
Abstract
The goal of this study was to monitor Shiga toxin–producing Escherichia coli (STEC) serogroups and virulence genes in cattle (n = 30) originating from a closed herd. Fecal samples were collected (1) at weaning, (2) upon arrival to a feedlot, (3) after 30 days on feed (DOF), and (4) after 135 DOF. DNA was extracted from feces for detection of virulence and serogroup genes by polymerase chain reaction (PCR) and immunomagnetic separation and pulsed-field gel electrophoresis (PFGE) were performed to collect and subtype STEC isolates. The prevalence of each serogroup measured by PCR from weaning to 135 DOF was 23.3–80.0% for O26, 33.3–46.7% for O45, 70.0–73.3% for O103, 36.7–86.7% for O111, 56.7–6.7% for O121, 26.7–66.7% for O145, and 66.7–90.0% for O157. Total fecal samples positive for virulence genes were 87.5% for ehxA, 85.8% for stx 1, 60.0% for stx 2, 52.5% for eae, and 44.2% for the autoagglutinating adhesion gene, saa. The prevalence of each serogroup and virulence gene tended to increase by 135 DOF, with the exception of O121, stx 2, and saa. The frequency of detection of some virulence genes was largely affected over time, most notably with saa and stx 2 decreasing, and eae increasing when cattle were transitioned to concentrate-based diets. PFGE analysis of O157 and O103 fecal isolates revealed dominant pulsotypes, but the presence of identical O103 isolates, which differed in virulence profiles. Overall, this study showed that fecal shedding of E. coli serogroups and virulence-associated genes are highly variable over time as cattle move from ranch to feedlot. To mitigate STEC, it is important to understand the factors affecting both prevalence of individual serogroups and the presence of virulence factors.
Introduction
S
STEC carry genes for two major Shiga toxin subtypes (stx 1, stx 2), which are present in multiple prophages within the genome, and individual strains may encode either stx 1, stx 2, or both genes (Besser, 1999). The virulence gene eae encodes intimin, which is necessary for intimate adherence to the intestinal mucosa (Etcheverria and Padola, 2013). Strains lacking eae may encode an autoagglutinating adhesion (saa) gene putatively involved in adherence, although STEC carrying saa have not been associated with HUS (Jenkins et al., 2003). The plasmid-encoded gene ehxA, which encodes hemolysin, is among the most common virulence factors associated with STEC (Paton and Paton, 1998). The presence of stx 2, eae, and ehxA genes have been associated with an increase in severity of disease in humans, suggesting that certain virulence genes may account for differences in pathogenicity among strains (Boerlin et al., 1999; Blanco et al., 2004; Basu and Tumer, 2015).
E. coli O157:H7 is the most commonly studied serotype and several factors, including age (Ferens and Hovde, 2011), transportation (Schwartzkopf-Genswein et al., 2007), season (Stanford et al., 2005), housing (Synge et al., 2003), and diet (Berg et al., 2004), have been linked to fecal shedding in cattle. The ecology of E. coli O157:H7 in cattle has been intensively studied, but considering the immense diversity of STEC, factors promoting shedding of non-O157 STEC in cattle may differ compared to those influencing shedding of O157. The objectives of this study were to evaluate the distribution of the top seven potentially pathogenic serogroups from a closed herd of cattle sourced from natural pastures at weaning until finishing phases to determine if their distribution varies over time and identify genes associated with virulence.
Materials and Methods
Cattle
Thirty Charolais-sired steer calves (initial body weight 258 ± 27 kg) were sourced from a closed herd with ∼700 cow–calf pairs that were managed under extensive grazing conditions on a yearly basis. Steers were weaned at 196 ± 21 d of age on October 29, 2013. On the day of weaning, steers were transported from the ranch to a commercial auction market (∼3 h duration) and held overnight. The steers were then hauled on a trailer for 15 h before transfer to the Lethbridge Research Centre feedlot, which has a capacity of 600 cattle, to approximate common transportation durations and associated stresses after auction market purchase (Schwartzkopf-Genswein et al., 2007). Calves were housed in three pens (n = 10 per pen) that measured 21 × 27 m. For the first 84 d in the feedlot, steers were fed a silage-based diet (70% barley silage, 25% barley grain, and 5% mineral and vitamin supplement; dry matter basis). Steers were then adapted to a barley grain-based finishing diet (85% barley, 10% barley silage, and 5% supplement; dry matter basis) over a 21 d transitioning period and remained on the finishing diet until slaughter. Cattle were treated in accordance with the Canadian Council on Animal Care (2009) Guidelines.
