
Abstract
Aiming at investigating the potential sources of Campylobacter spp. contamination in pig slaughterhouse in eastern China, a total of 2056 samples were collected in pork production chain stretching from the upstream farm to the slaughterhouse, including 54 cloacal swabs from 54 live pigs on farm, 1726 samples from 214 pig carcasses along the eight main steps in the slaughtering line, and 276 samples from slaughtering environment. Campylobacter spp. were found, may be propagated in each slaughtering step, with an average prevalence of 19.3% (333/1726). The isolation rate of Campylobacter spp. in samples collected before the slaughter (42.5%, 4.87 ± 1.23 log colony-forming units [CFU]/g), dehairing (28%, 2.40 ± 0.49 log CFU/500 cm2), and evisceration (29.4%, 2.50 ± 0.54 log CFU/500 cm2) were significantly higher than other slaughter processes (p < 0.05). The prevalence of Campylobacter spp. of pigs on farm was 18.5% (10/54), compared to the slaughtering environment, which was 27.9% (77/276). Campylobacter spp. isolates were obtained from a batch of samples in slaughterhouse and farm belonged to ST-828 complex. Interventions are required to minimize Campylobacter spp. contamination in slaughtering line, especially during dehairing and evisceration. The upstream farm was an additional and usually neglected source of contamination. These findings may provide a new perspective regarding the safety provision of Campylobacter spp. contamination in pork meat production.
Introduction
Human campylobacteriosis remains an estimated 400 to 500 million cases of diarrhea worldwide, although Campylobacter infections are sporadic and self-limiting. A low infection dose of about 500 Campylobacter jejuni cells can derive the initial clinical symptoms; campylobacteriosis is characterized by watery and sometimes bloody diarrhea, fever, abdominal cramps, and vomiting (Ruiz-Palacios, 2007; Korsak, 2014; Skarp et al., 2016; Zang et al., 2018).
Most of the Campylobacter infections in humans are associated with the consumption of C. jejuni (80–90%) and Campylobacter coli (5–10%) contaminated meat (Biasi et al., 2011). In European countries, pork meat remains a main source for transmission of Campylobacter spp. to humans (Ghafir et al., 2008). Contaminated pig carcasses have been found in slaughtering line, where pig carcasses undergo communal scalding early and the skin will remain on the carcass during the following slaughtering procedures (Epps et al., 2013). C. coli isolates are often recovered from pig feces (Stella et al., 2017).
Although Campylobacter spp. contamination in animal origin food has been reported in several studies, most researches were focused on poultry and poultry products. It is not clear that the source of Campylobacter spp. contamination is on pig carcass, although pig has also been considered potential reservoir of Campylobacter spp. (Alban et al., 2008). In the farm stage, Campylobacter spp. can colonize the gastrointestinal tract of pigs and passes out of the host in its feces. During slaughter and subsequent steps, Campylobacter spp. might spread through the slaughtering environment and slaughtering operations, and finally contaminate the food chain (Hansson et al., 2007; Hermans et al., 2012).
The risk of Campylobacter infection through processing of meat needs to be investigated carefully as large quantities of pork meat are consumed, especially in eastern China (Nesbakken et al., 2003). However, the epidemiological data on Campylobacter spp. infection in Asia is still limited, and incidences reported by other countries may vary substantially (Kaakoush et al., 2015).
To investigate the potential source of Campylobacter spp. contamination, a quantitative research on the prevalence of Campylobacter spp. in pigs were conducted in a slaughter house. Campylobacter spp. isolates were isolated from each batch of pigs in the slaughtering line, slaughtering environment, and the upstream farm. Multilocus sequencing typing (MLST) tool was used to interpret the mode of the transmission of Campylobacter spp.
Materials and Methods
Sample collection
A total of 2056 samples from 12 batches of 214 pigs (214 pigs = 11 batches × 18 pigs per batch +1 batch × 16 pigs in one batch) were collected from a representative slaughterhouse and its upstream farm in Jiangsu province between March 2016 and April 2017. The slaughterhouse is affiliated to a provincial large-scale food processing enterprise with a canonical production pattern and management system, which extends from the farm to the market. The upstream farm selling the pigs to the slaughtering house and the average daily slaughter capacity of the slaughterhouse were 500–1000 pigs.
In detail, the samples contain 54 cloacal swabs from 54 pigs on the farm, 1712 wipe samples from 214 pig carcasses under 8 main slaughtering processes, 14 cloacal swabs from 14 pigs at lairage, and 276 samples from the slaughtering environment. Each batch samples from the farm and the slaughter house were collected on the same day. At 5 AM, 54 pigs were randomly selected and labeled at the farm, 54 cloacal swab samples were collected and the front parts of these swabs were completely immersed into Cary-Blair transport medium for storage. Then, it takes about 3 h for pigs to be transported from the farm to the slaughterhouse.
