
Abstract
Emergence and re-emergence of foodborne pathogens is a continuing concern for public health agencies and organizations, the food industry, and consumers. Several factors that contribute to the emergence include changes in the behavior of microorganisms and consumers, changes in agricultural practices and animal husbandry, increase in foreign travel, food distribution through a global marketplace, and climate changes. Furthermore, advances in molecular technologies and pathogen detection methods are allowing increased recognition of the presence of new pathogens. Emerging foodborne pathogens are often zoonotic in origin and may include Gram-negative and Gram-positive bacteria, parasites, and viruses. Previously established foodborne pathogens may re-emerge as more virulent pathogens after the acquisition of new virulence factors, including antibiotic resistance determinants. In this review, various important emerging foodborne pathogens, including non-O157 Shiga toxin–producing Escherichia coli serogroups, pathogenic hybrid E. coli, extraintestinal pathogenic E. coli, drug-resistant foodborne bacteria, Clostridium difficile, hepatitis E virus, and others, are discussed, as well as factors that may be involved in their emergence. Reducing the number of foodborne illnesses and the emergence/re-emergence of pathogens require global partnerships among government agencies, the food industry, and other groups involved in food safety.
Introduction
The food supply chain (i.e., from farm-to-fork) is complex; the composition of foods differs, and there have been changes in the processing, distribution, and consumption of foods, as well as changes in the microorganisms and in agricultural practices. This can result in the emergence of new foodborne pathogens. In the past 30 years, foodborne agents that have emerged include bacteria, viruses, parasites, and biotoxins, and many were identified during outbreak investigations. An established foodborne pathogen may acquire increased pathogenic potential and re-emerge as a new pathogen, and new foodborne pathogens emerge when unrecognized pathogens are identified and linked to foodborne transmission. Known pathogens not previously associated with food transmission are identified as new foodborne pathogens when they are shown to be transferred to humans through food. Many of these new foodborne pathogens arise from animal reservoirs and reach the human population through both animal- and plant-derived foods (Behravesh et al., 2012).
Why New Pathogens Emerge and Old Pathogens Re-emerge
An emerging pathogen has been defined as “the causative agent of an infectious disease whose incidence is increasing following its appearance in a new host population or whose incidence is increasing in an existing population as a result of long-term changes in its underlying epidemiology” (Cleaveland et al., 2007). The definition of emerging infections by van Doorn (2014) is “Emerging infectious diseases are those in which the incidence in humans has increased within the past two decades or threatens to increase in the near future.” There are many factors involved in emergence/re-emergence of foodborne pathogens, and these are listed in Table 1. These include human demographics, consumer attitudes, changes in how foods are processed and handled, in pathogen behavior, and in agricultural practices, inadequate emphasis on education of consumers, inadequate public health services, societal related factors, and an increasing number of multistate (and multicountry) foodborne outbreaks.
A major factor impacting food safety is industrial food animal production with larger farms and high number of animals on each farm, leading to problems of waste disposal and treatment. Also, food processing facilities are fewer but larger, and if there is a microbial problem during processing, the food distribution system may spread the problem through multiple states or even globally. In general, this type of industrial production results in cheaper food available year round (Miller et al., 1998); however, it can also potentially result in major foodborne outbreaks.
Another agricultural issue relates to the two systems for raising swine for meat in the developed world. The conventional method (intensive animal husbandry) is characterized by placing a large number of pigs in a confined enclosed area with no exposure to the outside environment, whereas in the natural system, pigs are allowed to freely roam in open pastures (Thompson, 2010). However, due to contact with wild animals and the potentially pathogen-contaminated environment of pastures, pasture-raised pigs have a higher risk of contracting parasitic and bacterial infections. Gebreyes et al. (2008) indicated that Salmonella seropositivity is higher in free-range-raised pigs. In a review on intensive swine production, Davies (2011) stated that the presence of microorganisms, such as Salmonella, Campylobacter, Listeria, and Yersinia enterocolitica, were similar in both intensive- and free-range-raised pigs. However, free-range pigs are more likely to be exposed to foodborne parasites.
Emerging Zoonotic Pathogens and Zoonotic Diseases
Zoonotic pathogens are organisms that are naturally transmitted between vertebrate animals and humans with or without an arthropod vector (Alemayehu, 2012). Zoonotic diseases have a serious impact on commerce, travel, and economics throughout the world, and they are of concern to risk groups, such as the elderly, children, pregnant women, and immunocompromised individuals. Factors important for the emergence of zoonotic diseases are (1) an increase in exposure to zoonotic agents, (2) an increase in individuals with decreased immune function, (3) an increase in emergence and widespread presence of antibiotic-resistant organisms, (4) differences in crop and animal raising, and (5) differences in processing, distribution, and handling of foods in the global market place (Alemayehu, 2012). At least 75% of recently emerging infectious human diseases have a zoonotic origin, and 60% of human pathogens are zoonoses (van Doorn, 2014).
Important zoonotic foodborne pathogens include Campylobacter spp., Escherichia coli (including O157:H7 and non-O157 STEC [Shiga toxin–producing Escherichia coli], and extraintestinal pathogenic Escherichia coli [ExPEC]), Salmonella, methicillin-resistant Staphylococcus aureus (MRSA), Clostridium difficile, Arcobacter, Helicobacter, Aeromonas, Cronobacter, Cyclospora, Gnathostoma, and foodborne viruses. Poultry is the major reservoir of Campylobacter (Wilson et al., 2008) and Arcobacter (Collado and Figueras, 2011), and poultry is also an important reservoir of nontyphoidal Salmonella (Stevens et al., 2009). In addition, fowl is a reservoir for avian pathogenic E. coli (APEC) strains, which are closely related to human ExPEC (Ewers et al., 2009).
Shiga Toxin–Producing E. coli
STEC are diarrheic foodborne E. coli and major causative agents of hemorrhagic colitis and postdiarrheal hemolytic uremic syndrome (HUS), which can lead to severe kidney disease and death. E. coli O157:H7 has been the major STEC serotype causing HUS; however, the number of infections caused by non-O157 STEC serogroups is overtaking O157:H7 as the main cause of STEC-associated illness (Scallan et al., 2011; Gould et al., 2013). For the period 2003–2012, Heiman et al. (2015) identified 390 outbreaks of STEC O157 in the United States with 4928 illnesses, 1272 hospitalizations, 299 cases of HUS, and 33 deaths. Food was responsible for 255/390 (65.4%) of the outbreaks, and ground beef and other beef products (78/255; 30.6%) were identified as the most frequent food vehicles followed by leafy vegetables (29/255; 11.4%) (Heiman et al., 2015). Majowicz et al. (2014) estimated the annual global incidence of O157 and non-O157 STEC as 2,801,000 acute illnesses, 3890 cases of HUS, 270 cases of permanent end-stage renal disease, and 230 deaths each year. STEC illnesses impact infants and children to a greater degree than adults (Majowicz et al., 2014).
