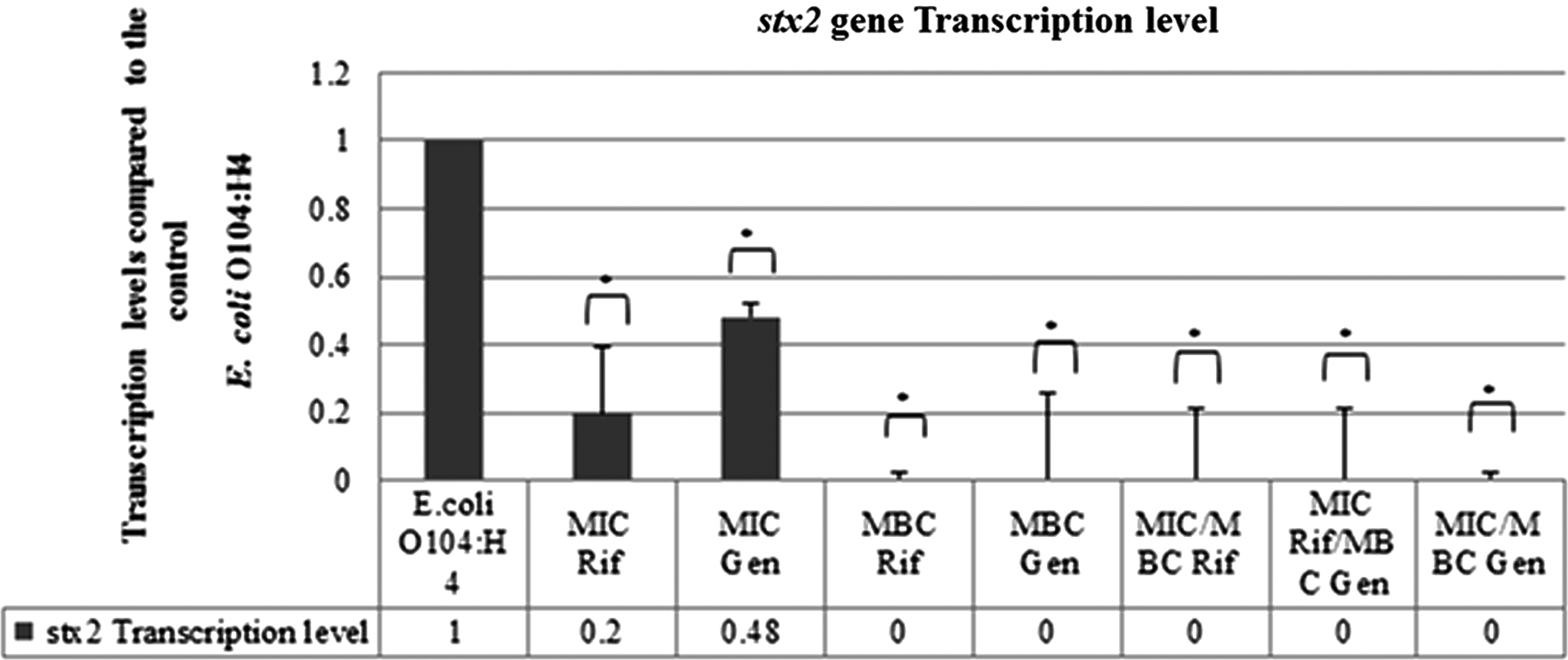
Transcription levels of the stx2 gene in the outbreak Escherichia coli O104:H4 strain D3774/C22711 treated with rifampicin, gentamicin, or both at the minimum inhibitory concentration (MIC) and minimum bactericidal concentration (MBC) levels. SEM values: E. coli O104:H4: 0.11430, MIC Rif: 0.19482, MIC Gen: 0.03904, MBC Rif: 0.0212, MBC Gen: 0.25431, MIC/MBC Rif: 0.20874, MIC Rif/MBC Gen: 0.20989, MIC/MBC Gen: 0.0212. Rif, rifampicin; Gen, gentamicin. *p-value<0.05.
qRT-PCR results showed that the outbreak E. coli O104:H4 samples subjected to the sub-MIC 1 and 2 of rifampicin (8 μg/mL and 4 μg/mL, respectively) led to a lower recA (p-value: 0.0369 and 0.3881, respectively) and higher stx2 transcript levels (p-value: 0.8993 and 0.5674, respectively) in comparison to the control. On the other hand, gentamicin at the sub-MIC 1 and 2 concentrations (0.5 μg/mL and 0.25 μg/mL respectively), resulted in a higher recA (p-value: 0.0859 and 0.0744, respectively) and stx2 gene transcript levels (p-value: 0.2434 and 0.0163, respectively) when compared with the control (Fig. 4).
