
Abstract
Trans-cinnamaldehyde (TC), a typical plant-derived compound, has been widely used in the control of foodborne pathogen contamination. Nevertheless, the risk associated with the occurrence of viable but nonculturable (VBNC) bacteria induced by TC remains unclear. The results of this study showed that Salmonella Enteritidis (S. Enteritidis) entered the VBNC state after being induced by TC at a minimum inhibitory concentration of 312.5 μg/mL and survived for at least 22 days under TC treatment. Enhanced resistance was found against heat treatment (75°C, 30 s), antibiotics (i.e., ampicillin, ceftriaxone sodium, chloramphenicol), and hydrogen peroxide (3%) in VBNC S. Enteritidis. A synergistic effect against VBNC S. Enteritidis occurred when TC was combined with acid treatment, including lactic acid and acetic acid (pH = 3.5). VBNC and resuscitated S. Enteritidis by sodium pyruvate treatment (100 mM) were found to retain the infectious ability to Caco-2 cells. Relative expression levels of the stress-related genes relA, spoT, ppx, lon, katG, sodA, dnaK, and grpE were upregulated in VBNC S. Enteritidis. Accumulation of reactive oxygen species (ROS) and protein aggregates was observed in VBNC cells. Besides, the resuscitation of VBNC cells was accompanied with clearance of ROS and protein aggregates. In summary, this study presents a comprehensive characterization of stress tolerance and resuscitation of VBNC S. Enteritidis induced by cinnamaldehyde, and the results provide useful information for the development of effective control strategy against VBNC pathogenic bacteria in food production.
Introduction
Salmonella enterica is one of the most important foodborne pathogens responsible for global public health. It was estimated to cause 93.8 million cases of gastroenteritis with 155,000 deaths worldwide annually (Majowicz et al., 2010). Foods such as poultry, fresh fruits, and vegetables have been reported as vehicles for salmonellosis (Ehuwa et al., 2021). Food production provides stressors (e.g., antimicrobials, acidity, and osmotic pressure) for foodborne pathogens, which may promote bacteria to enter a viable, but nonculturable (VBNC) state (Li et al., 2020). Despite losing culturability, VBNC pathogenic bacteria maintain respiratory activity and infectious ability (Highmore et al., 2018), and is hard to detect under stressed environment (Mao et al., 2021); thus, the presence of VBNC Salmonella poses serious threats to food safety.
Biological characteristics of pathogenic bacteria may alter under VBNC state, which is generally associated with changes in metabolic pathways (Li et al., 2021). VBNC Salmonella was found to shift from rod shaped to spherical (Fu et al., 2020). Increased heat tolerance, antimicrobial resistance, as well as altered virulence and gene expression have been reported in VBNC Salmonella in various foods (Chen et al., 2021, Jayeola et al., 2022, Li et al., 2020).
Accumulation of reactive oxygen species (ROS), protein aggregation, and the involvement of (p)ppGpp synthesis were proposed to be critical mechanisms for the formation of VBNC bacteria (Zhang et al., 2023). The oxidative stress regulator OxyR has been suggested to be associated with the induction of VBNC state (Li et al., 2014). Protein aggregation is a key factor that can drive the development and persistence of VBNC state (Bollen et al., 2021). Besides, protein disaggregation could lead to the release of amino acids for osmoprotection (Wang et al., 2023b). The alarmone ppGpp, whose production is regulated by relA and spoT, could trigger the stringent response of bacteria in harsh conditions and result in the advent of the VBNC state (Bai et al., 2021).
Trans-cinnamaldehyde (TC) is a typical kind of plant essential oil, which can be obtained from the inner bark of trees within the genus Cinnamomum (Shreaz et al., 2016). This natural bacteriostatic substance is an approved food additive and has been widely applied in foods for its antimicrobial activity (Friedman, 2017). Treatment of TC might induce foodborne pathogens such as Pseudomonas aeruginosa and Staphylococcus aureus into VBNC state (Bouhdid et al., 2010). Besides, it was reported that cinnamon oils could induce cross-resistances in S. enterica (Chen et al., 2022). Nevertheless, the effect of TC treatment in the induction of VBNC Salmonella is still unclear.
This study investigated changes of morphology, physiology, resistance, and gene expression in VBNC Salmonella enterica serotype Enteritidis (S. Enteritidis) induced by TC, which can provide a scientific basis for the control of VBNC Salmonella in food production and facilitate a better understanding for the mechanism of VBNC.
Materials and Methods
Strains and culture
The bacterial strain S. Enteritidis CICC 21482 was purchased from China Center of Industrial Culture Collection (Beijing, China) and stocked at −80°C. The stock culture was activated in tryptic soy broth with yeast extract (TSB-YE, Guangdong Huankai, China) at 37°C overnight, followed by the transfer of a single colony to 10 mL TSB-YE medium for cultivation for 16 h at 37°C. Log phase S. Enteritidis was then harvested by centrifugation at 8,000 × g, 4°C for 10 min using a high-speed centrifuge (Hunan Xiangyi, China), and resuspended with sterile physiological saline (PS, w/w, 0.85%), with a final concentration of ∼8 log10 CFU/mL.
