
Abstract
This study characterized the prevalence and antimicrobial resistance characteristics of foodborne Salmonella isolates from March 2016 to February 2017 in Shanghai, China. A total of 147 (14.2%) nonduplicate foodborne Salmonella isolates were obtained from 1035 food samples. The Salmonella isolates were most frequently identified in fresh meat samples (28.0%), followed by ready-to-eat foods (9.0%), frozen convenience foods (7.1%), and fresh produce (4.5%). The top 3 serovars were Salmonella Enteritidis (46.3%; 68/147), Salmonella Typhimurium (32.7%; 48/147), and Salmonella Derby (6.8%; 10/147). The majority of isolates were resistant to sulfisoxazole (93.9%; 138/147) and trimethoprim/sulfamethoxazole (61.2%; 90/147). Interestingly, frozen convenience food isolates exhibited an extremely high multidrug resistance rate (86.7%; resistant to ≥3 classes of antimicrobials). Among 81 quinolone-resistant isolates, aac(6′)-Ib-cr (100%), oqxAB (84.0%), qnrS1 (23.5%), D87Y (49.4%), and D87N (33.3%) mutations in GyrA, and T57S in ParC (12.3%) were observed. The β-lactamase genes bla TEM-1 (100%) were present in 63 ampicillin-resistant isolates. Polymerase chain reaction-based plasmid replicon typing revealed that 147 isolates represented 6 plasmid incompatibility groups (IncFIIs, IncHI2, IncI1, IncP, IncFIC, and IncA/C), among which, IncFIIs (59.2%) and IncHI2 (26.5%) were predominant. The genetic relationship of isolates was elucidated using multilocus sequence typing (MLST) and pulsed-field gel electrophoresis (PFGE). MLST results indicated that ST34 and ST11 were predominate types in Salmonella Typhimurium (56.3%; 27/48) and Salmonella Enteritidis (95.6%; 65/68), respectively. Importantly, 96.3% (26/27) of ST34 Salmonella Typhimurium isolates possessed the ACSSuT resistance pattern (ampicillin, chloramphenicol, streptomycin, sulfamethoxazole, and tetracycline). PFGE analysis of ST34 isolates showed clonal dissemination across all four types of retail foods. Our findings highlight the high prevalence of antimicrobial-resistant Salmonella isolates in retail foods in Shanghai, especially the clonal expansion of ST34 isolates with MDR-ACSSuT resistance, which might pose a public health threat.
Introduction
Salmonella enterica is an important foodborne pathogen that is implicated in ∼94 million illnesses and 155,000 deaths worldwide annually (Eng et al., 2015). Salmonella is a common cause of human bacterial gastroenteritis, and an increasing trend in the isolation rate of Salmonella from diarrheal patients from 2013 to 2016 was presented by Chinese foodborne disease surveillance systems (Liu et al., 2018). Previous studies have shown that the majority of the human salmonellosis was linked to the ingestion of contaminated foods, including meats and fresh produce (Jackson et al., 2013; Chai et al., 2017). Recently, Salmonella outbreaks associated with ready-to-eat and frozen foods have been also reported (Jones et al., 2016; Vestrheim et al., 2016; Huusko et al., 2017). Increased consumption of these foods has given rise to the high risk of Salmonella infection.
The inappropriate use of antimicrobials in clinics and agriculture has resulted in the emergence and spread of antimicrobial resistance among Salmonella isolates (Klein et al., 2018). An increase of multidrug-resistant (MDR) Salmonella in food has been described in recent years (Li et al., 2014; Hu et al., 2017; Ren et al., 2017). Given these reports, it is of great importance to protect the public from Salmonella infections, and the data on the prevalence and antimicrobial resistance of Salmonella in various foodstuffs could be used to strengthen overall food safety regulations.