When the pigs arrived at the slaughterhouse, besides of the former labeled 54 pigs in the farm, another 160 pigs were also randomly selected and labeled. To determine the prevalence of Campylobacter spp. in the slaughter house accurately, each labeled pig or carcass was tracked along the whole slaughtering line, samples were only collected from the labeled pigs or carcasses (Huang et al., 2017), based on the biological characteristics of Campylobacter spp. and the sample collection method suggested in the Hazard Analysis and Critical Control Point (HACCP) regulations.
At the lairage of slaughter house, 14 cloacal swab samples were collected after the pig entered the slaughterhouse lairage using sterile swabs. Then, 1712 samples were collected from pigs during the following eight main steps in the slaughtering line: entrance (n = 214), dehairing (n = 214), flaming (n = 214), evisceration (n = 214), washing and chilling (n = 214), polishing (n = 214), spray cooling (n = 214), and flash-freezing (n = 214).
At entrance, cloacal swab samples were collected before the slaughter. The prevalence and load of Campylobacter spp. isolates from these cloacal samples were considered as the initial contamination load. When the pig carcasses under the processes of dehairing and singeing modification, cotton balls (immersed with 2 mL sterile phosphate buffered saline [PBS], pH 7.2) were used to wipe the surfaces of the pig carcasses, including the following five sampling sites: neck, feet, abdomen, back, and hips, 100 cm2 of each part was wiped with one cotton ball, five cotton balls combined into one sample covering a total of 500 cm2 (Huang et al., 2017).
Later, the pig carcass was cut in half and the following slaughtering processes were undertaken: evisceration, washing and chilling, polishing, spray cooling, and flash-freezing. In these processes, both the inner and outer surfaces of pig neck, back, and bottom were wiped evenly (100 cm2 of each part was wiped with one cotton ball, six cotton balls combined into one sample covering a total of 600 cm2). To facilitate calculation, the unit of Campylobacter spp. load in the surface of wiping sample was converted to colony-forming units (CFU)/500 cm2, while the unit of Campylobacter spp. load in cloacal swab sample was CFU/g.
A total of 276 environmental samples were collected during slaughter processes using a five-point sampling method with cotton balls, one cotton ball for sampling was immersed with 2 mL sterile PBS (pH 7.2) in advance. The sampling sites included the surface of the floor, countertop, operating tools, worker's gloves, and soles. The environmental samples were sealed tightly in sterile plastic wraps and transported to laboratory for Campylobacter spp. isolation.
Isolation and quantification of Campylobacter spp.
Campylobacter spp. was quantified using the direct plating method (Scherer et al., 2006). Each cloacal swab sample was immersed in 1 mL sterile PBS for 20 min, after several times of vortexing, removing the swab to access the original rinse solution. For carcass wiping sample, the original rinse solution was accessed by compressing cotton balls. The rinse solutions were diluted 10-fold with PBS sequentially, then 0.1 mL diluted rinse solution at each dilution ratio was added onto Campylobacter blood-wet selective agar plates (modified CCDA-Preston; Oxoid, United Kingdom). These plates were cultured in microaerophilic conditions (10% CO2, 5% O2, 85% N2) and incubated at 42°C for 48 h. A repeated trial was performed at the same time.
The dilution that contained the confirmed colonies within the countable range (15–300 CFU per plate) was used for enumeration (Ministry of Health of the People's Republic of China, 2008). Five potential Campylobacter spp. colonies that showed moist and gray with similar colony morphologies in the plate were picked and cultured on Mueller Hinton Agar with Blood plates (Difco, BD, Sparks, MD) for another 48 h at 42°C. A single suspected colony was selected into a sterile fingerlike tube with 100 μL PBS added and then heated at 100°C for 10 min for lysis. The sample was then centrifuged at 16,000 × g for 3 min, and the supernatant containing bacterial DNA was immediately transferred to a new test tube and stored at −20°C until used.
Campylobacter were identified at the species level using multiplex polymerase chain reaction (PCR), which concurrently identified the 16S rRNA for all Campylobacter species, mapA gene for C. jejuni, and ceuE gene for C. coli (Nielsen et al., 1997). The PCR mixture included 2 μL of deionized water, 1 μL of each upstream and downstream primer (10 μmol/L), 2 μL of bacterial DNA, 10 μL of Taq Master Mix (Vazyme, Nanjing, China), and the final volume was 20 μL of water. The amplification reaction was carried out according to the following procedures: first, predenaturation at 95°C for 10min; then, the following 35 cycles were composed of three parts: denaturation at 95°C for 30 s, annealing at 59°C for 90 s, and extension at 72°C for 1 min. Finally, it was extended for 10 min at 72°C.