Induction of VBNC S. Enteritidis
Trans-cinnamaldehyde (TC, Shanghai Aladdin, China) and Tween 80 were mixed at the ratio of 10% (w/w) and 6% (w/w), respectively, with supplement of TSB up to 10 mL. The emulsion of TC was employed for the determination of minimum inhibitory concentration (MIC) against S. Enteritidis (∼8 log10 CFU/mL), referring to the study (Al-Bakri et al., 2009). The assay was performed using 96-well microplates with TC serially diluting to 2500, 1250, 625, 312.5, 156.25, and 78.125 μg/mL. After a 24-h cultivation at 180 rpm, at 37°C, the MIC value was determined as the lowest concentration, which prevented visible growth (Palaniappan and Holley, 2010), under which S. Enteritidis was considered to be in VBNC state. The turbidity was compared with a control without TC. Quantitative analysis of the presence of culturable and VBNC cells was performed as described in the section below.
Quantification of VBNC S. Enteritidis
After TC treatment, S. Enteritidis was plated onto TSB-YE plates and incubated at 37°C for enumeration. The number of resultant colonies was counted as colony-forming unit (CFU), which was used as an indicator of culturability.
Viable population of S. Enteritidis was quantified by qPCR with propidium monoazide dye treatment (PMAxx-qPCR) (Emerson et al., 2017). PMAxx-qPCR was conducted according to the procedure described previously (Ma et al., 2024). The qPCR reaction was performed on genomic DNA using primers (invA-F and invA-R) and probe (invA-Tx-208). VBNC cells were measured by the difference between the number of viable and culturable S. Enteritidis. Quantification of VBNC S. Enteritidis was conducted under MIC treatment every three days for a 22-day survival assay.
Resuscitation of VBNC S. Enteritidis
Catalase (CAT, 1000U/mL), sodium pyruvate (SP, 100 mM) and cell-free supernatant (CFS, 1:100) of log phase S. Enteritidis were used for resuscitation assays. These reagents were diluted with sterile PS and sterilized using 0.22 μM filter. Pellets of VBNC cells were resuspended with SP, CAT, and CFS dilution, respectively, followed by incubation at 37°C for 72 h. Each 100 μL aliquots of bacteria was spread on TSB-YE plates for CFU determination every 24 h. TSB medium supplemented with PS was used as the negative control for CFS group.
Intracellular ROS levels of S. Enteritidis
ROS levels of VBNC and resuscitated S. Enteritidis were measured. Approximately 7 log10 CFU/mL of bacteria was harvested by centrifugation at 10,000 × g for 5 min, and then incubated with H2DCFDA probe (10 μM, Beijing Labgic Technology, China) in dark for 30 min. They were then resuspended with PS before being pipetted to 96-well plates (100 μL/well) for measurement, with excitation and emission at 485 nm and 538 nm by microplate reader (TriStar2S, Berthold Technologies, Germany) (Kalyanaraman et al., 2012).
Resistances of VBNC S. Enteritidis to heat, acid, antibiotics, and H2O2
The culturable and VBNC S. Enteritidis were resuspended with PS and heated at 60°C for 10 min or 75°C for 30 s in heating block. They were then immediately immersed in ice water for cooling, followed by PMAxx-qPCR for viable quantification. As for acid treatment, the pellets of culturable and VBNC S. Enteritidis were resuspended with acetic acid and lactic acid, respectively (pH = 3.5 and 4.5), and thereby incubated for 24 h at 37°C in a shaker. In antibiotics challenge, culturable and VBNC S. Enteritidis were treated with 20 × MICs of levofloxacin (LCF), ceftriaxone sodium (CEF), ampicillin (AMP), and chloramphenicol (CHL) for 24 h at 37°C in a shaker at 180 rpm. MICs of antibiotics were determined by twofold dilution method. Under treatment of H2O2, the pellets were resuspended with 3% H2O2 in dark for 24 h, followed by PMAxx-qPCR for quantification.
Static gastrointestinal simulation assay
The gastrointestinal simulation assay of the culturable, VBNC, and resuscitated S. Enteritidis was conducted using a standardized in vitro digestion model (Brodkorb et al., 2019), with detailed procedure described previously (Li et al., 2023). In the gastric phase, approximately 7 log10CFU of bacterial pellets was added to a gastric simulation system, which was adjusted to pH 2.0 and then incubated at 37°C for 40 min. For the intestinal phase, the digestive juice was transferred to an intestinal simulation system, adjusted to pH 7.0, and then incubated at 37°C for 120 min. Viable quantification by PMAxx-qPCR was performed before gastric phase, after gastric phase, and after intestinal phase, respectively.