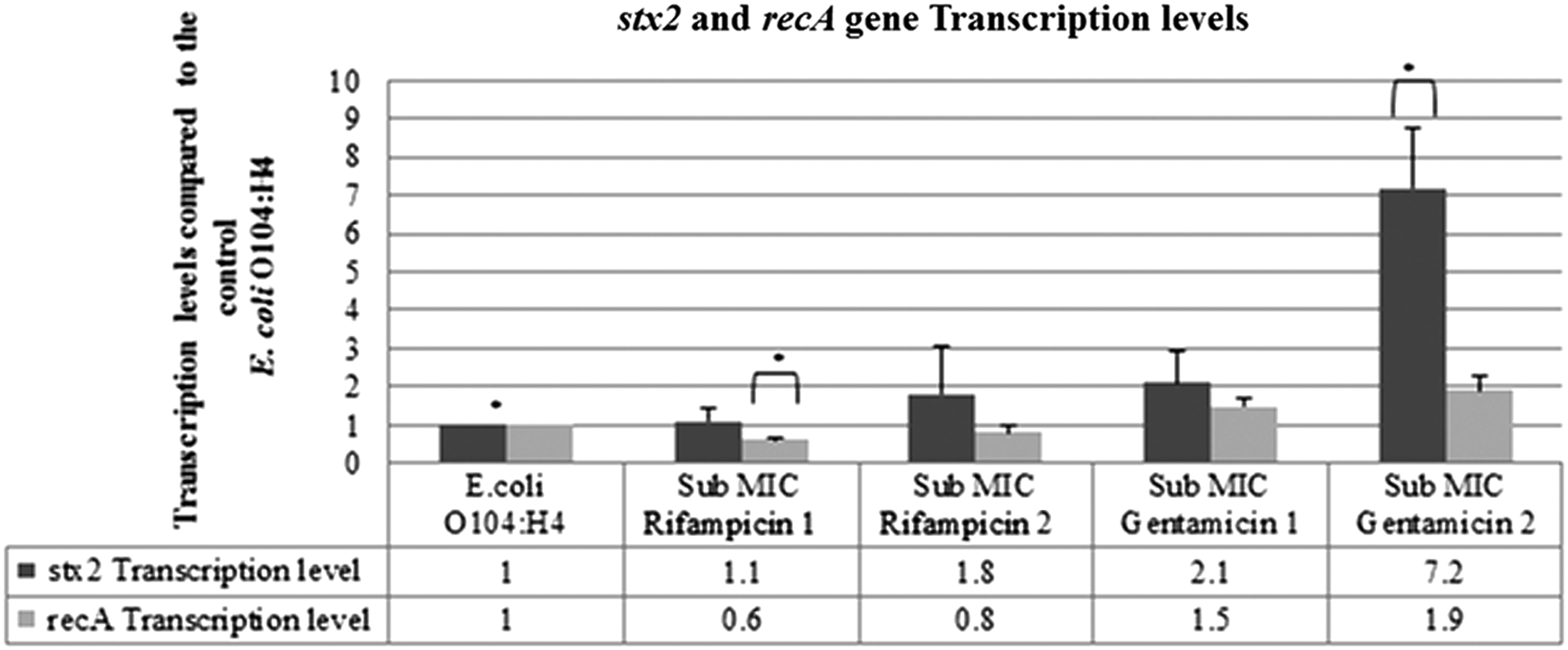
Transcription levels of the recA and stx2 genes in the outbreak Escherichia coli O104:H4 strain D3774/C22711 treated with subinhibitory concentrations of rifampicin and gentamicin. SEM: E. coli O104:H4: stx2 gene: 0.16209, recA gene: 0.11461, sub-minimum inhibitory concentration (MIC) rifampicin 1: stx2 gene: 0.33558, recA gene: 0.08335, sub-MIC rifampicin 2: stx2 gene: 1.28469, recA gene: 0.17978, sub-MIC gentamicin 1: stx2 gene: 0.0859, recA gene: 0.20212, and sub-MIC gentamicin 2: stx2 gene: 0.0744, recA gene: 0.36856. *p-value<0.05.