Sample collection and processing
Fecal samples were collected from individual steers by digital palpation at weaning (sampling time 1), 1 d after feedlot arrival (sampling time 2), after 30 days on feed (DOF), while fed the silage-based diet (sampling time 3), and after 135 DOF, while fed the finishing diet (sampling time 4). Fecal samples were transported on ice to a laboratory for processing and stored at 4°C overnight. Samples were then thoroughly mixed by hand and a subsample (15 g) was mixed with 135 mL E. coli broth (EMD Millipore, Gibbstown, NJ) in a stomacher (Seward Laboratory Systems, Davie, FL) at 230 × g for 1 min. An aliquot of suspension (10 mL) was transferred to a sterile culture tube and enriched at 37°C for 6 h. For polymerase chain reaction (PCR) analysis, 1 mL of the enriched culture was processed for DNA extraction using the NucleoSpin Tissue Kit (Macherey-Nagel, Duren, Germany) according to the manufacturer's instructions.
Detection of serogroup and virulence genes
DNA extracted from fecal sample enrichments was analyzed by separate multiplex PCR (mPCR) for seven serogroups (O26, O45, O103, O111, O121, O145, and O157) and virulence genes (stx 1, stx 2, ehxA, eae, and saa) using primers and conditions described previously (Conrad et al., 2014). O-serogroup-specific positive controls were used for serogroup mPCR (Conrad et al., 2012). For the virulence mPCR, the replication initiation gene repA was used as an internal positive and the positive control included a DNA template mixture (2 μL) of DNA from E. coli O157:H7 strain R508 for repA, ehxA, eae, and stx 2 genes and O26:H11 strain EC19960464 for the stx 1 gene (R.P. Johnson, PHAC, Toronto, Canada) with a sequence-verified plasmid containing the saa gene. All PCRs were performed using a Veriti™ Dx Thermal Cycler (Applied Biosystems, Burlington, Canada).
Bacterial isolation
Glycerol stocks previously stored from all 30 individual cattle from sampling times 1 and 4 were processed for the isolation of O157 and O103 E. coli serogroups. For serogroup O157, bacteria were enriched by immunomagnetic separation (IMS) using anti-O157 bead kits (Invitrogen, Carlsbad, CA) according to manufacturer's instructions. An aliquot of 50 μL of bead–bacteria complex was then streaked onto sorbitol MacConkey agar (MAC) with 2.5 mg/L potassium tellurite and 0.05 mg/L cefixime (CT-SMAC; Dalynn Biologicals, Calgary, Canada) and plates were incubated at 37°C for 18–24 h. Three nonsorbitol-fermenting colonies per plate were randomly selected and tested for O157 serogroup by latex agglutination (Oxoid, Nepean, Canada).
For serogroup O103, bacteria were isolated using the serogroup-specific IMS kit (Romer Lab Technologies) according to manufacturer's instructions. An aliquot of 25 μL of single-wash bead–bacteria complex was then streaked on MAC (Dalynn Biologicals). To confirm specificity of IMS isolation, three random colonies per plate were tested for O103 serogroup by PCR.
Typing of E. coli isolates
Isolates were typed by pulsed-field gel electrophoresis (PFGE) according to the standardized protocol for molecular subtyping of E. coli non-O157 STEC (CDC, 2013). Digested genomic DNA was separated using a CHEF DR II electrophoresis unit with a voltage of 6 V/cm and switch times of 2.2 and 54.2 s for 19 h (Bio-Rad Laboratories, Mississauga, Canada). Strain EDL-933 (accession No. NC_004431) was used as an E. coli O157:H7 reference strain. PFGE gel bands were analyzed using Bionumeric fingerprinting software (version 6.5; Applied Math, Inc., Austin, USA). Cluster analysis was performed applying the Dice coefficient (optimization of 0.5%, position tolerance of 1.0%) and grouped together according to unweighted-pair group method with arithmetic mean. Pulsotypes were created based on identical groupings (>99% similarity) of isolates.