Bacterial adhesion and invasion assay
Adhesion and invasion capacities of culturable, VBNC, and resuscitated S. Enteritidis were measured using gentamicin protection assay (Mechesso et al., 2021), according to the procedure described previously (Li et al., 2023). Human colon adenocarcinoma cells (Caco-2; Shanghai Qida Biotechnology, China) were used for assays. Gentamicin protection assay was performed on a 24-well plate affixed with monolayer Caco-2 cells and approximately 7 log10CFU of bacteria to determine the adhesion and invasion ability of S. Enteritidis. The adhered and invasive bacterial populations were determined by using PMAxx-qPCR.
RNA extraction and reverse transcription quantitative PCR
RNA extraction and reverse transcription quantitative PCR (RT-qPCR) were performed according to the procedure described previously (Li et al., 2023). Quantification of gene expression was performed using a real-time PCR system (CFX Connect™, Bio-rad, USA). Each qPCR reaction contained 10 μL 2 × TB green (TaKaRa Biotechnology, Japan), 0.8 μL of each primer F/R, 2 μL cDNA, and 6.4 μL ddH2O. The primers for the selected genes and the procedures for RT-qPCR are shown in Table 1. Relative difference was calculated as fold change in formula 2−ΔΔCT. Expression changes > twofold with p < 0.05 were considered statistically significant.
Primers and Procedures Used for Gene Expression
The final concentration of primers of relA, lon, spoT, katG, sodA, and ppx is 0.1 μM in qPCR reaction system, and the final concentration of primers of dnaK and grpE is 0.4 Μm.
Transmission electron microscopy (TEM) analysis
VBNC and culturable S. Enteritidis were harvested by centrifugation at 10,000 × g and fixed with 2.5% glutaraldehyde at 4°C for 12 h, and then postfixed with 1% OsO4 for 2 h. Specimen preparation and TEM observation were performed according to the procedure described previously (Ma et al., 2024) using a Hitachi Model H-7650 TEM.
Protein aggregation measurement
The extraction of insoluble protein was performed according to the procedure detailed previously (Zuily et al., 2022). Bacterial pellets were harvested from ∼8 log10CFU/mL bacterial suspension. Cells were lysed and the aggregated proteins were isolated and loaded on 12% SDS-PAGE. The gels were treated by Coomassie blue protein stain. Variation of protein aggregates among culturable, VBNC, and resuscitated cells was compared with integrated density (total gray value) by Image J analysis.
Statistical analysis
The results were expressed as mean ± standard deviation. Statistical significance was determined by paired t test and one-way ANOVA using SPSS version 21.0.
Results
Survival of VBNC S. Enteritidis