RPLA
RPLA results indicated that the treatment of the outbreak strain with rifampicin, gentamicin, or both at the MIC and MBC levels resulted in either a twofold or fourfold decrease in Stx2 release in comparison with the control, as presented in Figure 5. On the other hand, treating the outbreak strain with the sub-MIC 1 and 2 of rifampicin resulted in an increase in Stx2 release when compared to the control. Gentamicin at the sub-MIC 1 level produced an equal release of Stx2 in comparison to the control, whereas gentamicin at the sub-MIC 2 level resulted in a twofold increase in the release of Stx2 when compared to the control (Fig. 5).
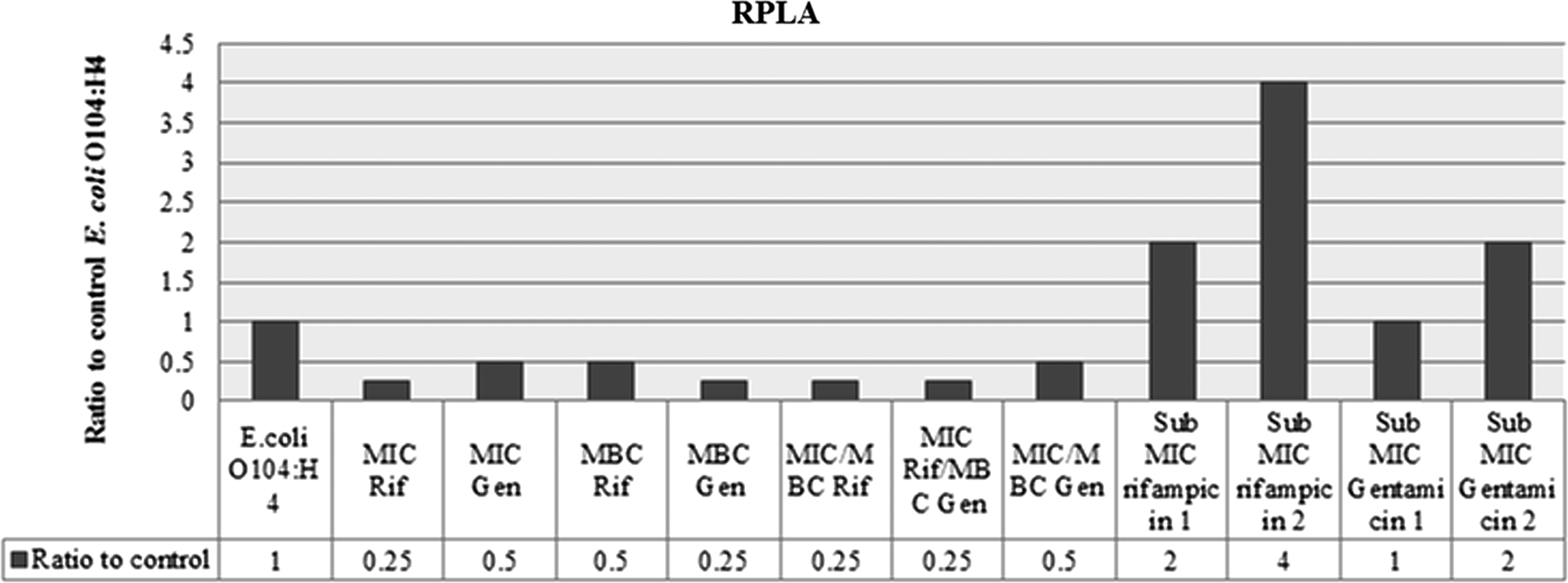
Reverse passive latex agglutination assay (RPLA) for Stx2 in supernatant of the outbreak Escherichia coli O104:H4 strain D3774/C22711 treated with different combinations and concentrations of rifampicin and gentamicin. Rif, rifampicin; Gen, gentamicin; MIC, minimum inhibitory concentration; MBC, minimum bactericidal concentration.