The number of Campylobacter spp. on a single plate (CFU) = the number of suspected Campylobacter spp. colonies × the number of positively identified Campylobacter spp. colonies/5 (Huang et al., 2015). The total amount of Campylobacter spp. in a single sample (CFU) = the mean number of Campylobacter spp. colonies on two duplicate plates × dilution ratio.
MLST analysis
To investigate Campylobacter spp. propagation in pig slaughtering line, 62 Campylobacter spp. isolates were obtained from one slaughtering batch (18 pigs were tracked for sample collection) in Jiangsu slaughterhouse, and its upstream farm was also selected for MLST analysis. The genomic DNA of Campylobacter isolates was extracted using TIANamp Bacterial DNA Kit (Tiangen Biotech, Beijing, China) according to the manufacturer's instruction and stored at −20°C. MLST was performed following the guidelines of the Campylobacter spp. MLST website (
Amplification primers and sequencing primers were used as prescribed in protocol. The PCR mixture included 1 μL 10 mM dNTP (TaKaRa, Dalian, China), 0.01 μmol/mL forward primer, 0.01 μmol/mL reverse primer, 0.5 U of TaqDNA polymerase (Vazyme), 2.5 μL 10 × PCR buffer (Vazyme), 1 μL template DNA, and RNase-free water (TaKaRa) was added to obtain a final volume of 25 μL. The PCR conditions were followed as initial denaturation at 94°C for 5 min, followed by 30 cycles of denaturation at 94°C for 30 s, annealing at 50°C for 1 min, and extension at 72°C for 5 min. Amplicons were detected on 1.5% ethidium bromide agarose gel and purified using QIAamp Gel Extraction Kit (Qiagen, Hilden, Germany) according to the manufacturer's instruction.
Sequences were analyzed by Molecular Evolutionary Genetics Analysis (MAGA). Alleles, sequence types (STs), and clonal complexes were assigned by submitting DNA sequences of amplicons to the MLST database (
Statistical analysis
SPSS (version 19.0; SPSS, Chicago, IL) was used for data analysis. Ninety-five percent confidence interval of Campylobacter spp. prevalence and load were determined. The p-value ≤0.05 was considered statistically significant.
Results
Prevalence of Campylobacter spp. positive in pig slaughtering line
A total of 333 Campylobacter spp. isolates (19.30%) out of 1726 different samples were isolated from pig slaughtering line (Table 1). The isolation rate of Campylobacter spp. in each link was ranged between 2.8% and 42.5%. The prevalence of Campylobacter spp. in lairage represented the original contamination (35.7%, 5/14), and the process of flash-freezing was considered the last Campylobacter spp. contaminated slaughtering link. Chi-square test was used for statistical analysis, data of all links will be compared with that of the previous link, and significant tests were conducted. Campylobacter spp. isolation rate was highest during entrance (42.5%, 91/214).
The Prevalence of Campylobacter spp. in Pig Slaughtering Line
The quantitative unit of Campylobacter was log CFU/g. In the processes of Lairage and Entrance, cloacal swab sample were collected, but in the other slaughtering processes, surface wiping samples were collected.
CFU, colony-forming units.
Compared to the isolation rate of Campylobacter spp. after dehairing (28%, 60/214), the contamination of Campylobacter spp. in flaming was significantly reduced to 2.8% (6/214, p < 0.05). Followed by flaming, the isolation rate of Campylobacter spp. after evisceration was significantly increased to 29.4% (63/214, p < 0.05). After the processes of washing and chilling, the isolation rate of Campylobacter spp. was significantly reduced to 18.2% (39/214, p < 0.05). After the next processes of the polishing (15.0%, 32/214) and spray cooling (13.6%, 29/21), the contamination of Campylobacter spp. in pig carcass was reduced, both the effects were not significant (p > 0.05). However, after the last slaughtering process flash-freezing, Campylobacter spp. isolation rate in pig carcass was significantly reduced to 3.7% (8/214, p < 0.05).
Quantitative analysis of Campylobacter spp. under different pig slaughter processes is shown in Table 1. The load of Campylobacter spp. in pig carcass samples collected along the slaughtering line ranged from 1.47 ± 0.38 log CFU/500 cm2 to 4.87 ± 1.23 log CFU/500 cm2.