Multilocus sequence typing (MLST) and pulsed-field gel electrophoresis (PFGE) are standardized methods for studying the evolution of Salmonella (Ma et al., 2018). Previous studies have uncovered an association between sequence types (STs) and antimicrobial resistance phenotypes (Sun et al., 2014; Cao et al., 2018). For instance, the ACSSuT phenotype representing the combined ampicillin, chloramphenicol, streptomycin, sulfamethoxazole, and tetracycline resistance was commonly associated with ST34 (Mulvey et al., 2013; Sun et al., 2014).
The objective of our study was to investigate the prevalence and to characterize antimicrobial resistance of Salmonella isolates recovered from retail foods in Shanghai, China. The relatedness between antimicrobial phenotypes and ST was determined using MLST profiles, and genetic relationships among ST34 isolates were further analyzed using PFGE.
Materials and Methods
Salmonella isolation and identification
A total of 1035 samples of retail foods from markets in 8 districts of Shanghai between March 2016 and February 2017 were collected. Samples included fresh meat (n = 361, including pork, chicken, beef, duck, and mutton), fresh produce (n = 243, including banana, grape, strawberry, tangerine, cabbage, celery, cauliflower, lettuce, carrot, lotus root, eggplant, cucumber, tomato, and pepper), ready-to-eat foods (n = 221, including prepacked milk, yogurt, milk beverage, salad, ham sausage, spicy strip, steamed stuffed bun, deep-fried dough stick, wonton, and ready-to-eat eggs), and frozen convenience foods (n = 210, including stuffed bun, dumpling, meatball, sausage, and spring roll). The detailed sampling information is shown in Supplementary Table S1.
Sample collection and Salmonella isolation were performed as previously described with some modifications (Abatcha et al., 2018). Briefly, 25 mL (or 25 g) samples were cultured in 225 mL of buffered peptone broth and were incubated for 24 h at 37°C. Samples of these pre-enriched cultures (0.1 mL) were incubated in 9.9 mL of Rappaport Vassiliadis Soya (RVS) broth for 24 h at 42°C. RVS cultures were then streaked onto xylose lysine deoxycholate agar, and incubated for 24 h at 37°C. Suspected Salmonella isolates were identified using API20E test strips (BioMerieux, France) and confirmed by polymerase chain reaction (PCR) amplification using invA as the target gene (Cui et al., 2006). The primers are listed in Supplementary Table S2. PCR was performed using the following conditions: 94°C for 10 min, followed by 30 cycles of 94°C for 30 s, 64°C for 30 s, 72°C for 30 s, and 72°C for 7 min.
Salmonella isolates were serotyped with commercial antiserum (Statens Serum Institute, Copenhagen, Denmark) according to the manufacturer's guidelines.
Antimicrobial susceptibility tests
Seventeen antimicrobials were selected based on World Health Organization ranking of antimicrobials according to their importance in human medicine (Collignon et al., 2016). Antimicrobial susceptibilities were determined using the agar dilution method and interpreted according to CLSI guidelines (CLSI, 2018). Seventeen antimicrobials were examined that included ampicillin (AMP), cefepime (CPE), ceftriaxone (CRO), imipenem (IMI), fosfomycin (FOS), nalidixic acid (NAL), ofloxacin (OFX), ciprofloxacin (CIP), gentamicin (GEN), amikacin (AMK), kanamycin (KAN), chloramphenicol (CHL), tetracycline (TET), azithromycin (AZM), trimethoprim/sulfamethoxazole (SXT), streptomycin (STR), and sulfisoxazole (SUL).
Salmonella isolates showing resistance to at least three classes of antimicrobials were defined as MDR. Escherichia coli ATCC 25922 was used as the quality control strain.
Determination of antimicrobial resistance genes
The presence of antimicrobial resistance genes (ARG) in Salmonella isolates resistant to each class of antimicrobial, including β-lactamases (bla TEM and bla CTX-M), quinolones [qnrA, qnrB, qnrS qepA, oqxAB, and aac(6′)-Ib-cr], aminoglycoside [aadA and aac(6′)-Ib], tetracycline (tetA, tetB, tetC, and tetG), and chloramphenicol (floR and cmlA) was screened by PCR. Quinolone resistance-determining region (QRDR) genes gyrA, gyrB, parC, and parE were amplified using PCR as previously described (Chen et al., 2007). All primers used are listed in Supplementary Table S2.