Western blotting
The results of a Western blot assessing the relative levels of LexA in the outbreak strain exposed to the sub-MIC levels of the antimicrobial agents indicated that the expression of this protein was completely inhibited at both sub-MICs of gentamicin (Fig. 6). In addition, rifampicin at the sub-MIC 1 level led to a low expression of the LexA protein when compared to the nonantimicrobial-agent-treated control. However, the sub-MIC 2 of rifampicin resulted in a higher expression of the LexA protein than the control.
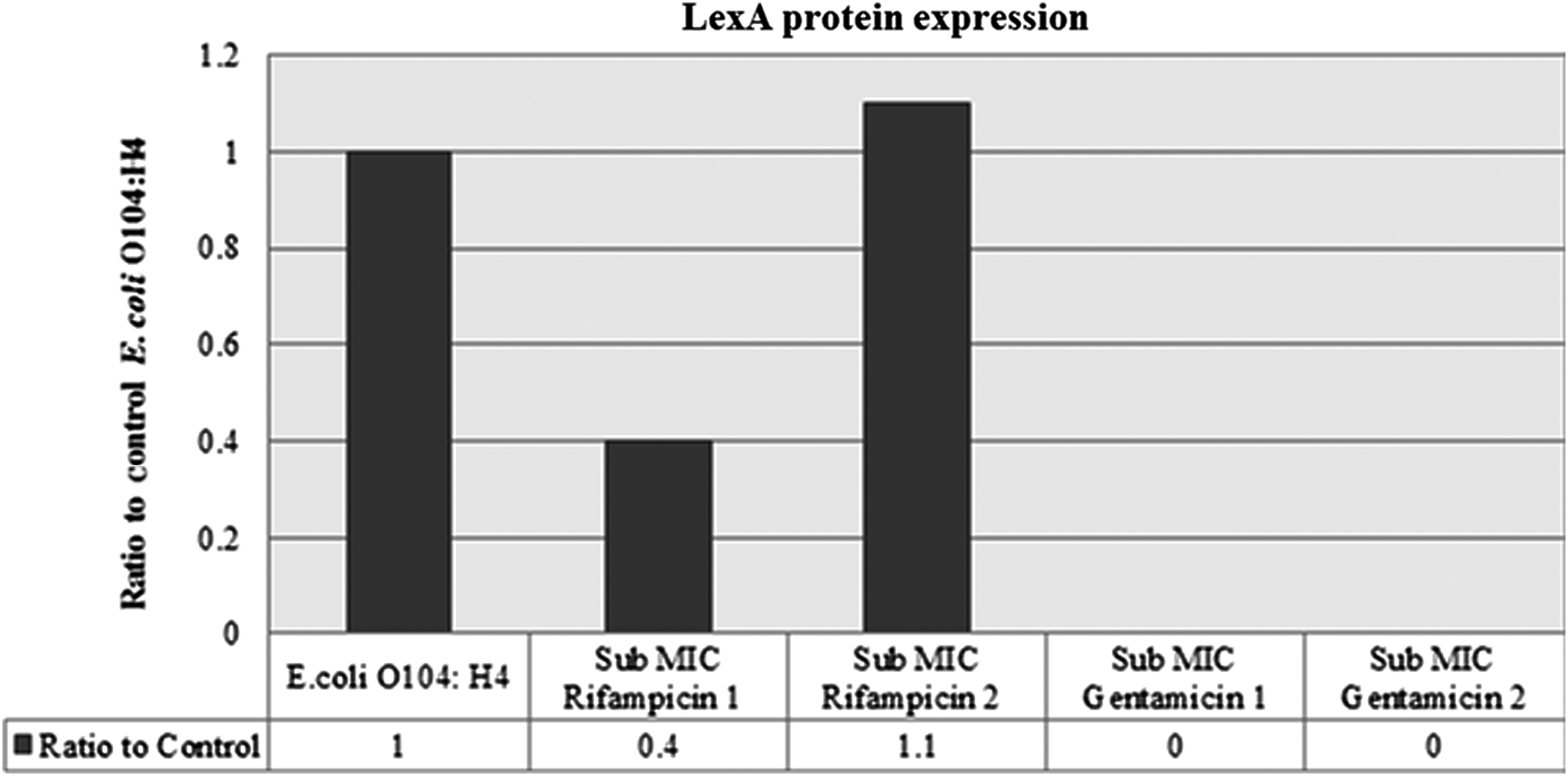
LexA protein expression in the outbreak Escherichia coli O104:H4 strain D3774/C22711 treated with subinhibitory concentrations of rifampicin and gentamicin. MIC, minimum inhibitory concentration.