Of the 38 non-O157 STEC single-etiology outbreaks reported during 1990 and 2010, over half occurred during 2007–2010 (Luna-Gierke et al., 2014). Non-O157 STEC belonging to serogroup O111 caused 36.8% of the outbreaks, and serogroup O26 caused 28.9%; 44.7% of outbreaks associated with food, and 39.5% had illnesses transmitted person to person. In general, the non-O157 STEC do not cause as severe a disease as STEC O157:H7, but some non-O157 STEC strains have caused HUS (Gould et al., 2013). The intestinal tracts of food animals, particularly cattle and other ruminants, are reservoirs of both O157 and non-O157 STEC; therefore, during slaughtering operations, the carcass may become contaminated, leading to meat products containing the pathogens (Kaspar et al., 2010). STEC O26, O45, O103, O111, O121, and O145 constitute ∼71% of the non-O157 STEC serogroups isolated with cases of illness in the United States for the years 1983–2002 (Brooks et al., 2005). These serogroups are considered to be adulterants if present in beef (Anonymous, 2011b). STEC that carry the virulence genes, stx2 (encodes Shiga toxin 2, Stx2) and eae (encodes intimin), are associated with more severe infections (Brooks et al., 2005).
Flour: a new vehicle for STEC infections
For the period of December 2015 through September 2016, an STEC-related outbreak occurred involving 56 people from 24 states. All but one of the patients was infected with STEC O121, and one patient was infected with STEC O26. The ages of the patients ranged from 1 to 95 years (median age was 18 years); 77% of patients were female, 16 patients were hospitalized, 1 patient developed HUS, and no deaths were reported (CDC, 2017b; Crowe et al., 2017). Raw flour was implicated at the source of the STEC strains, and the implicated organisms were isolated from the flour. Illness was associated with tasting unbaked dough or batter. In addition, 30 individuals from 6 Canadian provinces were infected with STEC O121, and in a smaller outbreak from another province, 6 individuals were infected with STEC O121 (Harris and Yada, 2017). In an earlier outbreak that occurred in 2009 that was probably linked to flour containing STEC O157:H7, 77 patients from 30 states were involved, and 10 developed HUS. Illness was associated with eating uncooked commercial prepackaged cookie dough (Neil et al., 2012; Harris and Yada, 2017). Since flour may be contaminated with pathogens, it is advisable that consumers not eat raw dough or batter, and children should not be given to play dough made with raw flour.
Hybrid E. coli Pathotypes
An outbreak that occurred in Europe in 2011, involving >4000 infections, ca. 900 cases of HUS, and >50 deaths was associated with eating raw sprouts contaminated with an E. coli carrying an unusual virulence gene combination. It carried stx2a and aggR (plasmid-encoded transcriptional regulator of enteroaggregative E. coli [EAEC]) and was eae negative. Thus, the pathogen had the characteristics of EAEC with the ability to produce Shiga toxin. In addition, the outbreak strain was resistant to multiple antibiotics. The serotype was O104:H4 and was referred to as an enteroaggregative hemorrhagic E. coli (EAHEC) (Beutin and Martin, 2012; Muniesa et al., 2012; CDC, 2013c).
The 2011 EAHEC O104:H4 outbreak made food microbiologists very much aware that hybrid E. coli pathotypes exist, and the clinical significance of EAHEC O104:H4 indicated that such hybrids may be foodborne pathogens. Several other EAEC serotypes carrying the stx2a gene have been shown to cause Stx-related disease in humans. EAHEC serotype O111:H2 was linked to an outbreak of HUS occurring in France involving several children (Boudailliez et al., 1997; Morabito et al., 1998, 1999). A Japanese boy, who developed HUS and encephalopathy leading to death, was infected with EAHEC O86:HNM (Iyoda et al., 2000). In 2012, EAHEC O111:H21 was isolated from a 3-year-old Northern Irish female child with HUS and severe cerebral involvement; the strain caused diarrhea in the child's mother and 4-year-old male sibling (Dallman et al., 2012). EAHEC serotype O59:H− was isolated from a German patient with bloody diarrhea (Prager et al., 2014).
Other hybrid E. coli pathotypes have caused disease in humans. A patient presenting with HUS and bacteremia was infected with hybrid STEC serotype O80:H2 harboring stx2c and stx2d and a conserved virulence plasmidic region characteristic of ColV plasmids of ExPEC (Mariani-Kurkdjian et al., 2014). Soysal et al. (2016) isolated STEC O80:H2 from 54 French patients during 2005–2014. These isolates contained the S88 plasmid associated with APEC and neonatal meningitis E. coli (both are ExPEC), which may explain the bacteremia seen in some of the HUS patients. Therefore, these O80:H2 strains represent STEC/ExPEC hybrids. Experimental urinary tract infection (UTI) was induced in mice when inoculated with STEC serotype O2:H6 strains that carried both STEC- and ExPEC-related genes and were isolated from the stools of diarrheic patients (Bielaszewska et al., 2014).
Hybrids of STEC and enterotoxigenic Escherichia coli (STEC/ETEC), including serotype O2:H27 strains that were eae negative, est1a (gene encoding the heat stable enterotoxin, STa) positive, and carried the stx genes, stx2a or stx2a/stx2d, were isolated from bovine feces and human patients by Nyholm et al. (2015a, 2015b). One child from Finland had a diarrheal infection with an O2:H27 STEC/ETEC strain that was eae negative, and stx2a and STa (heat-stable enterotoxin encoded by esta) positive. Another child developed HUS when infected by an O101:H− STEC/ETEC strain that was positive for the eae and stx2a genes, and the gene coding for STa (Nyholm et al., 2015a). The presence of STEC/ETEC hybrids in animals suggests that animal contact or ingestion of food contaminated with these hybrids could cause illness in humans. Johura et al. (2017) isolated multidrug-resistant STEC/ETEC hybrids with serotypes O76:H19, O43:H2, O87:H16, OR:H2, and O152:H8 in livestock from Bangladesh. In addition, Monday et al. (2006) isolated STEC/ETEC hybrids of serotype Ont:H52 from produce and clinical cases.
An E. coli strain, harboring the eae (encodes intimin involved in the formation of A/E lesions) and elt (encodes the heat-labile toxin) genes but not Shiga toxin genes, was isolated from a child with severe diarrhea, and it was referred to as an enteropathogenic (EPEC)-ETEC hybrid with an atypical EPEC genetic background (serotype not given) (Dutta et al., 2015). STEC isolates from swine often carry a variety of virulence genes typical of ETEC, EAEC, and ExPEC; however, little is known on the potential of these strains to cause human illness (Tseng et al., 2014; Baranzoni et al., 2016). In summary, hybrids of E. coli pathotypes have caused human illness, and given the substantial genomic plasticity of E. coli, other pathotype hybrids will likely be found in the future.
Extraintestinal Pathogenic E. coli
ExPEC are pathogenic E. coli that cause illness outside of the gut. The intestinal pathogenic E. coli are obligate pathogens, whereas ExPEC are facultative pathogens that behave as harmless commensal gut organisms. When they escape the intestinal tract, they can cause illness in nonintestinal sites such as the urinary tract, central nervous system, circulatory system, or the respiratory system (Smith et al., 2007). ExPEC strains are very diverse and have few common virulence factors among them. Virulence genes include those that encode for adhesins, toxins, iron acquisition systems, capsule production, and protectins and invasins (Dale and Woodford, 2015). It is difficult to differentiate ExPEC and nonpathogenic E. coli by molecular epidemiological techniques.