Statistical analysis
Data were analyzed using SAS 9.3 (SAS Institute, Cary, NC). Binomially distributed data (presence/absence) for serogroups and virulence genes were analyzed using sampling time as a repeated measure with animal as the experimental unit in the GLIMMIX procedure of SAS. Type III test effects were used to determine significance (p < 0.05) or tendencies (p < 0.10). Least square means analysis (α = 0.05) was used to evaluate significant differences among STEC, serogroups, and virulence genes over time.
Results
PCR detection of serogroups
All steers (n = 30) harbored at least one serogroup in their feces for each sampling time with the exception of three animals from the sampling time 1, and one animal from sampling times 2 and 3, where feces were negative for all seven serogroup markers (data not shown). The most commonly detected serogroup marker across all sampling times was O103 (75.8%) followed by serogroups O157 (70.0%), O26 (60.8%), O45 (45.8%), O111 (44.2%), O121 (42.5%), and O145 (32.5%; Table 1). The prevalence of serogroups O45 and O103 did not change significantly among sampling times 1, 2, 3, and 4 (p > 0.05). The number of steers carrying O26 increased by sampling time 2, after transportation to the feedlot, and highest prevalence was observed when cattle were at the feedlot for sampling times 3 and 4 (p < 0.05). Serogroups O145 and O111 increased (p < 0.05) and O157 tended to increase (p = 0.07) at sampling time 4 when cattle were fed a concentrate-based diet, compared to sampling time 1 at weaning. Only the prevalence of serogroup O121 decreased (p < 0.05) by sampling time 4 (6.7%) when compared to weaning (sampling time 1; 56.7%). Although, when steers were fed a silage-based diet, (sampling time 3) the prevalence of this serogroup (83.3%) was higher than that at sampling time 1.
Cattle were sampled at weaning (Sampling 1), 1 d after feedlot arrival (Sampling 2), after 30 DOF, while fed a silage-based diet (Sampling 3), and after 135 DOF, while fed a finishing diet (Sampling 4).
Total prevalence of cattle positive for each serogroup across four sampling periods (n = 120).
Values within a row differ (p < 0.05).
Values within a row tend to differ (p < 0.1).
DOF, days on feed.
PCR detection of virulence genes
Across all sampling times, total samples positive for virulence genes were 87.5% for ehxA, 85.8% for stx 1, 60.0% for stx 2, 52.5% for eae, and 44.2% for saa (Table 1). Fecal samples from each animal were positive for at least one virulence gene with the exceptions of two animals from sampling time 1 and two animals from sampling time 4 (data not shown). The number of steers positive for stx 2 was lower at sampling time 4 when steers were fed a concentrate-based diet, compared to the first three sampling times (p < 0.05). Steers positive for saa decreased significantly in finishing phases compared to sampling times 1 and 2 (p < 0.05). Only the prevalence of eae increased significantly, where the number of steers positive was greater at sampling times 3 and 4, compared to sampling times 1 and 2 (p < 0.05).
Across all time points, half of all fecal samples (60/120) were positive by PCR for at least one serogroup present with eae and at least one stx gene. The total prevalence of samples positive for a combination of eae, stx, and at least one serogroup was 20.0% for sampling time 1, 36.7% for sampling time 2, 70.0% for sampling time 3, and 73.3% for sampling time 4. Sampling times 1 and 2 had lower prevalence compared to sampling times 3 and 4 (p < 0.05). One animal in sampling time 3 was positive for all virulence genes, yet was negative for all seven serogroups (data not shown).