Mice treatment
The LD50 of the outbreak E. coli O104:H4 strain was determined and was found to be 5.16×106 CFU. The 3×LD50 dose was then used for assessing the effect of antimicrobial treatment in infected mice. All mice that did not receive the bacterial injections (those in the control groups) survived the monitoring period. On the other hand, all mice in the untreated group, infected with E. coli O104:H4 only, were dead 5 days after the infection (Fig. 7). During the monitoring period, the highest survival percentage (50%) was observed in the group of infected mice that received the MBC of gentamicin. A 37.5% survival rate was observed in the infected groups that received the MIC of rifampicin, the MBC of gentamicin, the MIC of rifampicin followed by the MBC of gentamicin, and the MIC of gentamicin followed by its MBC. The lowest survival rate (25%) was observed in the infected group that received the MIC of rifampicin and the group that received the MIC of rifampicin followed by its MBC (Fig. 7). The results were statistically significant according to the log rank (Mantel–Cox) test (p-value: 0.001).
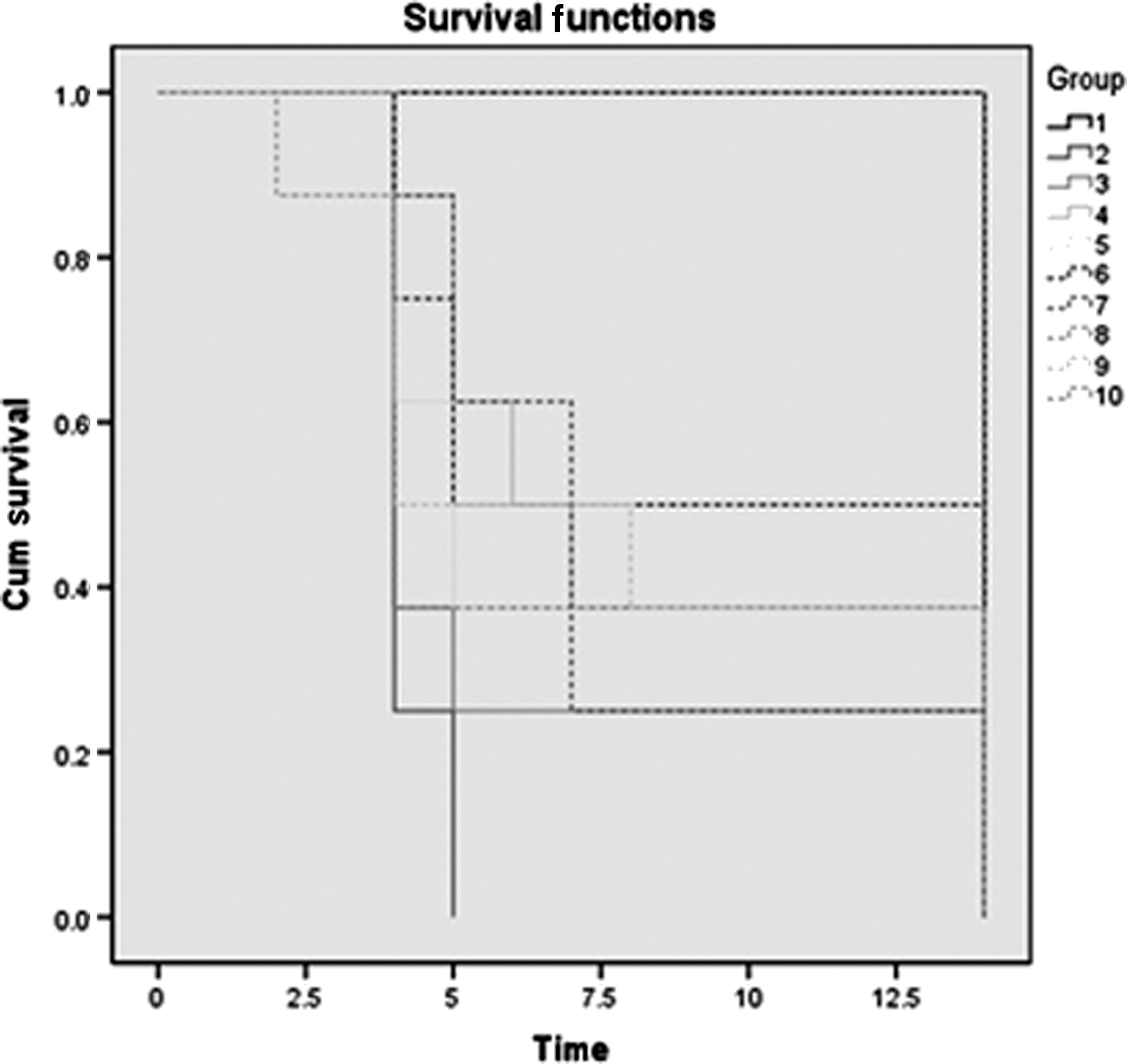
Kaplan–Meier mouse survival curves after infection with the outbreak Escherichia coli O104:H4 strain D3774/C22711 and treatment with different combinations of rifampicin and gentamicin for the 14-day monitoring period. (Group numbers correspond to those in Table 3). Cum, cumulative.
Discussion
STEC O104:H4, is a rare serotype and only sporadic cases have been reported in humans (Scheutz et al., 2011). The similarity between the new pathotype and the two pre-outbreak strains determined by PFGE in this study suggests that the three different strains might have diverged recently from a common ancestor. Our results are in accordance with those of the study performed by Ahmad et al., which showed that the Georgian isolates are the closest genomically to the outbreak strain so far; however, several differences in their stx2 prophages, genomic islands, and plasmids are evident (Ahmed et al., 2012). Furthermore, our study showed that the transcription levels of the stx2 gene differed between the Georgian and the outbreak strains. This might be due to differences in phage genotype in the outbreak and pre-outbreak strains. Drug-free cultures of E. coli strains, harboring diverse stx2 prophages, may yield a wide range of Stx2 toxin production (Wagner et al., 1999).