PCR products were purified from gels after electrophoresis using a commercial kit (Takara, Dalian, China) and sequenced by Shanghai Sunny Biotechnology (Shanghai, China). QRDR mutations were determined using the Basic Local Alignment Search Tool (
Molecular typing
MLST analysis was conducted by sequencing aroC, dnaN, hemD, hisD, purE, sucA, and thrA as previously described (Ma et al., 2018). The sequence data acquired of each gene were imported into Salmonella enterica MLST database (
Plasmid replicon typing
Plasmid incompatibility (Inc) groups were assigned using PCR-based replicon typing with genomic DNA samples of 147 isolates as templates. Each isolate was examined for the presence of 18 plasmid replicons using 5 different multiplex-PCR and 3 simplex-PCR assays as described previously (Carattoli et al., 2005).
Results
Prevalence of Salmonella isolates in food samples
Overall, 147 out of 1035 (14.2%) food samples were found to be positive for Salmonella. Nine serovars were identified, including Enteritidis (n = 68), Typhimurium (n = 48), Derby (n = 10), Newport (n = 6), Agona (n = 4), Choleraesuis (n = 4), Senftenberg (n = 3), Aberdeen (n = 2), and Thompson (n = 2). The detailed information of 147 Salmonella isolates is listed in Supplementary Table S3.
Table 1 shows the prevalence of Salmonella isolates in various retail food types. Of the 361 fresh meat samples, 101 (28.0%) were positive for Salmonella, including 46/143 (32.2%) of pork samples, 28/102 (27.5%) of chicken samples, 16/66 (24.2%) of beef samples, 6/25 (24.0%) of duck samples and 5/25 (20.0%) of mutton samples. The prevalence (4.5%; 11/243) of Salmonella in ready-to-eat foods and frozen convenience foods was 9.0% (20/221) and 7.1% (15/210), respectively. The lowest prevalence (4.5%; 11/243) of Salmonella was observed in fresh produce.
Prevalence of Salmonella spp. in Different Retail Food Sources
Antimicrobial resistance profiles
Antimicrobial susceptibilities of 147 isolates are exhibited in Supplementary Table S4. The Salmonella isolates displayed high resistance to sulfisoxazole (93.9%; 138/147), trimethoprim/sulfamethoxazole (61.2%; 90/147), nalidixic acid (49.7%; 73/147), streptomycin (46.3%; 68/147), and ampicillin (42.9%; 63/147), with lower rates of resistance to tetracycline (25.9%; 38/147), ciprofloxacin (25.2%; 37/147), chloramphenicol (21.8%; 32/147), gentamicin (18.4%; 27/147), kanamycin (18.4%; 27/147), ofloxacin (15.6%; 23/147), and fosfomycin (2.0%; 3/147). All the isolates were susceptible to cefepime, ceftriaxone, amikacin, and imipenem.
Overall, 70 of 147 (47.6%) Salmonella were MDR, and 31 of them were resistant to at least 5 classes of antimicrobials. The most common MDR profiles observed were AMP-STR-SUL-NAL-SXT (n = 21) in Salmonella Enteritidis and ACSSuT-GEN-KAN-NAL-OFX-CIP-SXT (n = 9) in Salmonella Typhimurium (Supplementary Table S5). It is notable that isolates from frozen convenience foods showed the highest rate of multidrug resistance (86.7%; 13/15). Lower resistance rates were found in the isolates from ready-to-eat foods (70.0%; 14/20), fresh produce (54.5%; 6/11), and fresh meat (36.6%; 37/101), respectively (Table 2).
Prevalence of Antimicrobial Resistance in 147 Salmonella Isolates
MDR, multidrug resistant.