Detection and characterization of O157 isolates
No serogroup O157 isolates could be recovered from sampling time 1 and nine isolates from 5/30 animals were recovered from sampling time 4. All O157 isolates recovered were positive for the virulence genes ehxA, eae, and stx 1 and negative for stx 2 and saa. After analysis of PFGE restriction profiles, the O157 isolates clustered into two pulsotypes: I and II (Fig. 1A). Pulsotype I included eight indistinguishable isolates with 88.2% similarity to reference strain EDL933. A single O157 isolate in pulsotype II had 78.0% similarity to pulsotype I.
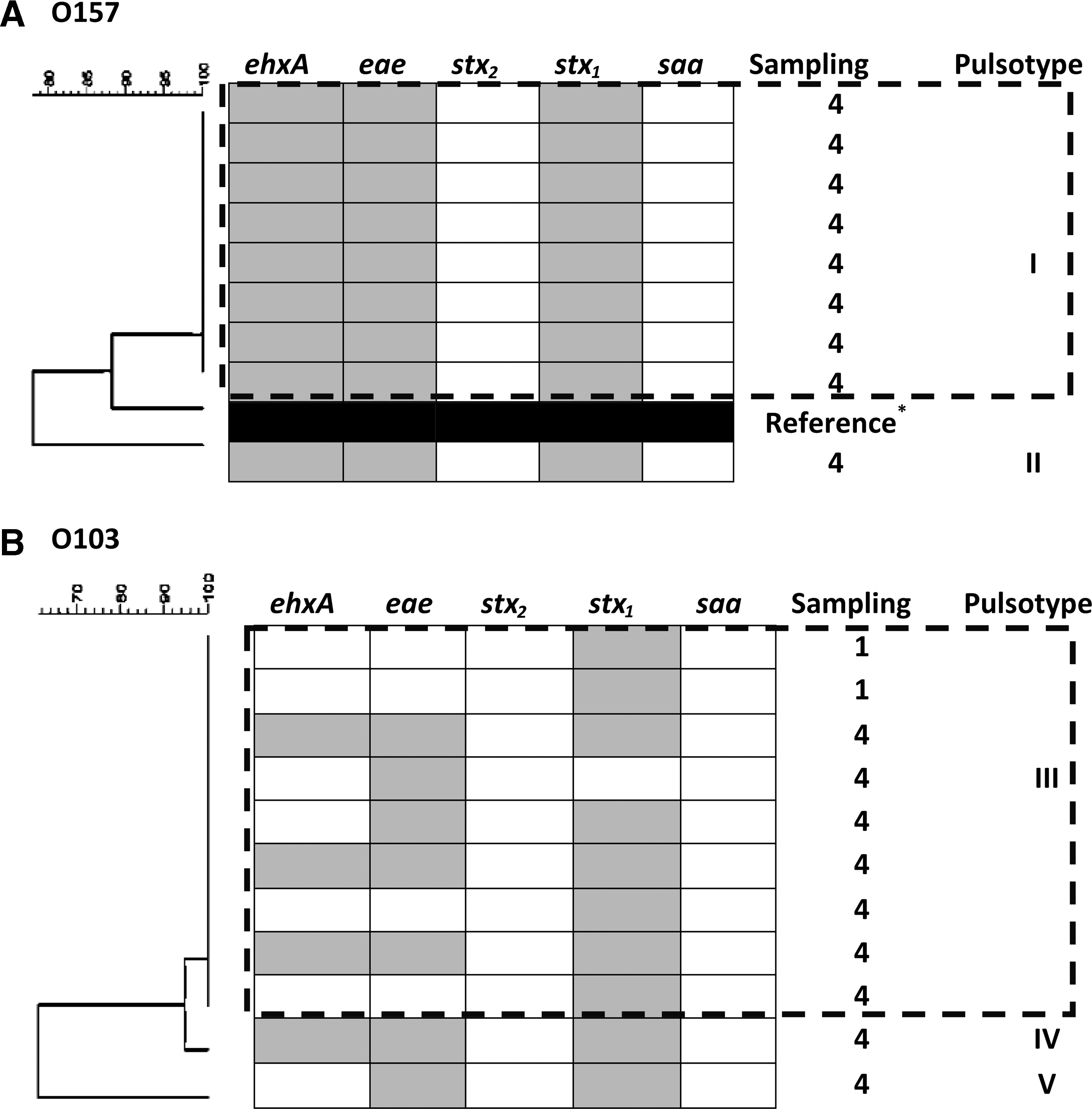
Representative pulsed-field gel electrophoresis dendrograms, virulence gene profiles (ehxA, eae, stx
1, stx
2, and saa), and pulsotypes of XbaI-digested genomic DNA from Escherichia coli O157
Detection and characterization of O103 isolates
Six serogroup O103 isolates from 2/30 animals were recovered from sampling time 1 and 27 isolates from 9/30 animals were recovered from sampling time 4. All isolates tested were negative for stx 2 and saa, but ehxA, eae, and stx 1 genes were heterogeneously distributed among O103 isolates. Each animal contained identical strains that clustered into three pulsotypes grouped as III, IV, and V (Fig. 1B). Pulsotype III isolates included nine animals from both sampling times with identical pulsotypes. A single isolate assigned as pulsotype IV had 94.7% similarity to other O103 isolates and similar banding pattern to pulsotype III, with the exception of the presence of a band at ∼350 kb and absence at ∼100 kb. Pulsotype V included one isolate with 63.0% similarity to other O103 isolates.
Discussion
PCR detection
A substantial proportion of serogroups do not possess genes that encode Stx and there are limitations in correlating virulence genes with specific serogroups without culturable isolates. Despite this, simultaneous screening of serogroups and virulence genes in cattle identifies individuals that have the potential of harboring pathogenic serogroups. The detection of serogroups in this study varied over time, but all serogroups were present in the cattle herd from early weaning until after 135 DOF in agreement to a previous report that found only 2.3% of sampled cattle were negative for the same serogroups analyzed in our study (Paddock et al., 2012).