Antimicrobial treatment of STEC infections has been controversial, as it is associated with an increased frequency of HUS (Wong et al., 2000; Dundas et al., 2001; MacConnachie and Todd, 2004). Antimicrobial agents may augment the production of Stxs from STEC strains due to bacterial lysis and consequent release of Stxs (Kimmitt et al., 2000). Potential modes of treatment using different regimens of rifampicin and gentamicin were assessed in this study. The in vitro results concerning the transcription levels of the stx2 gene and Stx2 toxin production indicated that subjecting the outbreak strain to the different regimens of the antimicrobial agents at the MIC and MBC levels was effective in lowering the expression of the gene and protein. Similarly, the study done by Rahal et al. showed a lowering effect of different combinations of rifampicin and gentamicin at the MIC and MBC levels on the stx2 transcription levels and Stx2 release in E. coli O157:H7 (Rahal et al., 2011b).
The different treatment regimens applied in vitro were tested in vivo, in a mouse model. Groups that received either the MIC or MBC of gentamicin had a higher survival rate than groups that were treated with either the MIC or MBC of rifampicin, respectively, possibly due to the observation that E. coli develops resistance to rifampicin when used singly as therapy (Tarr et al., 1990). The highest survival rate was observed in the group of infected mice that received the MBC of gentamicin. In contrast, the in vivo study done by Rahal et al. showed that mice that were initially infected with E. coli O157:H7 and then treated with the MBC dose of gentamicin died (Rahal et al., 2011b; Rahal et al., 2012). The difference between the treatment of E. coli O157:H7 and E. coli O104:H4 may be due to the fact that E. coli O157:H7 can produce Stx1, which is stored in the periplasmic space, and Stx2, which is released to the extracellular media (Shimizu et al., 2009). Therefore, gentamicin-mediated elimination of E. coli O157:H7 might rupture of the bacteria and lead to enhanced release of stored toxins, making the infection worse. Gentamicin does not seem to affect the outbreak E. coli O104:H4 strain similarly, since the bacteria produce Stx2 only.