ARG prevalence
The 63 ampicillin-resistant isolates possessed bla TEM-1, while bla CTX-M was not found. All 32 chloramphenicol-resistant isolates carried floR and cmlA. Genes aac(6′)-Ib (100%) and aadA (95.7%) were present in 70 aminoglycoside-resistant isolates. The genes tetA (97.4%), tetB (97.4%), and tetC (15.8%) were identified in 38 tetracycline-resistant isolates, while tetG was not found. Plasmid-mediated quinolone resistance (PMQR) determinants that included qnrS1 (23.5%), aac(6′)-Ib-cr (100%), and oqxAB (84.0%) were confirmed in 81 quinolone-resistant isolates, while qnrA, qnrB, or qepA was not found (Table 3).
Genotypic Characteristics of 140 Antimicrobial-Resistant Salmonella Isolates
QRDR mutations were identified in 73 of 81 quinolone-resistant isolates. Single mutations were present in GyrA in 63 isolates that included D87Y (42.0%; 34/81), D87N (33.3%; 27/81), and D87G (2.5%; 2/81). A single mutation T57S in ParC (4.9%; 4/81) was also identified. Double QRDR mutations were found in six isolates and these were D87Y in GyrA and T57S in ParC. All isolates possessed no mutation in GyrB and ParE. Notably, all Salmonella Typhimurium and Salmonella Enteritidis isolates carried the GyrA mutations D87N and D87Y, respectively (Table 4).
Quinolone Resistance-Determining Region Mutations in 81 Quinolone-Resistant Salmonella Isolates
Molecular subtypes
The 147 Salmonella isolates were grouped into 14 STs. ST11 (65/68) predominated in Salmonella Enteritidis isolates, while the remainder were subtyped into ST1925 (3/68). A variety of STs were identified in the Salmonella Typhimurium isolates, which included ST34 (27/48), ST19 (11/48), ST1544 (8/48), ST36 (1/48), and ST1936 (1/48). Interestingly, the remaining STs were serovar associated as follows: ST40 (Salmonella Derby), ST14 (Salmonella Senftenberg), ST13 (Salmonella Agona), ST26 (Salmonella Thompson), ST33 (Salmonella Newport), ST1583 (Salmonella Choleraesuis), and ST426 (Salmonella Aberdeen).
All ST34 of Salmonella Typhimurium were MDR isolates that encompassed seven resistance profiles, while the remaining STs (19, 1544, 36, and 1936) possessed less or no resistance (Supplementary Table S5). This result indicated an association between resistance phenotype and ST in Salmonella Typhimurium. To analyze this in more detail, the genetic similarity and evolutionary relationships among the ST34 isolates were determined by PFGE subtyping. A total of five pulsotypes were identified (Figure 1). Sixteen isolates from different foods in different areas belonging to Group C shared 100% similarity, and antimicrobial resistance phenotypes and genes of which were highly similar, suggesting the clonal expansion of ST34 isolates in different retail foods in Shanghai.