Sporadic community-acquired UTI are common in the United States and involve more than 8 million physician visits per year. E. coli strains that infect the urinary tract (uropathogenic E. coli [UPEC]) cause 70–95% of UTIs; these strains belong to the ExPEC pathotype. UPEC strains invade the cells of the bladder (cystitis), and ascension of the organism through the ureters to the kidney can lead to infection of the kidneys (pyelonephritis). UPEC can enter the urinary tract from the gastrointestinal tract through the urethra (Smith et al., 2007; Lo and Alonto, 2011). Approximately 80% of UTIs occur in women, and 50–60% of women have at least one UTI during their lifetime. Furthermore, recurrent UTI occurs in 20–30% of women. The cost of UTIs to the economy of the United States is ca. 3.5 billion dollars per year (Smith et al., 2007; Lo and Alonto, 2011). UTIs are generally treated with antibiotics but, unfortunately, drug resistance is increasing in UPEC strains (Nordstrom et al., 2013).
UTIs caused by EAEC/UPEC hybrid E. coli strains have been reported (Lara et al., 2017; Nunes et al., 2017). These hybrids were present in feces, and while not shown to be associated with food ingestion, it is possible that they could have been foodborne. Ramchandani et al. (2005) suggested the UPEC had an animal origin and that the organisms might be foodborne. Studies indicate that UPEC are present in animals and food. E. coli O25:H4-ST131 and O114:H4-ST117 (from retail chicken meat) and O2:H7-ST95 (from ready-to-eat honeydew melon) were identical or closely related to clinical UPEC suggesting that dissemination of UPEC by foodborne transmission is possible (Vincent et al., 2010).
Using a mouse model for ascending UTI developed by Hvidberg et al. (2000), Jakobsen et al. (2010a, 2010b) demonstrated that E. coli strains from animals and animal products colonized the bladder and kidneys of the mice. Jakobsen et al. (2010b) isolated phylogroup B2 E. coli (most UTI-associated E. coli belong to phylogroup B2) from broiler feces and meat, as well as from pig feces and pork and inoculated the isolated organisms into the empty bladders of the UTI model mice. E. coli isolated from these sources were found in the bladders and kidneys of the mice. Similar results were found with clonal group A E. coli (phylogroup D) (Jakobsen et al., 2010a). The E. coli isolates from the studies of Jakobsen et al. (2010a, 2010b) behaved similarly in the mouse model as did isolates from UTI patients tested in the same animal model, indicating that UPEC can be isolated from both animals and meat. In another study examining the potential zoonotic risk of ExPEC from a chicken source, isolates from chicken meat and eggs were tested for virulence in rodent models of sepsis, meningitis, and UTI (Mellata et al., 2018). The investigators showed that chicken and egg E. coli isolates were similar to human ExPEC in terms of their potential to cause extraintestinal infections in a mammalian host.
APEC are important disease ExPEC agents of poultry leading to large economic losses to the poultry industry (Dho-Moulin and Fairbrother, 1999). Jakobsen et al. (2010c) found ExPEC virulence genes in E. coli strains isolated from broiler chickens and pigs, as well as from chicken meat and pork and suggested that there may be an animal source for pathogenic ExPEC in humans. Other workers have shown that some human ExPEC and APEC strains have similar phylogenetic backgrounds and share some of the same virulence genes (Ewers et al., 2009; Zhao et al., 2009; Mellata, 2013). It is probable that chickens are an important reservoir for human ExPEC strains, including UPEC strains (Bergeron et al., 2012). This relationship between ExPEC and APEC suggests that APEC may be zoonotic pathogens that can lead to human diseases, such as UTIs through the ingestion of poultry meat.
Outbreaks of community-acquired UPEC-induced UTIs have been reported (George and Manges, 2010), but foodborne outbreaks of UTIs have not been unequivocally documented. Recent data indicate that UPEC can be found in animal feces (particularly chickens), as well as in animal meats (Jakobsen et al., 2010a, 2010b; Vincent et al., 2010), and thus, a large portion of sporadic cases of UTIs are probably foodborne caused by UPEC originating from avian species.
Escherichia Albertii
Escherichia albertii was first isolated from diarrheic children in Bangladesh by Albert et al. (1991, 1992); however, Albert and his coworkers misidentified the causative organisms as Hafnia alvei. A few years later, Huys et al. (2003) described the strains as belonging to a novel taxon in the genus Escherichia and proposed the name E. albertii sp. nov. The organism has been responsible for outbreaks and sporadic cases of foodborne diarrhea. Nimri (2013) considers E. albertii as a newly emerging enteric pathogen.
E. albertii strains have been isolated from Canadian environmental waters, including well water, surface and recreational water, and sewage (Maheux et al., 2014). E. albertii has also been isolated from food, such as chicken giblets (Maeda et al., 2015), raw chicken liver sashimi (Asoshima et al., 2015), and raw meat (pork, mutton, chicken, and duck but not raw beef) (Wang et al., 2016). Sharma et al. (2007) demonstrated that E. albertii strains were more sensitive to heat, acid, and pressure as compared with wild-type E. coli O157:H7, suggesting that food processing conditions that destroy E. coli O157.H7 should also inactivate E. albertii.
E. albertii inoculated into raw ground beef (85% lean) and stored at 5°C showed no growth after 15 d of storage; however, at 22°C, there was ca. 4 log CFU (colony-forming unit)/g increase within 3 d of storage and at 35°C, there was 4–4.3 log CFU/g increase by 24 h storage (Perez et al., 2013). Jones-Ibarra et al. (2016) showed that E. albertii in ground beef (6.6 log CFU/g) cooked to an internal temperature of 62°C, 71.1°C, and 76°C showed reductions of 3.9, 4.4, and 4.9 log CFU/g, respectively.
E. albertii is associated with sporadic disease and epidemics in birds; pathologic features showed evidence of enteritis (Oaks et al., 2010). Wild birds but not poultry appear to be a reservoir of E. albertii; however, the organism has been isolated from poultry, apparently due to contact with environments contaminated with feces from infected wild birds (Gordon, 2011). In a survey of fecal specimens from 102 cattle, 45 swine, and 45 broiler chickens in Japan, Hinenoya et al. (2014) found only 1 E. albertii strain (in 1 pig). Olsen and Besser (2007) did not find E. albertii in diarrheic and nondiarrheic calves (n = 56) or adult cows (n = 66). A number of Australian vertebrate species were tested for the presence of E. albertii, and interestingly, the organism was not found in fish, frogs, snakes, lizards, crocodiles, or mammals. However, it was found in various bird species (Gordon, 2011).
E. albertii was responsible for a foodborne outbreak in Japan in 2003 linked to boxed lunches (Asoshima et al., 2014). An outbreak of gastroenteritis occurred in 2011 in Japan, and Ooka et al. (2013) declared that while two Escherichia species (E. albertii and E. coli O183:H18) were involved in the outbreak, E. albertii was the major causative pathogen. Pathogenic factors in E. albertii that contribute to gastroenteritis include the production of attaching and effacing lesions and the presence of cytolethal distending toxin and the Shiga toxin variant Stx2f (Friesema et al., 2014; Jones-Ibarra et al., 2016). One reason why there have been a limited number of studies in determining the role of E. albertii as a foodborne disease agent is likely due to difficulties in isolation and identification of the organism. It is interesting that most of the reported outbreaks and sporadic cases occurred in Japan. Foodborne illnesses due to E. albertii have not been reported in the United States.