In the process of entrance, the load of Campylobacter spp. in pig cloacal swab samples was found to be the highest (4.87 ± 1.23 log CFU/g), then the load of Campylobacter spp. in pig carcass samples collected after dehairing was significantly reduced to 2.40 ± 0.49 log CFU/500 cm2 (p < 0.05). Campylobacter spp. load in samples collected after evisceration were increased to 2.50 ± 0.54 log CFU/500 cm2, although this change was not significant (p > 0.05). After the next processes of washing and chilling, polishing, and spray cooling, Campylobacter spp. contamination load in pig carcass was slowly decreased to 1.88 ± 0.14 log CFU/500 cm2 (p > 0.05). After the last slaughtering process flash-freezing, the load of Campylobacter spp. in pig carcass samples was reduced to 1.89 ± 0.15 log CFU/500 cm2 (p > 0.05).
Prevalence of Campylobacter spp. in slaughtering environments and farm
The isolation rate of Campylobacter spp. in slaughtering environmental samples was 27.9% (77/276), and 64 Campylobacter spp. were isolated from the environment of evisceration. Among the 64 Campylobacter spp. isolates, 39 were isolated from the knife's surface (60.9%).
The Campylobacter spp. isolation rate in the samples collected from ground, water, and countertop was 10.9%, 9.4%, and 7.8%, respectively. Furthermore, the isolation rate of Campylobacter spp. in samples collected after washing and chilling, polishing, and spray cooling was 16.7%, 6.3%, and 8.3%, respectively. Campylobacter spp. strains were not found in the environment of flaming and flash-freezing (Table 2). Moreover, the isolation rate of Campylobacter spp. in pig cloacal swabs on the farm was 18.5% (10/54), this sampled farm was assumed as another possible contamination source, which has sold live pigs to the slaughtering house.
The Prevalence of Campylobacter spp. in the Environment of Pig Slaughterhouse
CFU, colony-forming units.
Quantitative analysis of the slaughtering environmental samples showed that the Campylobacter spp. load was as follows: evisceration (3.13 ± 1.89 log CFU/500 cm2) > spray cooling (2.56 ± 0.92 log CFU/500 cm2) > dehairing (2.36 ± 0.78 log CFU/500 cm2). In addition, the load of Campylobacter spp. in pig cloacal swabs on the farm was 2.25 ± 0.78 log CFU/500 cm2.
MLST analysis of Campylobacter spp. isolates
Sixty-two Campylobacter spp. isolates were obtained from one batch of the samples and identified as C. coli by multiplex PCR. Through cluster analysis, these isolates were grouped into seven clusters (A–G) (Fig. 1). Among the 62 Campylobacter isolates, there were 51 C. coli grouped in cluster-C, and all these C. coli strains belonged to ST-828 complexes. The sources of these C. coli strains covered the whole slaughtering line, slaughtering environment, and the upstream farm.
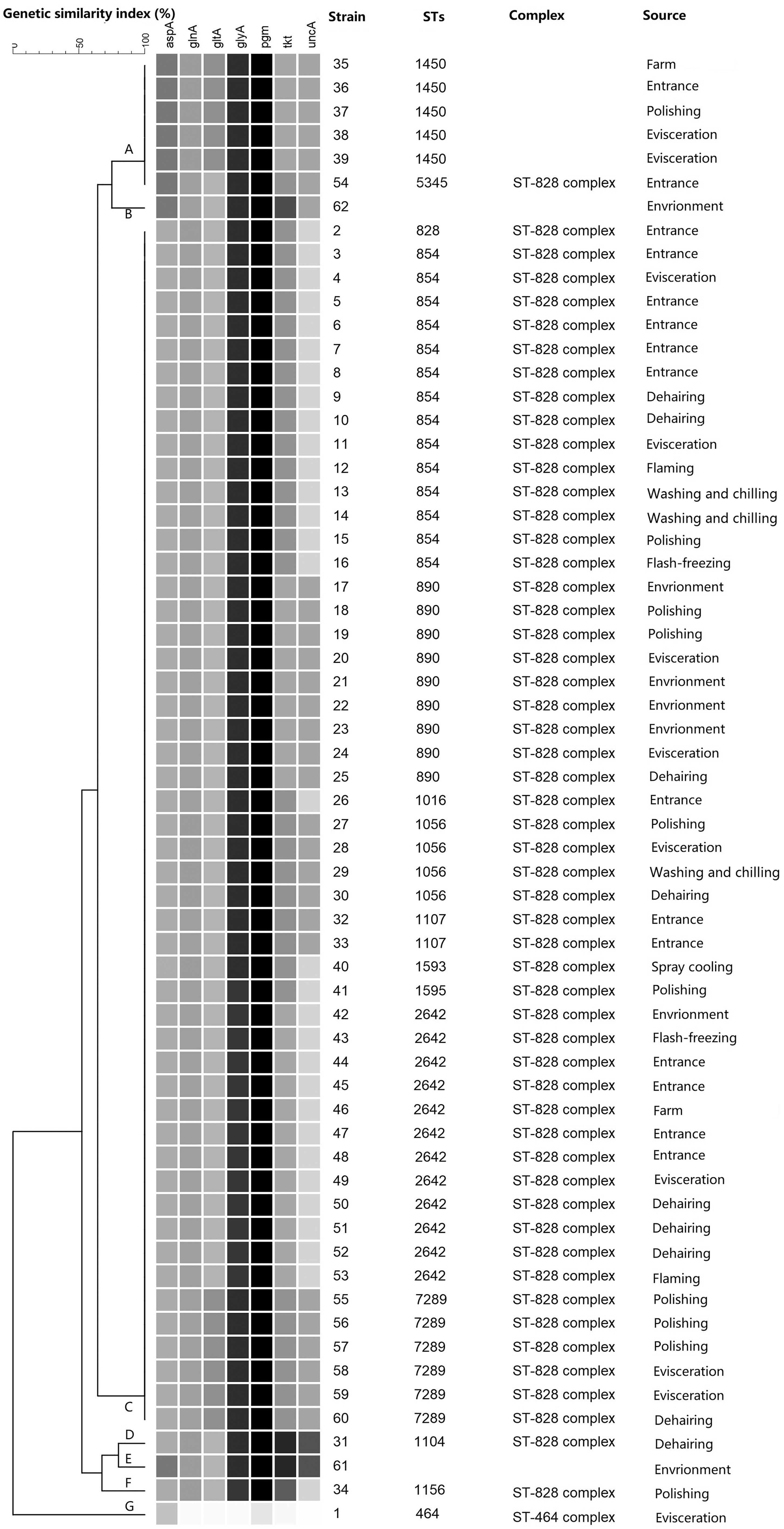
An UPGMA dendrogram of Campylobacter coli isolates from pig farm and slaughterhouse. Sixty-two C. coli isolates were grouped into seven clusters (A–G) with high homology. STs, sequence types; UPGMA, unweighted pair group method with arithmetic means.