Enhanced toxin production in E. coli could also be due to the induction of the stx-encoding prophages, activated by the SOS response (Kimmitt et al., 2000). The SOS response can be activated by a number of factors, one of which is antimicrobial agents. Subsequently, the effect of the sub-MIC levels of antimicrobial agents on the induction of release of Stx2 via the SOS response was assessed. Gentamicin at both sub-MIC levels induced high recA gene transcript levels, completely inhibited the LexA protein expression, and led to high stx2 gene transcript levels and Stx2 toxin release, indicating that the SOS response was activated. The stx2 gene transcript levels at both sub-MICs were higher when compared to the Stx2 release. This could be due to the mode of action of gentamicin, as it usually affects protein synthesis. In contrast to our study, the study by Nassar et al. demonstrated that gentamicin at the sub-MIC levels did not activate the SOS response in E. coli O157:H7 (Nassar et al., 2013). Although gentamicin at the MIC and MBC levels proved to be effective in treating the outbreak strain, gentamicin at the sub-MIC levels might induce the SOS response and lead to an increased release of Stx2. Therefore, gentamicin should be further assessed prior to employment in the treatment of the outbreak strain infections.
Rifampicin at the both sub-MIC levels resulted in a lower recA gene transcript level, higher transcript level of the stx2 gene, and toxin production when compared to the control, indicating that the SOS was not activated. In addition, the expression of the LexA protein was reduced at the sub-MIC 1 of rifampicin; however, its expression was increased at the sub-MIC 2 of the antimicrobial agent. The transcription level of the recA gene at the sub-MIC 1 of rifampicin was low possibly due to the lag period between the transcription of the recA gene and its translation. This could also be due to a possibility that an intermediate DNA damage and subsequently an intermediate level of the LexA protein can elicit the expression of some DNA repair genes without altering the expression of the recA gene (Brent, 1982). In addition, the magnitude of the increase in the Stx2 release was higher than that in the stx2 gene transcript level. Although rifampicin inhibits mRNA synthesis, which is expected to limit protein synthesis, there is a delay in protein inhibition. The rate of RNA degradation (transit time of RNA polymerase) and protein synthesis (lifetime of mRNA) are not necessarily correlated and the rate of the second is higher such that some mRNA remain for some time after being produced (Reid and Speyer, 1970). Furthermore, the transcription of the stx2 gene and the Stx2 release was high even though the SOS response was not activated, possibly due to the existence of additional processes that induce the stx2 phage other than the SOS response. Sigma32-dependent promoters, responsible for the heat shock response, are not affected by rifampicin as much as the primary sigma (sigma70) (Wegrzyn et al., 1998). The heat shock response can be activated by antimicrobial agents, which in sequence can induce the stx phage by a mechanism that is different from the SOS response (Foster, 2005; Rokney et al., 2008). The study by Nassar et al. showed similar results with our study (Nassar et al., 2013).
Few studies were conducted concerning the effect of antimicrobial agents including rifampicin and gentamicin at the MIC and sub-MIC levels on the stx2 gene expression and Stx2 release in the outbreak strain E. coli O104:H4. A study conducted by Corogeanue et al. demonstrated results similar to our study regarding the effect of rifampicin at the MIC level on the transcription levels of the stx2 gene; however, the results were different regarding the Stx2 release (decrease in the Stx2 levels). Moreover, the study showed a similar effect of rifampicin at the sub-MIC level on the stx2 transcription levels and Stx2 release. In addition, it indicated similar results when compared to our study when E. coli O104:H4 was subjected to the MIC and the sub-MIC of gentamicin (Corogeanu et al., 2012). Similar results with gentamicin at the sub-MIC level in one strain of E. coli O104:H4 were observed in another study done by Bielaszewska et al.; however, these results were insignificant (Bielaszewska et al., 2012).
In conclusion, the use of antimicrobial agents in E. coli O104:H4 infections seems to be effective and provides a promising ground for the treatment of human infections with this agent at the MIC and MBC levels. However, the use of the antimicrobial agents at the sub-MIC levels might lead to an increase in the expression of the stx2 gene and hence not be as effective. Therefore, the effect of antimicrobial agents is dependent on the strain, concentration of the drug, and the mode of action.
Footnotes
Disclosure Statement
No competing financial interests exist.