Antibiotic-Resistant Bacteria
Antibiotics are used in humans and animals to prevent, control, and treat disease. Another use for such drugs is for growth promotion in food animals; however, feeding subtherapeutic amounts of antibiotics to animals may lead to increased drug resistance (Mathew et al., 2007; Andersson and Hughes, 2014). Unfortunately, a successful antimicrobial drug is confronted with the potential ability of microorganisms to develop tolerance (i.e., resistance) to the harmful effects of that drug. Antibiotic resistance is a property of microorganisms that gives them the ability to inactivate or exclude a drug or confers upon them the ability to block the inhibitory or killing effect of the drug. Another unfortunate aspect is that fewer and fewer new antibiotics are being developed due to economic and regulatory constraints, and thus there are few new drugs to replace the old, less useful ones (Nathan, 2004).
Annually in the United States, ca. 2 million individuals are infected with antibiotic-resistant bacteria leading to 23,000 deaths. The estimated cost of antibiotic resistance to the U.S. economy is $55 billion annually (Demirjian et al., 2015). Drug-resistant microorganisms enter the food chain because of their presence in food animals. Fecal excretion of drug-resistant organisms leads to eventual contamination of the environment. Thus, animal food products, fruits, and vegetables can be contaminated with resistant microorganisms (Davies and Davies, 2010; Spellberg et al., 2013). Another part of the problem is that antibiotic resistance is rampant in the developing world due to the availability of antibiotics without a prescription, and there could be transfer of resistant microorganisms to developed countries (Reardon, 2014).
Antimicrobial stewardship can be defined as optimal selection, dosage, and duration of the antibiotic with maximizing the benefit of therapy while minimizing harm to the patient and the community and having minimal impact on subsequent resistance (Doron and Davidson, 2011; Demirjian et al., 2015). The proper drug used at the proper dosage for the proper microorganism will aid in prevention of resistance. The development of resistance to antimicrobial agents can be slowed down by eliminating unnecessary antibiotic use in humans and animals. Drug resistance spread can be prevented by immunization, good personal hygiene, and safe food preparation. Tracking antimicrobial-resistant infections, resistant microorganisms, and risk factors for infection will allow the development of strategies that will aid in the prevention of infections and spread of resistant microorganisms. And finally, since antibiotic resistance is due to evolution, new antimicrobials will be needed to fight the current resistant microorganisms, and new diagnostic test development will be necessary to track resistance to new antibiotics (CDC, 2013a).
Martin et al. (2015) reported that 80% of antibiotics in the United States are used in animal production, of which 70% are important in human medicine. Placing a cap on the amount of antibiotics used in animal production and/or imposing user fees for antibiotic usage in food animals have the potential to lead to decreased resistance worldwide (Van Boeckel et al., 2017). The U.S. Food and Drug Administration issued a Guidance for Industry on “The judicious use of medically important antimicrobial drugs in food-producing animals” (FDA, 2012) in efforts to slow the development of antibiotic resistance by phasing out the use of medically important antibiotics in food animals for production purposes and limiting their use in treatments that require veterinary oversight. Action plans to combat antibiotic resistance with outlining strategies for public health, health care, and veterinary partners have been established by the U.S. government in 2015 (
Antibiotic resistance may be intrinsic, and therefore, some microorganisms are naturally resistant to a drug, or drug resistance may be due to gene mutations or to horizontal transfer of DNA containing antibiotic resistance genes. Resistance mechanisms may include: acquisition of efflux pumps that extrude the drug before it can reach the target; enzymatic destruction or inactivation of the drug; and microbial synthesis of new metabolic pathways not inhibited by the drug (Tenover, 2006).
Bacteria resistant to multiple antibiotics are referred to as multidrug resistant, which is defined as acquired resistance to at least one agent in three or more antimicrobial categories (Magiorakos et al., 2012). An E. coli strain resistant to both colistin and carbapenem antibiotics was isolated from the urine of a patient with a UTI (Mediavilla et al., 2016). This is of concern since colistin is used as a treatment of last resort in patients infected with carbapenem-resistant strains. Furthermore, Liu et al. (2017) reported the isolation of E. coli-carrying plasmids harboring mcr-1 (colistin resistance), carrying carbapenem resistance (bla NDM), as well as a plasmid carrying both genes, which may greatly hasten the horizontal transmission of both genes among bacteria.
Krueger et al. (2014) found that of 875 nontyphoidal Salmonella isolates, 705 (80.6%) were pan-susceptible (i.e., susceptible to antibiotics likely to be prescribed by a physician), 165 (19.2%) were resistant to at least one drug, and 55 (6.3%) of the isolates were resistant to five or more drugs. Nontyphoidal Salmonella enterica serotypes are a major cause of foodborne illness worldwide. When the Salmonella infection is severe, antibiotic therapy is required, and there has been an increase in drug resistance in these pathogens (Crump et al., 2011). For example, there has been an increase in fluoroquinolone and third-generation cephalosporin resistance in Salmonella (Hur et al., 2012; Krueger et al., 2014). Some data from the National Antimicrobial Resistance Monitoring System presented in Table 2 show that the percentage of salmonellae from retail chicken that were pansusceptible, decreased from 51.7% in 2002 to 37.1% in 2012, and there has been a several-fold increase in multidrug resistance since 2002. A similar pattern is also seen for ground turkey (Anonymous, 2011a).
Multidrug-Resistant Salmonella in Retail Chicken, United States (Anonymous, 2012)
Methicillin-Resistant S. aureus
S. aureus is a Gram-positive nonspore-forming coccus that is part of the normal flora of the human skin and nose. In addition, it is a major human pathogen causing skin, soft tissue, bone, and blood infections, as well as postoperative wound infections. The organism produces toxins that induce toxic shock syndrome and food poisoning. Penicillin-binding proteins (PBPs), located in the S. aureus cell membrane, bind β-lactam antibiotics, such as methicillin, which results in inhibition of cell wall synthesis, leading to cell death. Methicillin resistance depends on the mecA gene, which encodes a PBP, PBP2a, with reduced affinity for the antibiotic, and thus, cell wall synthesis can occur (Kluytmans, 2010). Paterson et al. (2014) recently reviewed the emergence of mecC MRSA. MRSA-contaminated food was the source of a community-acquired infection in low-risk individuals (Jones et al., 2002).
Hospital-acquired and community-associated MRSA infections are a serious public health concern. In the past two decades, new emerging MRSA clones that cause community-associated infections in individuals with no known risk factors have emerged (David and Daum, 2010). MRSA have been found in animals used for food and in companion animals (Leonard and Markey, 2008; Cuny et al., 2010). The presence of MRSA in animals (livestock-associated MRSA, LA-MRSA) indicates that meat can contain the organisms, and indeed, the data in Table 3 show that MRSA are present in retail beef and pork products.
Methicillin-Resistant Staphylococcus aureus in Food Products
MRSA, methicillin-resistant Staphylococcus aureus.
O'Brien et al. (2012) cited 10 studies examining for the presence of MRSA in meats (beef, pork, chicken, and turkey) from the United States, Canada, the Netherlands, and Spain. Percentages ranged from 0% to 36% of samples that were positive for MRSA. Parisi et al. (2017) found that the prevalence of MRSA in horses in 4 Italian slaughter houses was 15% (26/171), whereas the incidence of MRSA in farm/riding stables and race courses was 1% (1/217). MRSA was detected in 12% (2/16) of personnel who worked in horse slaughter houses. Many of the MRSA-positive horses from slaughter houses were imported from Spain and Poland (Parisi et al., 2017). The authors concluded that individuals who work in horse slaughter houses and those who handle horse meat or eat poorly cooked or raw horse meat are at risk for MRSA colonization. Interestingly, foodborne outbreaks involving MRSA strains have only been reported twice (Table 3), one linked to ingestion of barbecued pork and the other caused by an infected food handler, involving 27 patients and 5 deaths.