Discussion
Human campylobacteriosis remains the most commonly reported gastrointestinal disease worldwide (Rodgers et al., 2017; Zang et al., 2017). The main transmission mode of Campylobacter spp. is through the consumption of undercooked meat (EFSA, 2017; Narvaezbravo et al., 2017). Pig slaughtering line, where the occurrence of Campylobacter spp. is common, has always been considered as a key link in pork meat safety control. However, due to the sampling difficulties at slaughterhouse and difficulty with growing and enumerating Campylobacter spp., few studies have been able to trace microbial contamination to an individual pig carcass (Biasi et al., 2011; Maramski, 2012).
In this study, Campylobacter spp. may be transmitted along the pig slaughtering line with an average prevalence of 19.3%. In previous studies, the Campylobacter spp. isolation rate in pig carcasses at slaughterhouse was reported to range from 13.0% to 81.3%, differences could probably result from the sampling method, regional differences, and slaughtering processes (Mattheus et al., 2012; Morales-Partera et al., 2018). We found that the isolation rate of Campylobacter spp. in cloacal swab samples collected at the entrance was 42.5%, which was consistent with a previous study reporting the isolation rate of Campylobacter spp. in pig intestinal content samples collected in China slaughterhouses from 2008 to 2014 was 48.9% (Wang et al., 2016).
In our study, the isolation rates of Campylobacter spp. in samples collected after dehairing (28%) and evisceration (29.4%) were qualitatively higher than other slaughtering processes. The leakage of intestinal contents or nonstandard operations during evisceration could lead to the contamination of Campylobacter spp. The processes of dressing and chilling can reduce the contamination of Campylobacter spp. in pig carcasses (Maramski, 2012). The isolation rate of Campylobacter spp. decreased significantly after washing and chilling (18.2%, p < 0.05) and flash-freezing (3.7%, p < 0.05), which demonstrated that a low temperature could effectively inhibit the contamination of Campylobacter spp.
However, it is critical to provide the quantitative data rather than the prevalence of Campylobacter spp. to identify the risk for consumers, as it is known that a reduction of counts on livestock carcass surfaces could result in a significant reduction in human campylobacteriosis cases (Nauta et al., 2009). In this study, the contamination load of Campylobacter spp. in samples collected after dehairing was 2.40 ± 0.49 log CFU/500 cm2, then it decreased to 1.89 ± 0.15 log CFU/500 after the process of flash-freezing (p > 0.05).
The most common species of Campylobacter spp. associated with human infections are C. jejuni and C. coli, C. coli is the more common Campylobacter species recovered from swine (Horrocks et al., 2009). In some studies, for instance, C. coli has been recovered from swine fecal samples at greater than 99% (Alter et al., 2005; Thakur et al., 2005).