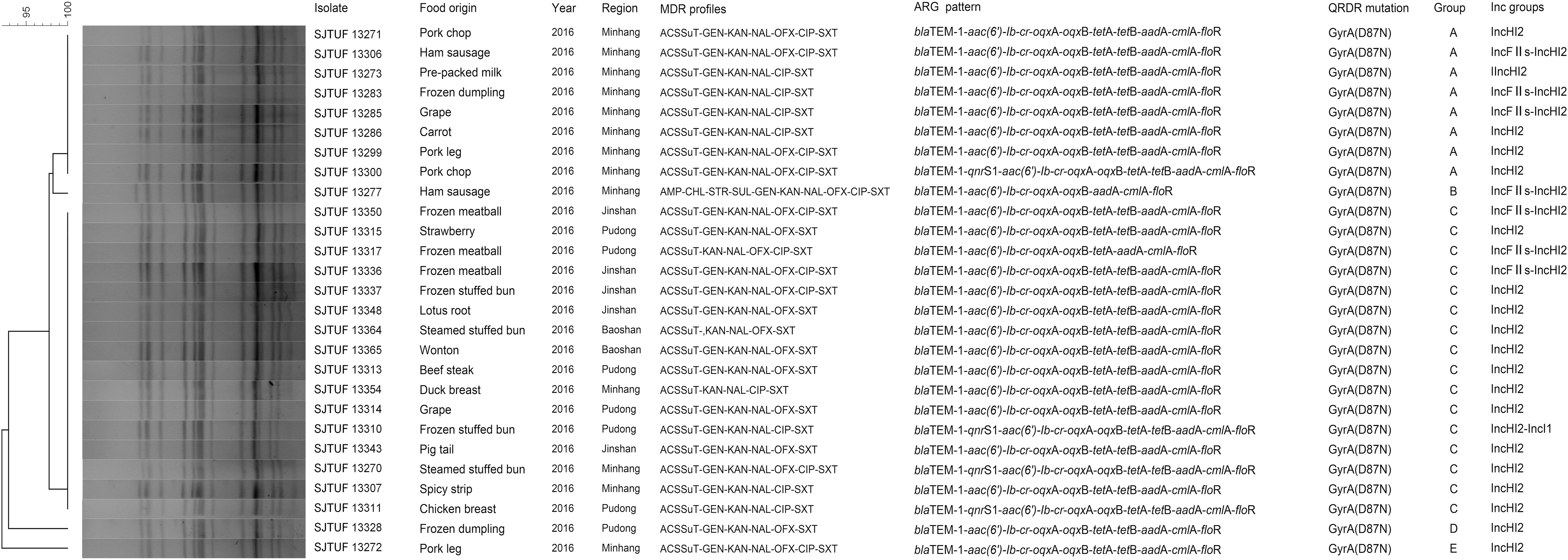
XbaI PFGE patterns of 27 ST34 Salmonella enterica isolates recovered from retail foods. PFGE, pulsed-field gel electrophoresis.
Plasmid replicon types
Six plasmid incompatibility groups were identified in the 147 isolates. The most prevalent was IncFIIs (59.2%; 87/147), followed by IncHI2 (26.5%; 39/147), IncI1 (15.0; 22/147), IncFIC (6.1%; 9/147), IncP (3.4%; 5/147), and IncA/C (1.4%; 2/147). None of the isolates harbored IncHI1, X, L/M, N, FIA, FIB, Y, W, T, FrepB, K, or B/O replicon type.
In our study, 83 isolates possessed single plasmid replicon type, including IncFIIs (37.4%; 55/147), IncHI2 (15.6%; 23/147), IncFIC (2.7%; 4/147), and IncI1 (0.7%; 1/147). Thirty-six isolates presented two Inc groups, including IncFIIs-IncI1 (8.8%; 13/147), IncFIIs-IncHI2 (8.2%; 12/147), IncI1-IncP (2.7%; 4/147), IncFIIs-IncFIC (2.0%; 3/147), IncHI2-IncI1 (1.4%; 2/147), IncFIIs-IncA/C (0.7%; 1/147), and IncP-IncFIC (0.7%; 1/147). Three isolates carried the combination of three replicon types (IncFIIs-IncHI2-IncFIC, IncFIIs-IncHI2-IncI1, and IncFIIs-IncI1-IncA/C). Notably, Salmonella Enteritidis and Salmonella Typhimurium have shown the preference to certain Inc groups. IncFIIs plasmid was predominant in Salmonella Enteritidis, while IncHI2 was the most prevalent in Salmonella Typhimurium (Table 5).
Plasmid Replicon Types of 147 Salmonella Isolates
Discussion
In recent years, an increase of antimicrobial-resistant Salmonella in retail foods has been reported (Hu et al., 2017; Ma et al., 2018). In this study, we analyzed the prevalence and antimicrobial resistance characteristics of foodborne Salmonella isolates in Shanghai.