Community-Onset C. difficile-Associated Disease
C. difficile is an anaerobic Gram-positive, spore-forming toxin-producing rod that causes antibiotic-associated diarrhea, colitis, and pseudomembranous colitis. Spores present in the intestinal tract remain dormant unless the normal bacterial flora is disrupted. Disruption of the intestinal flora allows the C. difficile spores to germinate with the release of toxins leading to diarrhea or colitis (Juneau et al., 2013). Risk factors for C. difficile infection include antibiotic or proton pump inhibitor use, increased age, gastrointestinal surgery, complicated chronic illness, or prolonged stays in health care settings. Thus, any medication or situation in which there may be a disturbance of the normal intestinal flora can induce C. difficile symptoms (Juneau et al., 2013).
C. difficile is commonly classified as a nosocomial pathogen, and the reservoirs may be health care patients, health care workers, and/or the contaminated hospital or other health care facility environment (Heinlen and Ballard, 2010). More recent studies indicate that C. difficile has great potential as a zoonotic pathogen. The organism is present in companion animals such as dogs, cats, and horses (Freeman et al., 2010; Hensgens et al., 2012; Rodriguez-Palacios et al., 2013). C. difficile may also be present in the intestinal tract of animals used as a food source (Freeman et al., 2010; Hensgens et al., 2012; Rodriguez-Palacios et al., 2013). C. difficile present in food animals such as swine, cattle, and poultry can contaminate meat produced from these animals. The spores of C. difficile persist for long periods in the environment and water supplies contaminated by animal feces.
The data shown in Table 4 indicate that a variety of retail foods, particularly meat products, may contain the pathogen. It is probable that the source of C. difficile in meat products is from the animal itself. Seafood may be contaminated by water containing animal feces, whereas the ingredients in salads may be contaminated by the environment in which they were grown or contaminated by a food handler. While C. difficile is present in food products, foods have not been confirmed as a cause of C. difficile illness (Hensgens et al., 2012).
Clostridium difficile in Food Products
A study conducted in 1991–2005 of C. difficile infections in Minnesota, Khanna et al. (2012) identified 385 cases; 41.0% (158/385) were considered to be community associated. Individuals who acquire health care facility-onset disease are generally elderly (>65 years), have been administered antibiotics, have a more severe illness, and are more likely to have comorbidities. Otten et al. (2010) suggested that the sources of C. difficile for community-associated disease are the environment, contact with infected or colonized patients, contact with infected or colonized animals, and ingestion of contaminated food or water. Interestingly, Marsh (2013) discounts the possibility that foods are a source of community-acquired C. difficile infections; nonetheless, the potential for foodborne transmission appears to be real. Since the spores are heat resistant, cooking processes for most foods will not inactivate spores, and Rodriguez-Palacios et al. (2013) suggest heating foods to 85°C for 10–15 min to minimize the counts or sub-boiling (96°C) for 2–3 min to give a 6 log10 reduction.
It appears that C. difficile infections are increasing; mortality data for the United States indicate that death due to C. difficile increased from 793 in 1999 to 7483 in 2008 (Lessa et al., 2012). C. difficile infections have long been associated with health care institutions; however, community-associated infections make up at least one-third of C. difficile cases (Lessa, 2013). The community-associated cases may be underestimated since younger individuals (who are more likely to develop community-associated C. difficile infections) do not report mild diarrheal illnesses. More research is needed concerning epidemiology, risk factors, control measures, and methods for treatment of community-associated C. difficile infections.
Campylobacter Species
While Campylobacter jejuni and Campylobacter coli are considered the most clinically important Campylobacter species, other species, including Campylobacter concisus, Campylobacter upsaliensis, and Campylobacter lari also cause gastrointestinal illness (Kaakoush and Mitchell, 2012). C. concisus has been found in feces of individuals with intestinal disease, as well as in healthy controls. The organism has been associated with enteritis, Crohn's disease, ulcerative colitis, and inflammatory bowel disease; however, more studies are needed to clarify the role of C. concisus in intestinal diseases (Man, 2011; Kaahoush and Mitchell, 2012). Epidemiological evidence indicates that C. upsaliensis is a cause of gastroenteritis, particularly in children and immunocompromised individuals (Man, 2011). The importance of C. upsaliensis as a foodborne pathogen is not clear; however, Man (2011) reported that C. upsaliensis is an emerging pathogen. C. lari is found in poultry and other domestic food animals and has been associated with gastroenteritis and bacteremia; however, the literature is sparse on the number of cases of C. lari infections (Matsuda and Moore, 2011). Only one outbreak has been reported for C. lari (Man, 2011).
Lynch et al. (2011) reported the frequency (%) of Campylobacter species (jejuni, coli, concisus, upsaliensis, lari) isolated from chicken, pork, and beef meats from Ireland retail markets (Fig. 1). All five species were present in beef meats and four were present in chicken (except C. upsaliensis), indicating that beef and chicken may be reservoirs. Pork meat contained only C. jejuni and C. coli. Bullman et al. (2011) isolated 340 Campylobacter species from the feces of 7194 Irish patients in 2009. C. jejuni was present in 72.4% of the samples; non-jejuni/coli species made up 27.4% of the Campylobacter found in the Irish fecal samples. C. ureolyticus, C. fetus, C. upsaliensis, and C. lari were present at 24.4%, 2.4%, 1.2%, and 0.6%, respectively. The incidence of infections caused by emerging Campylobacter species may be vastly underestimated due to the lack of optimal methods for detection and isolation of these organisms (Lastovica, 2006).