In our study, C. coli was the predominant species from pig carcass. However, C. jejuni was isolated from one wipe sample after dehairing. We speculated that this C. jejuni isolate was present in the hair of pig, as hair can easily adhere to the tools, ground, and conveyor belt in depilation area. It is also possible that C. jejuni was present in the remnants of the previous slaughtering batch or isolated due to cross contamination caused by the wrong operation of slaughterhouse worker, this requires further investigation. Moreover, 32 other Campylobacter spp. (non-C. jejuni and non-C. coli) were recovered during evisceration, it indicated that the possibility of finding vital forms of Campylobacter spp. in meat samples is greater than that in pig cloacal swabs (Maramski, 2012).
Analyzing the prevalence of Campylobacter spp. in slaughtering environment samples and the pig cloacal swabs from farm helped to better understand the contamination source of Campylobacter spp. in pig slaughtering line. The isolation rate of Campylobacter spp. in the pig cloacal swabs on farm was 18.5% (10/54), the farm which supplied pigs to slaughterhouse could be considered as a potential contamination source. The contamination load of Campylobacter spp. in the isolates from the farm was 2.25 ± 0.78 log CFU/g, while the load of Campylobacter isolates from the lairage of the slaughter house was 1.47 ± 0.38 log CFU/g. The carrier level of Campylobacter spp. on farm sample was almost flat with the data of lairage. Besides host animal, the environmental niche has also been considered an important contamination source of Campylobacter spp. (Whiley et al., 2013).
The isolation rate (82.1%) and load (3.13 ± 1.89 log CFU/500 cm2) of Campylobacter spp. in environmental samples collected during evisceration was the highest, and there could be a high possibility of cross-contamination during slaughtering operational processes.
Campylobacter spp. spread in the environment through feces, and they can also be found in soil and water (Hermans et al., 2012). In our study, Campylobacter spp. were isolated from the cutting tool samples (60.9%, n = 39), ground samples (10.9%), water samples (9.4%) and countertop samples (7.8%). Although the slaughtering knives are used based on the HACCP-based hygienic guidelines, there is always a risk of incomplete disinfection and operational errors. The surface-wiping sample of knife has been classified as an important part of the environment samples. Botteldoorn et al. reported that the knife samples in four slaughterhouses were found positive for Salmonella, when the knife blades were sampled by swabbing its tip to its base twice (Botteldoorn et al., 2003).
It is well known that a pig carcass can easily be contaminated with intestinal waste during slaughtering, and the knives are likely to be contaminated with Campylobacter spp. from the intestinal waste, especially in the process of evisceration. In this study, samples collected from the surface of the knife were positive for Campylobacter spp. We have informed the slaughtering enterprise that the knife is to be considered a risk factor during pork meat quality control. Better hygiene management on workers' standard operating procedure and slaughtering equipment could reduce the pathogen levels in pig slaughterhouse.
In addition, no Campylobacter spp. was isolated from the flaming, indicating that the Campylobacter spp. on the surface of pig carcass could be killed after the process of flaming. No Campylobacter spp. was detected in the environment of cold storage and flash-freezing as low temperature may have inhibited its growth. The occurrence of Campylobacter spp. in slaughtering environments and farm indicated that the cross contamination of Campylobacter was possible.
Epidemiological investigation combined with molecular typing could determine the transmission route of Campylobacter spp. By cluster analysis, 62 C. coli strains isolated from one batch in the Jiangsu province were grouped into seven clusters (A–G), cluster-C covered the whole slaughtering line, slaughtering environment, and farm, and all the strains in cluster-C were grouped to the 828-complex. Results of MLST indicated that C. coli could be propagated in the entire slaughterhouse (especially during dehairing and evisceration).
Our results highlighted that the upstream farm was another vital but often neglected contamination source for the pig carcasses in slaughterhouse. Horizontal transmission of Campylobacter spp. in the farm represents the usual route of transmission. Once Campylobacter spp. is introduced into a pig flock, it spreads rapidly and colonizes in the intestinal tracts of the majority of pigs until slaughter (Beatriz, et al., 2018). Campylobacter spp. would be widely present in the farm surrounding environment once a flock becomes positive, persist for several weeks, and in some occasions even before pig placement. The main purpose of this article is to explore the Campylobacter spp. prevalence in pig slaughter line, so we just collected one batch of farm samples.
The rules of breeding stage (farm) will be researched in the future. Besides the farm, the environmental factors in the slaughterhouse (contact surfaces and handling by workers) have also been considered as the major Campylobacter spp. contamination sources for pig carcasses (Borch et al., 1996). Therefore, appropriate practices and hygiene measures should also be implemented in the vital pig slaughtering processes and in the farms so that the risk of Campylobacter contamination could be reduced.