The most commonly detected serogroups in cattle feces were O103 (75.8%), O157 (70.0%), and O26 (60.8%). Another study similarly found that the top three serogroups detected by direct PCR in feces were O157 (49.9%), O26 (20.3%), and O103 (11.8%; Cernicchiaro et al., 2013). Although the cumulative prevalence was lower than our study, Cernicchiaro et al. (2013) reported within-cohort prevalence estimates that varied from 0% to 78.1%, suggesting that prevalence of these serogroups among cattle subpopulations varies widely. In support of this, pen-level prevalence of STEC has been shown to range from 20.0% to 80.0% (Renter et al., 2004). We observed that the prevalence of serogroups O26, O111, O121, O145, and O157 differed between ranch and feedlot environments. Several factors, including housing, water sources, diet, climate, wildlife exposure, and cattle density can affect the ecology of E. coli in cattle (Smith, 2014). Similar to this study, age has also been previously reported to be a significant factor in STEC prevalence where colonization of STEC persisted longer in calves than adults (Mir et al., 2015). In contrast to these findings, a previous study analyzing STEC in calves reported that isolates of STEC were numerically greatest postweaning and decreased in prevalence before slaughter (Ekiri et al., 2014), although differences in sampling and culture methods make it difficult to compare prevalence estimates.
In the second sampling period, after transportation, the prevalence of serogroups O111 and O121 decreased, whereas O26 increased and O45, O103, O145, and O157 did not change. Transportation is a source of stress to cattle (Schwartzkopf-Genswein et al., 2007) and the close proximity of cattle during transport can lead to increases in hide contamination (Arthur et al., 2007). The number of animals positive for E. coli O157 did not change between weaning and the first sampling at the feedlot, which was preceded by an overnight stay at an auction mart and 15 h of transportation. This suggests that stress and conditions associated with transport to the feedlot did not immediately increase shedding of E. coli O157 and most other serogroups, although we sampled fecal material of individual animals rather than hides.