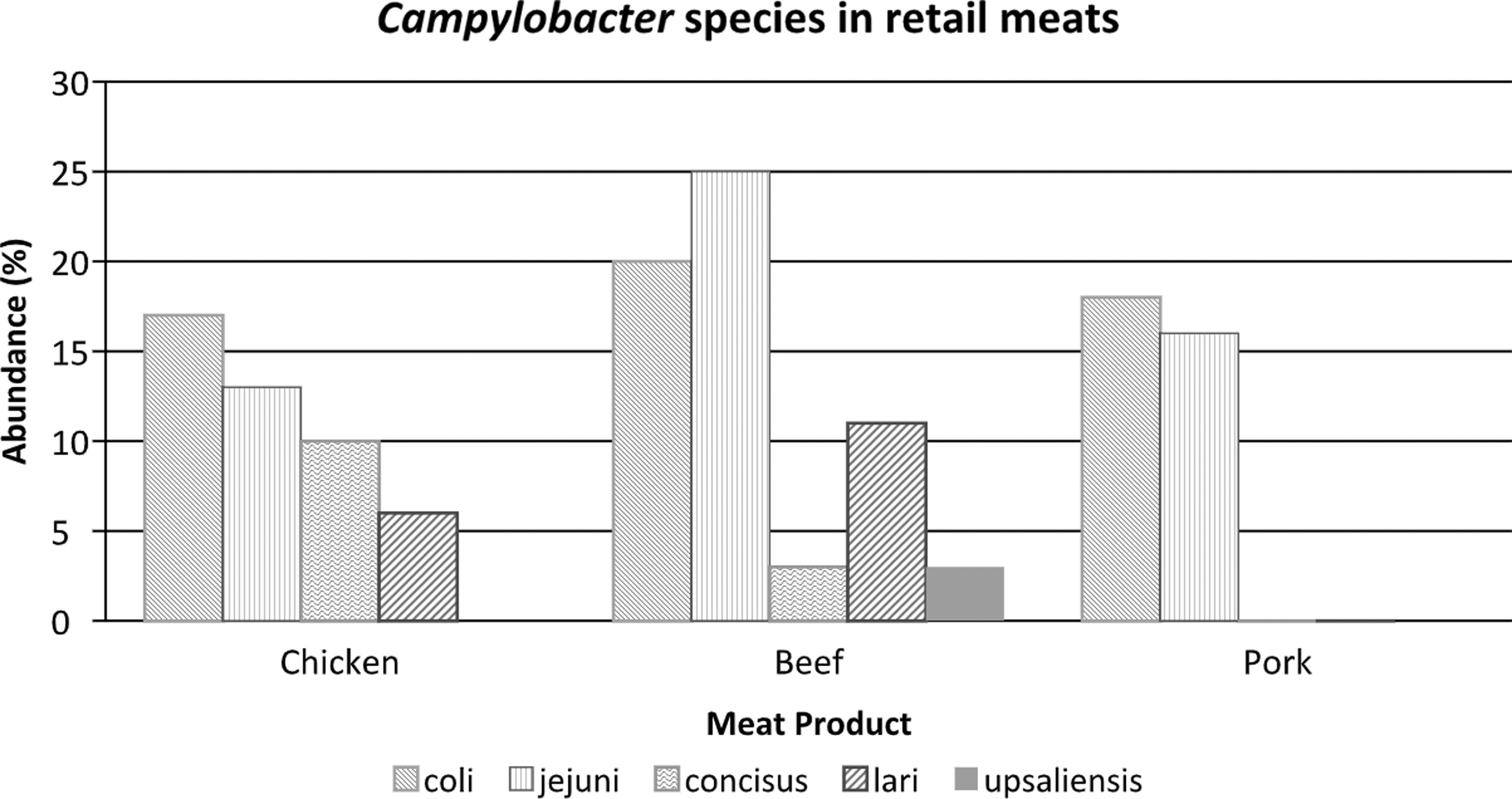
Percent frequency of Campylobacter species in raw retail meats from markets (from data from Lynch et al., 2011).
Arcobacter Species
Arcobacter species are Gram-negative spiral-shaped organisms that grow either microaerobically or aerobically. Hsu and Lee (2015) reviewed the global distribution and prevalence of Arcobacter spp. in food and water, and they concluded that cattle, dairy products, and sewage are the main source of contamination in farm animals, food, and water. The organisms are considered to be emerging foodborne pathogens (Lehner et al., 2005; Houf and Stephan, 2007; Kayman et al., 2012). Arcobacter species have been associated with diarrhea, as well as with traveler's diarrhea in individuals visiting developing countries (Collado and Figueras, 2011). For the period of 2008 to 2013, Arcobacter species were the fourth most common organisms isolated from fecal samples from patients with enteritis in Ghent, Belgium (Van den Abeels et al., 2014).
Slaughter house fecal samples indicated the presence of Arcobacter in 43.9% of pigs, 39.2% of cattle, and 16.1% of sheep in Belgium (Van Driessche et al., 2003). German slaughter house samples indicated that 43.0% of chicken broilers and 18.2% of turkey feces were positive for Arcobacter spp. (Atanassova et al., 2008), and they have been found in wild animals and companion animals (Collado and Figueras, 2011), as well as in retail meat products and seafood (Lehner et al., 2005; Collado and Figueras, 2011). In 2008, an outbreak of A. butzleri-induced infection occurred at a wedding reception in Wisconsin (Lappi et al., 2013). Fifty-one individuals were affected with diarrhea; eating broasted chicken served at the reception was associated with illness. Outbreaks due to Arcobacter species are rare, and sporadic cases are difficult to detect. While Arcobacter appears to be common in the intestinal tract of food animals and retail meat products, the importance of the organism as a foodborne pathogen is still not clear.
Helicobacter pylori and non-pylori Helicobacter Species
Helicobacter are Gram-negative microaerophilic bacteria. H. pylori is a world-wide colonizer of the human gastric mucosa, and most infected individuals are asymptomatic; however, the organism can cause gastritis, peptic ulcer, and gastric cancer (Meng et al., 2008; Vale and Vitor, 2010). The presence of H. pylori has been demonstrated in various types of water (river, waste, drinking) and in milk (raw goat, sheep, cow milk) samples (Vale and Vitor, 2010). Using multiplex polymerase chain reaction, Meng et al. (2008) detected H. pylori in 4/11 (36.4%) of fresh retail raw chicken and in 8/18 (44.4%) of ready-to-eat raw tuna from a restaurant.
The non-pylori Helicobacter species, including Helicobacter cinaedi, Helicobacter fennelliae, Helicobacter pullorum, Helicobacter canadensis, Helicobacter canis, Helicobacter winghamensis, and others have been associated with gastroenteritis and are considered to be zoonotic organisms (O'Rourke et al., 2001; Solnick, 2003). H. canadensis was implicated in cases of diarrhea in Canada and has been described as an emerging pathogen (Fox et al., 2000; Loman et al., 2009). Since many of these non-pylori helicobacters have been associated with animals, it is conceivable that they can be foodborne and considered emerging pathogens.
Streptococcus suis
Streptococcus suis causes meningitis, septicemia, endocarditis, septic shock, and arthritis in both pigs and humans. The organism is a Gram-positive facultatively anaerobic coccus (Lun et al., 2007). There are 35 serotypes based on the differences in capsular antigens; the most pathogenic strains for humans belong to serotype 2. Multilocus sequence typing indicates that serotype 2 strains can be divided into 16 sequence types (STs) (Fittipaldi et al., 2011). In Canada and the United States, 51% of serotype 2 strains belong to ST28, 44% are ST25, and 5% are ST1. There was 100% survival at 10 d when an ST28 strain was inoculated into a mouse model, 40–50% survival with ST25 strains, and 10% survival at 10 d with an ST1 strain (Fittipaldi et al., 2011). The ST1 strains are highly virulent and are responsible for a large proportion of human infections worldwide.
S. suis is generally an occupational disease of pig breeders and workers in slaughter houses, and pork processing or consuming undercooked pork are also risk factors (Fongcom et al., 2001). Data indicate that more than 85% of the cases of S. suis occur in Asian countries (Wertheim et al., 2009); however, Feng et al. (2014) stated that many sporadic cases occur in North America and are not reported. A 69-year-old swine farmer (raised ca. 2000 pigs) from Ontario, Canada was diagnosed with severe S. suis serotype 2 infection with eventual recovery (Gomez-Torres et al., 2017). Only eight human cases of S. suis infection have been reported in the United States and Canada, and the authors suggested that in North America, S. suis infections are missed or misidentified as another species. In a U.S. study of 73 individuals who were regularly exposed to swine, Smith et al. (2008) found that 7 (9.6%) had antibodies against S. suis serotype 2.