The contamination rate of Salmonella spp. in retail foods in our study in Shanghai from 2016 to 2017 was 14.2%. Fresh meat samples possessed the highest prevalence at 28.0%, which was significantly higher than that in raw meat category collected during the period of 2006–2016 in China (5.3%; Narayan et al., 2018). Compared to other meat types, higher isolated rates of Salmonella in pork (32.2%; 46/143) and chicken (27.5%; 28/102) were identified. Furthermore, our results were higher than those reports in north and south of Yangtze River (Zeng et al., 2019), in Shaanxi, China (Yang et al., 2010), and in Czech Republic (Myšková and Karpíšková, 2017). Besides, surveillance data of Salmonella in fresh meat samples in this study were similar to that in corresponding animal resources in China between 2003 and 2011 (Wu et al., 2018), which suggested that food animals, especially chicken and pig, were particular reservoirs of Salmonella. Therefore, sanitary control in the process of treating such meat products must be strengthened from farm to fork.
Frozen convenience foods are ready to cook and have a long shelf-life, which made it increasingly popular in China. However, Salmonella contamination in frozen convenience foods was found in our study, and 7.1% isolates were identified. The study about the prevalence of Salmonella in frozen convenience foods was rare in China, and our results were higher than that in other countries, including Estonia (Kramarenko et al., 2014) and Brazil (Medeiros et al., 2011). More importantly, frozen convenience foods might act as potential vectors for the transmission of Salmonella into retail markets, with potential hazard to human health through cross-contamination with Salmonella during the handling process.
Earlier research conducted on Chinese food commodities between 2006 and 2016 revealed that vegetables and fruits were in low risk (3.0%) of contamination by Salmonella (Narayan et al., 2018). In this study, 4.5% of fresh produce was Salmonella positive, which was much higher than in fresh produce from U.S. chain markets (0.3%) (Liu and Kilonzo-Nthenge, 2017) and Canadian marketplace (1.3%) (Denis et al., 2016). Despite the increased importance of fresh produce as vehicles for the transmission of human pathogens, there are currently limited surveillance data of Salmonella in fresh produce in China, and continuous monitoring is necessary to ensure the safety of such foods.
High prevalence (47.62%) of MDR Salmonella isolates was observed in this study (Table 2). It was of particular concern that 86.7% of frozen convenience food isolates was resistant to at least three classes of antimicrobials in this study, which was higher than that in other food sources. Furthermore, 46.7% was found to be resistant to at least five classes of antimicrobials. Compared with our study, a previous study in Iraq showed a significantly lower level (60.8%) of MDR Salmonella isolates from frozen foods (Harb et al., 2018). Frozen convenience foods is the complex of various raw materials, and its preparation generally involves more complicated processing and a long supply chain, in which food contamination with pathogens can occur at multiple steps (Elhadi, 2014; Ye et al., 2015).
Once there are antimicrobial-resistant strains in the raw materials, the mobile resistance genetic elements might be transmitted among different strains during the long food chain. This may explain the higher prevalence of MDR strains in frozen convenience foods than in other food types with a single, short supply chain. Due to the fact that foodborne pathogens could be transferred to humans through food chain (Brands et al., 2005; Zhang et al., 2018), ingestion of contaminated foods with antimicrobial-resistant pathogens may lead to more serious infections in humans, and make it difficult to be treated in clinics. Therefore, the regulation and supervision of frozen foods must be accelerated by the government departments to reduce the risk of human infections with antimicrobial-resistant pathogens.
PMQR, aad, tet, bla, floR, and cmlA, the plasmid-located ARG, generally conferred reduced susceptibility of Salmonella to quinolones, aminoglycoside, tetracycline, β-lactamase, and chloramphenicol (Blair et al., 2015; Nair et al., 2018). High detection rates of aac(6′)-Ib-cr (100%) and oqxAB (84.0%) were identified in quinolone-resistant isolates, as well as that of bla TEM-1 (100%), aadA (95.7%), tetA (97.4%), tetB (97.4%), cmlA (100%), and floR (100%) in the isolates resistant to the corresponding antimicrobial in this study. These results may account for the existence of antimicrobial resistance phenotypes of these isolates.