An increase in the number of O157-, O145-, and O111-positive animals occurred after cattle were shifted from silage to high-grain diets. Dietary changes have been linked to shifts in microbial populations as changes in sugars or starch composition alter the digestive habitat, potentially opening a niche for varying bacteria to occupy (Shanks et al., 2011). The cause of increased shedding after diet change is currently unknown, although several factors, including the ability to adhere to the GI tract, may allow certain serogroups to persist in some animals (Arthur et al., 2010). In contrast, prevalence of O121 was higher when cattle were fed a forage-based diet compared to the finishing diet. This is similar to a previous study where serogroup O121 was most often detected from calves and dams in a pasture-based system (Baltasar et al., 2014), although unknown environmental factors may also contribute to prevalence of this serogroup.
The stx 1 gene was consistently detected in most cattle for the duration of the study. Interestingly, serogroups O26, O111, and O103 have been shown to predominately harbor stx 1 alone (Verstraete et al., 2013) and these same serogroups were among the most commonly detected in this study. The number of cattle positive for stx 2 decreased by 135 DOF. STEC containing stx 2a, stx 2c, and stx 2d have been more frequently associated with HUS than stx 1 (Scheutz et al., 2012), thus it would appear that the risk for cattle carrying genes associated with HUS decreased after 135 DOF. The prevalence of eae in feces was low (23.3%) during weaning, suggesting that cattle at this time did not frequently carry strains that use this highly virulent attachment mechanism. Another study has reported that 95.6% (86/90) of calves were positive for stx genes by PCR, although only 4.2% of isolated STEC encoded eae and 2.4% contained the enterohemolysin gene hlyA (Baltasar et al., 2014). In diarrheic calves, stx genes were detected in 51.3% of fecal E. coli isolates, but only 9.8% carried the eae gene (Nguyen et al., 2011), and in steer calves, the prevalence of STEC in feces ranged from 16% and 23%, yet 5% were positive for eae and 82% for ehlyA (Ekiri et al., 2014). Changes in the GI tract microflora may impact adherence mechanisms (Etcheverria and Padola, 2013), and adherence factors, such as saa, are often present in STEC that lack the eae gene (Bardiau et al., 2010).
Culture isolation
Recovery of isolates was low compared to detection by mPCR, which has been reported previously (Paddock et al., 2012). E. coli O157 isolates with identical gene profiles and similar pulsotypes were isolated from cattle in our study, which is similar to previous reports showing the persistence of dominant O157 strains (LeJeune et al., 2004; Jeon et al., 2013). Each of the O157 isolates was positive for ehxA and eae, an association that has been previously reported in 86.2% of O157 strains (Cookson et al., 2007), with ehxA genes also conserved among E. coli O157 (Verstraete et al., 2013). Most of the O103 isolates had identical pulsotypes, but differed in virulence profiles, confirming that PFGE typing alone is not always enough to discriminate among groups of closely related strains, although some strains persist throughout beef production. These results would support those of previous studies where O103 isolates, some collected over several years, had indistinguishable pulsotypes with differing virulence profiles (Beutin et al., 2005; Verstraete et al., 2013; Ju et al., 2014).
In conclusion, all of the “top seven” E. coli serogroups were detected in cattle from weaning to slaughter, indicating that colonization occurs early in production in a closed herd. The frequency of detection of some virulence genes was largely affected over time, most notably with saa and stx 2 decreasing, and eae increasing when cattle were transitioned to the feedlot. Similar to previous studies of O157, analysis of O103 isolates showed that certain strains persisted in cattle throughout the production cycle, but differed in virulence profiles suggesting higher discriminatory methods are needed for differentiation among these strains. Overall, this study showed that fecal shedding of E. coli serogroups and virulence-associated genes are highly variable over time as cattle move from ranch to feedlot. To mitigate STEC, it is important to better understand the factors affecting both prevalence of individual serogroups and the presence of virulence factors throughout the beef production system.
Footnotes
Acknowledgments
We thank Jenilee Peters, Susanne Trapp, Yidong Han, Pamela Lussier, and Fiona Brown for their assistance in the laboratory, as well as the crew at the Lethbridge Research Centre (LRC) Feedlot for sample collection. This research was supported by a grant from the Government of Canada's Genomic Research Development Initiative and Alberta-Innovates Biosolutions.
Disclosure Statement
No competing financial interests exist.