Pig slaughter is an open process with many opportunities for the contamination of the carcass with Campylobacter spp., it does not contain any step that could be considered completely free of hazard (Biasi et al., 2011). Slaughtering plant should control the contamination of Campylobacter spp. from the source in construction project, according to cleaner production principle.
Conclusions
In this study, we investigated the possible contamination source of Campylobacter spp. in the pig slaughtering line. The prevalence of Campylobacter spp. during entrance, dehairing, and evisceration were qualitatively and quantitatively higher than other processes. In addition, farm and slaughtering environment were vital but often underestimated Campylobacter spp. contamination sources in pig slaughtering chain. Special attentions are required during dehairing and evisceration, and the upstream farm needs to be monitored continuously for potential contamination. Our study provides essential data to support the strategies of Campylobacter spp. alleviation in pork meat production.
Footnotes
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Key Research and Development Program of China (2018YFD0500500), National Natural Science Foundation of China (31872493), the Priority Academic Program Development of Jiangsu Higher Education Institutions, Leading Talents Program of Yangzhou University, and the six talent peaks project in Jiangsu Province (2015-SWYY-02).
Appendix
Multilocus Sequencing Typing Date of Campylobacter spp. in Pig Slaughtering Line for the Construction of Dendrogram
| Isolate | aspA | glnA | gltA | glyA | pgm | tkt | unc |
ST | Clonal complex | Source |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 33 | 38 | 30 | 79 | 104 | 35 | 17 | 2642 | ST-828 complex | Environment |
| 2 | 53 | 38 | 30 | 82 | 104 | 43 | 36 | 5345 | ST-828 complex | Entrance |
| 3 | 33 | 39 | 30 | 82 | 104 | 43 | 36 | 1056 | ST-828 complex | Polishing |
| 4 | 33 | 39 | 30 | 82 | 104 | 43 | 17 | 828 | ST-828 complex | Entrance |
| 5 | 33 | 38 | 30 | 79 | 104 | 35 | 17 | 2642 | ST-828 complex | Flash-freezing |
| 6 | 24 | 2 | 2 | 2 | 10 | 3 | 1 | 464 | ST-464 complex | Evisceration |
| 7 | 33 | 38 | 30 | 82 | 104 | 35 | 36 | 890 | ST-828 complex | Environment |
| 8 | 33 | 39 | 30 | 82 | 104 | 43 | 36 | 1056 | ST-828 complex | Evisceration |
| 9 | 33 | 38 | 30 | 82 | 104 | 35 | 36 | 890 | ST-828 complex | Polishing |
| 10 | 33 | 38 | 30 | 82 | 104 | 35 | 36 | 890 | ST-828 complex | Polishing |
| 11 | 33 | 38 | 44 | 82 | 104 | 43 | 36 | 7289 | ST-828 complex | Polishing |
| 12 | 33 | 38 | 44 | 82 | 104 | 43 | 36 | 7289 | ST-828 complex | Polishing |
| 13 | 33 | 38 | 44 | 82 | 104 | 43 | 36 | 7289 | ST-828 complex | Polishing |
| 14 | 33 | 38 | 30 | 79 | 104 | 43 | 17 | 1595 | ST-828 complex | Polishing |
| 15 | 33 | 38 | 30 | 82 | 104 | 43 | 17 | 854 | ST-828 complex | Entrance |
| 16 | 33 | 38 | 30 | 79 | 104 | 35 | 17 | 2642 | ST-828 complex | Entrance |
| 17 | 33 | 38 | 30 | 79 | 104 | 35 | 17 | 2642 | ST-828 complex | Entrance |
| 18 | 33 | 38 | 44 | 82 | 104 | 43 | 36 | 7289 | ST-828 complex | Evisceration |
| 19 | 33 | 38 | 44 | 82 | 104 | 43 | 36 | 7289 | ST-828 complex | Evisceration |
| 20 | 33 | 38 | 44 | 82 | 104 | 43 | 36 | 7289 | ST-828 complex | Dehairing |