Cronobacter Species
Hunter and Bean (2013) and Jaradat et al. (2014) stated that Cronobacter species are emerging opportunistic Gram-negative pathogenic bacilli; there are seven species. These organisms have been implicated in infections, such as meningitis, necrotizing enterocolitis, and bacteremia, in newborns and infants; however, immunocompromised adults and the elderly are also susceptible to Cronobacter infections. Cronobacter has been isolated from the environment, as well as from a large number of plant and animal food products (Jaradat et al., 2014). Importantly, powdered infant formula is a major source of Cronobacter (specifically, Cronobacter sakazakii) that infect infants (Jaradat et al., 2014). From 2003 to 2009, 544 cases of Cronobacter infections were found in 6 states in the United States (Patrick et al., 2014), with a patient median age of 59 (range 1 d to 100 years). The overall incidence of Cronobacter infections was 0.66/100,000 population. It is often believed that Cronobacter infections are more likely to occur in very young children; however, the data of Patrick et al. (2014) indicated that the elderly are more at risk than infants, and clinicians should be aware of this fact.
Aeromonas Species
Members of the genus Aeromonas are important Gram-negative pathogens of fish and other cold-blooded species; however, they are also etiologic agents responsible for infections in immunocompromised, as well as immunocompetent humans. There are 24 species, and the pathogens are ubiquitous in the environment, water, foods, domestic animals, birds, and invertebrates. Aeromonas hydrophila, Aeromonas caviae, and Aeromonas veronii bv. Sobria are responsible for ≥85% of human infections (Janda and Abbott, 2010). Aeromonas species have been found in seafood, raw milk, and meat from chicken, lamb, veal, pork, and ground beef (Janda and Abbott, 2010). They have been associated with gastroenteritis, septicemia, skin and soft tissue infections, peritonitis, infections of hepatobiliary and pancreatic systems, respiratory tract infections, urogenital tract infections, and eye infections. Aeromonads have been recovered from the gastrointestinal tract and have caused gastroenteritis (Vasaikar et al., 2002); however, their role as diarrheal agents warrants further investigation. No reports of outbreaks have been published that unequivocally implicate the organisms as diarrheic pathogens (Janda and Abbott, 2010).
Cyclospora cayetanensis
Cyclospora species are microscopic unicellular parasites that cause intestinal disease in humans and animals. Humans are the only hosts of Cyclospora cayetanensis. Symptoms in humans include nausea, flatulence, fatigue, abdominal cramping, long-term watery diarrhea, low-grade fever, and weight loss. Illness is more severe in the elderly, immunocompromised individuals, and young children (Ortega and Sanchez, 2010; Chacin-Bonilla, 2017). The protozoan parasite is a fecal–oral pathogen; the oocysts from feces must mature in the environment (sewage, soil, water) for several days to become infectious. Transmission is primarily through ingestion of fruit, vegetables, or fecally contaminated water. Globalization of the food supply, increased fresh food consumption, human migration, and increased world travel has led to increased infections with C. cayetanensis (Chacin-Bonilla, 2017).
The data presented in Table 5 indicate that 33 outbreaks of foodborne Cyclospora infections have been reported for the years 2000 through 2015. Berries, snow peas, basil, and cilantro were reported as foods responsible for cyclosporiasis in the United States; most cases appear to be nonoutbreak associated. For example, for the year 2017, 1065 cases of laboratory-confirmed cyclosporiasis were reported (
Foodborne Cyclospora Infection Outbreaks in the United States: 2000–2016 (from CDC, 2017a)
Blank spaces indicate that responsible food source could not be determined.
Gnathostoma Species
The genus Gnathostoma consists of 12 species, and 4 species have been found in humans: Gnathostoma spinigerum, Gnathostoma hispidum, Gnathostoma doloresi, and Gnathostoma nipponicum (Herman and Chodini, 2009). Gnathostomiasis is a foodborne parasitic infection caused by the helminth Gnathostoma. It is a disease of tropical or subtropical regions and is endemic in areas where raw freshwater shellfish is ingested (Herman and Chiodini, 2009). There are several manifestations of gnathosomiasis, including development of cutaneous and visceral lesions. Visceral disease can include pulmonary, gastrointestinal, genitourinary, ocular, auricular, and central nervous system manifestations (Herman and Chiodini, 2009).
The life cycle of Gnathostoma is complex. In the definitive host (felines, canines, pigs), the adult worm releases eggs that are passed in the feces. In fresh water, the eggs hatch and release first-stage larvae that are ingested by copepods (small crustaceans; first intermediate host) where they develop into second-stage larvae. Ingestion of the copepods by the second intermediate host (fish, eels, frogs, birds, and reptiles) leads to second-stage larvae development into third-stage larvae in the host intestines followed by entry into muscle tissue of the second intermediate host. When the second intermediate host is eaten by a definitive host, the third-stage larvae develop into adults to continue the cycle. Humans are not definitive hosts but may be infected by the third-stage larvae (Herman and Chiodini, 2009) (
With the increase in foreign travel to tropical and semitropical countries, there is an increase in the possibility of eating foods that contain Gnasthostoma species. Data seem to be lacking concerning the incidence of gnathostomiasis in the United States. In the recent literature, two cases of gnathostomiasis have been reported for the United States (Catalano et al., 2009; Jarell et al., 2011). Either gnathostomiasis is rare in the United States or cases are under- or misdiagnosed.
In various areas of Asia, swamp eels (wild-caught or aquaculture-reared) are consumed as food. Swamp eels are a common source of third-stage larvae of Gnathostoma species. Live swamp eels are shipped to North American markets from Asia for the ethnic trade. Cole et al. (2014) found that 13/47 (27.7%) swamp eels (from three markets in New York, one market in Orlando, FL, and one market in Atlanta, GA) were positive for Gnathostoma (27.7%). In addition, Cole et al. (2014) determined the incidence of Gnathostoma in wild populations of swamp eels in waters of Florida and New Jersey; of 67 eels, 3 were positive (4.5%). The U.S. swamp eels represented populations of Asiatic eels released over the decades in which live eels have been imported. Gnathostoma present in wild swamp eels released in the waters of the United States may present a potential problem in the future since the environment contains all of the components necessary for the completion of the life cycle of the helminth. Furthermore, there is an increase in the exotic culinary tastes of U.S. residents and travelers. Strategies for the prevention of gnathostomiasis include educating consumers that meat from animal species that may be infected with Gnathostoma should be thoroughly cooked, and individuals with migratory subcutaneous swellings must have a medical evaluation (Diaz, 2015).
FoodBorne Viruses
The data presented in Table 6 indicate that foodborne outbreaks due to viral contamination of food in the United States are rare, except for norovirus outbreaks. Norovirus is the major cause of foodborne outbreaks and illnesses in the United States (CDC, 2011, 2013b, 2014a, 2014b, 2015). The major cause of foodborne outbreaks due to norovirus is the mishandling of food by infected workers. Mattison et al. (2007) have suggested that norovirus is a zoonotic agent since they identified human norovirus RNA sequences in 4/120 (3.3%) swine feces and 1/179 (0.006%) bovine feces, as well as in 1/156 retail raw meat (the positive sample was pork). However, others disagree with the zoonotic status of norovirus (Bank-Wolf et al., 2010).
Reported Viral Foodborne Outbreaks, United States 2008–2012
Includes both confirmed and suspected organisms.