Plasmids have been considered to be the major driving force in the transmission of antimicrobial resistance in Salmonella isolates because they generally carried ARG (Glenn et al., 2011; Jain et al., 2018). IncFIIs and IncHI2 have been reported to be associated with multidrug resistance since both of them harbored genes encoding resistance to β-lactamases, aminoglycoside, quinolone, tetracyclines, and streptomycin (Rozwandowicz et al., 2018). In our study, IncFIIs (59.2%) was the most prevalent replicon type, which had a preference in Salmonella Enteritidis. It should be noticed that IncFIIs plasmids were commonly associated with multidrug resistance determinants such as oqxA, oqxB, floR, tetA, bla TEM-1, and other sets of ARG (Yang et al., 2016; Oliva et al., 2018).
IncHI2, another dominant replicon type (26.5%), mainly existed in Salmonella Typhimurium isolates and was known to play a pivotal role in the acquisition of antimicrobial resistance in Enterobacteriaceae (Coelho et al., 2012). The transmission of aac(6′)-Ib-cr and oqxAB in Salmonella has been considered to be linked with such plasmids (Wong et al., 2016). In addition, the combinations of different plasmid replicons (e.g., IncFIIs-IncI1 and IncFIIs-IncHI2) were found to be co-resident in MDR Salmonella isolates in this study, and similar results were presented in other studies (Poole et al., 2009; Glein et al., 2013). Given these reports, it could be speculated that the widespread antimicrobial resistance in Salmonella isolates might be due to the transfer of these plasmids harboring ARG.
The ST34 European clone is a rapidly emerging cause of global MDR salmonellosis and is currently found throughout Europe and Asia (Campos et al., 2016; Li et al., 2017). ST34 (27/48) was the major ST of MDR foodborne Salmonella Typhimurium isolates in our study and this result correlates with other investigations in Guangdong and Hong Kong (Wong et al., 2013; Sun et al., 2014). Notably, particular antimicrobial resistance profiles have been found to be ST linked. In this study, ST34 was frequently associated with the MDR-ACSSuT phenotype, which was also identified in other studies (Wong et al., 2013).
Previous studies have shown the emergence of human clinical Salmonella ST34 clones possessing high-level resistance to quinolones and ACSSuT antimicrobials (Wong et al., 2013; Lu et al., 2019). Our PFGE results of ST34 isolates demonstrated their clonal spread in different retail food types. In addition, the clonal expansion of MDR-ACSSuT ST34 between patients and foods has been reported (Ma et al., 2018). The dissemination of antimicrobial resistance in ST34 Salmonella clone has been ascribed to the acquisition of conjugative plasmids by horizontal gene transfer, and ST34 MDR Salmonella harboring ARG within IncHI2 plasmids has been reported previously (Campos et al., 2016). It is notable that all of ST34 clone in this study carried IncHI2 plasmids (Supplementary Table S3), which revealed the wide spread of IncHI2 plasmids in retail foods. As a consequence, IncHI2 plasmids should be attached great importance since they may contribute to dissemination of undesirable antimicrobial resistance among Enterobacteriaceae. Based on these findings, Salmonella ST34 isolates must be of particular concern, and further studies about evolution of ST34 isolates can be conducted in a genomic level. Continuous surveillance to contain further transmission of this plasmid for new hosts or different settings is also urgently required.
Conclusions
Our results demonstrated the extensive existence of MDR Salmonella isolates in retail foods in Shanghai. The retail foods, especially fresh meat and frozen convenience foods, might act as reservoirs of antimicrobial-resistant Salmonella. Clonal dissemination of IncHI2-positive ST34 isolates with MDR-ACSSuT resistance existed in different food sources, the presence of which was potentially dangerous in case of human infections.
Footnotes
Disclosure Statement
No competing financial conflict of interests exist.
Funding Information
This work was supported by the National Key R&D program of China (No. 2017YFC1600100) and the National Natural Science Foundation of China (No. 31601562).
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.