| 21 | 33 | 38 | 30 | 82 | 104 | 35 | 36 | 890 | ST-828 complex | Evisceration |
| 22 | 33 | 38 | 30 | 82 | 104 | 43 | 17 | 854 | ST-828 complex | Evisceration |
| 23 | 32 | 39 | 30 | 82 | 104 | 43 | 17 | 1593 | ST-828 complex | Spray cooling |
| 24 | 33 | 38 | 30 | 82 | 104 | 35 | 36 | 890 | ST-828 complex | Environment |
| 25 | 33 | 38 | 30 | 82 | 104 | 35 | 36 | 890 | ST-828 complex | Environment |
| 26 | 33 | 38 | 30 | 82 | 104 | 35 | 36 | 890 | ST-828 complex | Environment |
| 27 | 33 | 39 | 30 | 79 | 104 | 64 | 17 | 1156 | ST-828 complex | Polishing |
| 28 | 33 | 38 | 30 | 82 | 104 | 35 | 36 | 890 | ST-828 complex | Evisceration |
| 29 | 33 | 38 | 30 | 82 | 118 | 43 | 17 | 1016 | ST-828 complex | Entrance |
| 30 | 33 | 38 | 30 | 82 | 104 | 43 | 17 | 854 | ST-828 complex | Entrance |
| 31 | 33 | 38 | 30 | 82 | 104 | 43 | 17 | 854 | ST-828 complex | Entrance |
| 32 | 33 | 38 | 30 | 82 | 104 | 43 | 17 | 854 | ST-828 complex | Entrance |
| 33 | 33 | 38 | 30 | 79 | 104 | 35 | 17 | 2642 | ST-828 complex | Farm |
| 34 | 33 | 38 | 30 | 79 | 104 | 35 | 17 | 2642 | ST-828 complex | Entrance |
| 35 | 33 | 38 | 30 | 79 | 104 | 35 | 17 | 2642 | ST-828 complex | Entrance |
| 36 | 53 | 39 | 44 | 82 | 104 | 35 | 36 | 1450 | – | Farm |
| 37 | 33 | 38 | 30 | 79 | 104 | 35 | 17 | 2642 | ST-828 complex | Evisceration |
| 38 | 53 | 39 | 44 | 82 | 104 | 35 | 36 | 1450 | – | Entrance |
| 39 | 33 | 38 | 30 | 82 | 104 | 43 | 17 | 854 | ST-828 complex | Entrance |
| 40 | 33 | 38 | 30 | 82 | 104 | 44 | 36 | 1107 | ST-828 complex | Entrance |
| 41 | 33 | 39 | 30 | 82 | 104 | 43 | 36 | 1056 | ST-828 complex | Washing and chilling |
| 42 | 33 | 38 | 30 | 82 | 104 | 43 | 17 | 854 | ST-828 complex | Dehairing |
| 43 | 33 | 38 | 30 | 82 | 104 | 43 | 17 | 854 | ST-828 complex | Dehairing |
| 44 | 33 | 38 | 30 | 82 | 104 | 43 | 17 | 854 | ST-828 complex | Evisceration |
| 45 | 53 | 39 | 44 | 82 | 104 | 35 | 36 | 1450 | – | Polishing |
| 46 | 33 | 38 | 30 | 82 | 104 | 44 | 36 | 1107 | ST-828 complex | Entrance |
| 47 | 33 | 39 | 30 | 82 | 104 | 43 | 36 | 1056 | ST-828 complex | Dehairing |
| 48 | 53 | 39 | 30 | 82 | 104 | 85 | 68 | – | – | Environment |
| 49 | 33 | 38 | 30 | 82 | 104 | 43 | 17 | 854 | ST-828 complex | Flaming |
| 50 | 33 | 38 | 30 | 82 | 104 | 43 | 17 | 854 | ST-828 complex | Washing and chilling |
| 51 | 33 | 39 | 30 | 82 | 104 | 85 | 68 | 1104 | ST-828 complex | Dehairing |
| 52 | 53 | 38 | 30 | 82 | 104 | 70 | 36 | – | – | Environment |
| 53 | 53 | 39 | 44 | 82 | 104 | 35 | 36 | 1450 | – | Evisceration |
| 54 | 33 | 38 | 30 | 79 | 104 | 35 | 17 | 2642 | ST-828 complex | Dehairing |
| 55 | 33 | 38 | 30 | 82 | 104 | 43 | 17 | 854 | ST-828 complex | Washing and chilling |
| 56 | 33 | 38 | 30 | 82 | 104 | 35 | 36 | 890 | ST-828 complex | Dehairing |
| 57 | 33 | 38 | 30 | 82 | 104 | 43 | 17 | 854 | ST-828 complex | Polishing |
| 58 | 33 | 38 | 30 | 79 | 104 | 35 | 17 | 2642 | ST-828 complex | Dehairing |
| 59 | 33 | 38 | 30 | 79 | 104 | 35 | 17 | 2642 | ST-828 complex | Dehairing |
| 60 | 33 | 38 | 30 | 79 | 104 | 35 | 17 | 2642 | ST-828 complex | Flaming |
| 61 | 53 | 39 | 44 | 82 | 104 | 35 | 36 | 1450 | – | Evisceration |
| 62 | 33 | 38 | 30 | 82 | 104 | 43 | 17 | 854 | ST-828 complex | Flash-freezing |
–, new complex or new ST.
ST, sequence type.