Group A and C rotaviruses have been shown to be zoonotic viruses and are transmitted by the fecal–oral route through person-to-person contact or through food contaminated by an infected food handler (Martella et al., 2010). Astroviruses and rotaviruses infect a large number of mammals and birds (De Benedictis et al., 2011). Historically, astroviruses have been considered to be species-specific, but the fact that new astroviruses are regularly found in humans and animals suggests that cross-species infection and adaptation to new hosts may be common; however, more information is needed to demonstrate that astroviruses as zoonotic organisms. Astroviruses are also transmitted by person-to-person contact or through food contaminated by infected food handlers. Data presented in Table 6 indicate that astrovirus and rotavirus are not very common as causes of foodborne outbreaks. Scallan et al. (2011) estimate that less than 1% of astrovirus, rotavirus, and sapovirus infections are foodborne, whereas hepatitis A virus and norovirus are responsible for 7% and 26%, respectively, of foodborne viral infections (Table 7).
Estimated Annual Number of Illnesses, Hospitalizations, and Deaths Due to Foodborne Viruses, United States (Data from Scallan Et al., 2011)
Hepatitis E virus (HEV) is a virus in the genus Hepevirus of the Hepeviridae family. It is a nonenveloped virus with a positive-sense, single-stranded 7.2-kb RNA genome with three open reading frames. There is only one serotype of HEV, but there are four major genotypes (HEV1, HEV2, HEV3, and HEV4) (Kamar et al., 2014). HEV1 and HEV2 are mainly seen in developing countries and are generally transmitted by water contaminated by human feces; travelers to developing countries can contact HEV from contaminated water sources. HEV1 and HEV2 infect only humans. In developed countries, HEV3 (genotype 3 has several subtypes) and HEV4 are transmitted to humans from animal reservoirs such as swine and deer.
The domestic pig is the major reservoir of HEV (Kamar et al., 2014). The HEV IgG seroprevalence of 4742 domestic pigs from 15 countries of North America, South America, Europe, and Asia was 49.8% (range 6.0–85.5%) (Vasickova et al., 2007). Teshale et al. (2010) have designated the United States as a country where HEV is endemic in the swine population. Sporadic cases or small clusters of cases of HEV infection have been reported in European countries, and in developed countries of the Asia-Pacific region (Ruggeri et al., 2013); however, few reports are available for the United States (Dalton et al., 2008). A recent report indicates that HEV infections are increasing in European countries (Kupferschmidt, 2016). Blood and blood products (used in transfusions) account for part of the increase, but a greater proportion of the European HEV infections is due to the ingestion of pork sausages and other pork products. Analysis of 919 French patients infected with genotype 3 HEV showed that 5 were infected with HEV3-ra (rabbit HEV). The virus has been found in farmed and wild rabbits. None of the five patients had contact with rabbits suggesting that HEV-ra infection in these five patients was either food or waterborne. (Abravanel et al., 2017).
HEV infection in humans causes acute liver disease with clinical signs in symptomatic patients similar to those of hepatitis A infection and includes hepatomegaly, anorexia (loss of appetite), myalgia (muscle pain), arthralgia (joint pain), jaundice, abdominal pain, nausea, vomiting, diarrhea, and fever. In the majority of patients, the hepatitis disease is self-limiting and lasts a few weeks. More rarely, HEV infection can induce extrahepatic syndromes, such as neurological disorders, renal disorders, pancreatitis, and hematological disorders. In immunocompromised individuals, HEV infection may progress to a chronic disease with liver cirrhosis and persistent viral shedding (Yugo and Meng, 2013; Kamar et al., 2014). Fatalities for the acute disease are in the range of 0.5–4%; however, pregnant women may undergo liver failure with 20–30% mortality (Song et al., 2014).
In industrialized countries, the incidence of clinical HEV is low in humans; however, the seroprevalence is high, indicating the occurrence of subclinical or underdiagnosed illnesses. Most of the studies involving seroprevalence to HEV have been done with blood donors. Blood donors from Switzerland showed a prevalence of HEV IgG of 4.9% (27/550) (Kaufmann et al., 2011), whereas, 52.3% (268/512) of French blood donors were HEV IgG positive (Mansuy et al., 2011). Blood donors from Austria were 14.3% positive for HEV IgG (Lagler et al., 2014). Slot et al. (2013) found that 1401/5239 (26.7%) of Dutch blood donors were positive for HEV IgG, and 49/5239 (0.93%) were IgM positive; HEV RNA was detected in 4 of the 49 IgM-positive sera (0.08%). Similar studies were conducted in Germany and the United States (Xu et al., 2013; Juhl et al., 2014). Juhl et al. (2014) found that 6.8% of individuals were HEV IgG positive, and Xu et al. (2013) found 18.8% positive individuals in the United States.
There is limited information on the incidence of clinical HEV in developed countries, but the estimated incidence in the general population of the United Kingdom is 2/1000 (Kamar et al., 2014) and 7/1000 for the United States (Faramawi et al., 2011). Foodborne HEV infections have been detected in the United Kingdom (Said et al., 2014). Tohme et al. (2011) reported two indigenous cases of HEV infection that occurred in Texas in 2009, and genotype 3a was isolated from one patient. Since the virus was closely related to U.S. swine HEV, Tohme et al. (2011) proposed that the illness could have been foodborne.
HEV RNA has been identified in a number of animals, including swine, chickens, deer, rats, wild boar, monkeys, mongoose, rabbits, ferrets, and bats. The only animals that act as reservoirs associated with HEV infections in humans are domestic pigs, wild boars, and deer. Meat products from those animals have been shown to induce hepatitis (Pavio et al., 2010; Van der Poel, 2014). Pork products from processing plants or from grocery stores can harbor HEV. For example, Berto et al. (2012) found that 6/63 (9.5%) of pork sausages obtained from retail sources in the United Kingdom were positive for HEV RNA. Consumption of pork pie, ham, and sausages purchased from U.K. supermarkets was significantly correlated with HEV infection in 17/25 (68.0%) patients (Said et al., 2014). HEV RNA was found in a number of French food products containing raw pork liver (Pavio et al., 2014). Ingestion of raw pig liver sausages (figatelli) induced HEV infection in members of a French family; the HEV genotype was 3f (Renou et al., 2014). Feagins et al. (2007) obtained from 127 packages of commercial pig livers from grocery stores in Virginia and demonstrated that 14 (11.0%) were positive for genotype 3 HEV RNA. They also demonstrated that homogenates from positive livers, when inoculated into specific pathogen-free pigs, induced HEV infection, indicating that the livers contained viable infectious virus.
Conclusions
The importance of emerging foodborne pathogens will likely continue to increase as a global issue. Increased research, surveillance, and vigilance are needed to understand the emergence and spread of pathogens in the food supply. Furthermore, risk assessments are important to identify and characterize foodborne hazards, and technologies for accurate detection of emerging pathogens and development of control interventions from farm to table are critical. Strong partnerships among government agencies (both domestic and foreign public health agencies), as well as health care, pharmaceutical, diagnostic, and food industries are needed to decrease the incidence of emerging foodborne infections through safe food production and acceleration of the public health response to foodborne illness at the local, national, and global level.
Disclaimer
Mention of trade names or commercial products is solely for the purpose of providing specific information and does not imply recommendation or endorsement by the U.S. Department of Agriculture.
Footnotes
Disclosure Statement
No competing financial interests exist.