In this study, MIC of TC against S. Enteritidis was 312.5 μg/mL, a 24-h treatment of which rendered S. Enteritidis into VBNC state. A viable population of about 6.71 ± 0.03 log10 CFU/mL was detected by using PMAxx-qPCR. Long-term survival of VBNC S. Enteritidis under MIC of TC was observed (Fig. 1). The population of VBNC cells decreased by 3.26 ± 0.09 log10 CFU/mL in the first 12 days, and were then maintained at ∼3 log10CFU/mL for the next 10 days (Fig. 1).
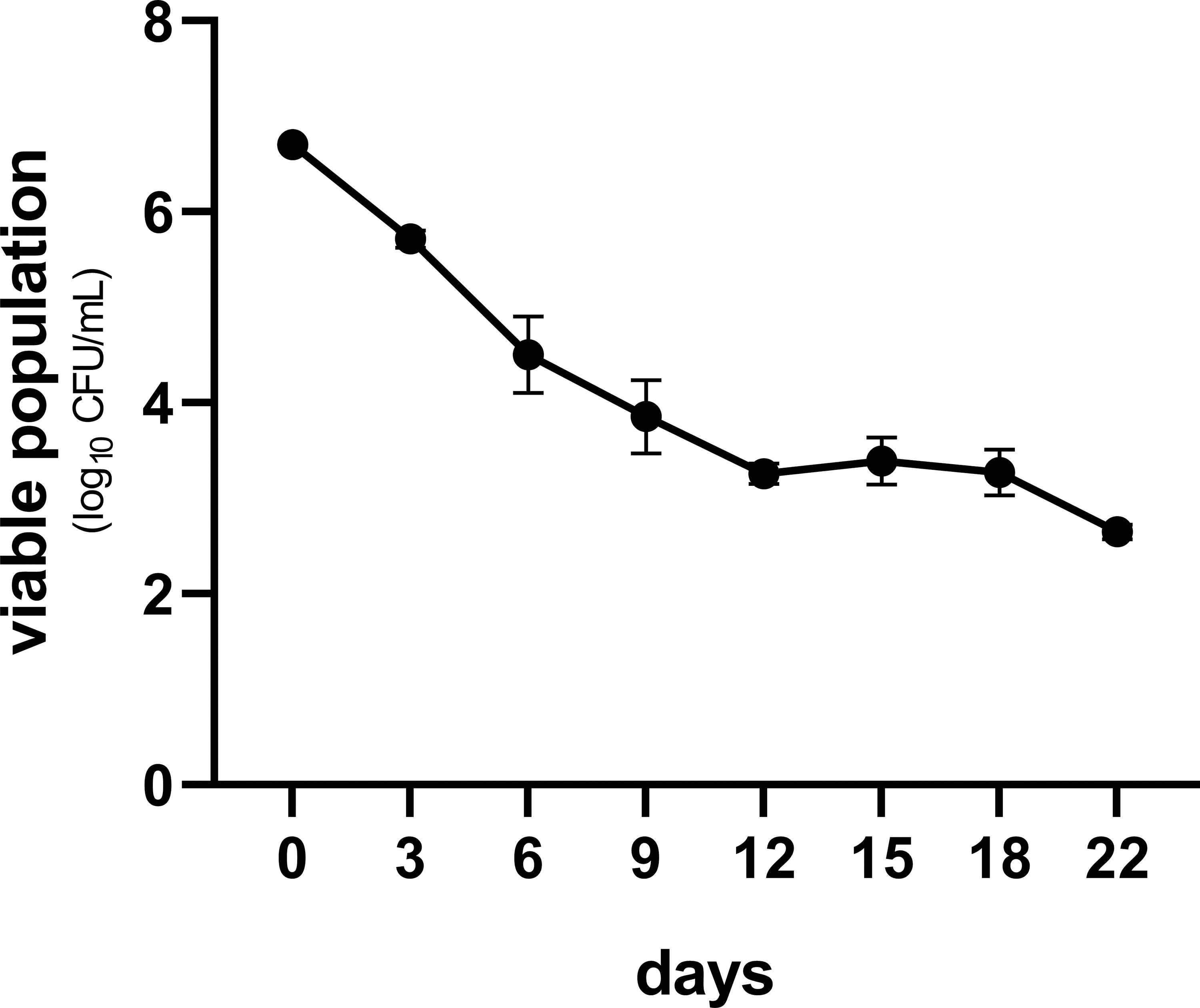
Survival of VBNC Salmonella Enteritidis (S. Enteritidis) under TC treatment. MIC (312.5 µg/mL) of TC against S. Enteritidis was used in the survival assay. The experiment was performed in triplicate. Standard deviations were represented by error bars. TC, trans-cinnamaldehyde; MIC, minimum inhibitory concentration.
Resuscitation of VBNC S. Enteritidis
Successful resuscitation only occurred with 100 mM SP. The treatment of SP yielded a bacterial concentration of 1.29 ± 0.92 log10CFU/mL and 3.28 ± 0.51 log10CFU/mL of VBNC cells after 48 h and 72 h, respectively (Table 2). Catalase and cell-free supernatant of log-phase S. Enteritidis failed to resuscitate the TC-induced VBNC cells.
Resuscitation of VBNC S. Enteritidis Using Different Resuscitation Reagents
“
Intracellular ROS levels of S. Enteritidis
Intracellular level of ROS in VBNC S. Enteritidis was significantly higher than that in culturable cells (p < 0.05) (Table 3). Fluorescence intensity of resuscitated cells with SP treatment showed no significant difference to the culturable cells (Table 3).
ROS Levels of Culturable and VBNC S. Enteritidis after Resuscitation Assays
Different letters indicate significant differences in means ± standard deviations between treatments by one-way ANOVA (p < 0.05).
Resistances of S. Enteritidis to heat, acids, antibiotics, and H2O2
There was no significant difference in thermal resistance between culturable and VBNC cells under 60°C treatment (Fig. 2a). However, under treatment of 75°C, a significantly higher population of VBNC S. Enteritidis was observed compared with the culturable cells (p < 0.05). After the treatment of lactic acid and acetic acid (pH = 3.5), the viable population of VBNC S. Enteritidis remarkably decreased (p < 0.05); in contrast, the reduced populations were not significant in culturable cells (p > 0.05) (Fig. 2b). Besides, after 3% (v/v) H2O2 treatment for 24 h, reduction of culturable cells (2.69 ± 0.10 log10CFU/mL) was greater than that of VBNC (1.61 ± 0.06 log10CFU/mL) (p < 0.01) (Fig. 2c). Enhanced resistances to antibiotics could be observed in Figure 2d. The reduction level of VBNC cells was significantly lower than that of culturable cells (p < 0.05) under treatment of each antibiotic, except LCF.
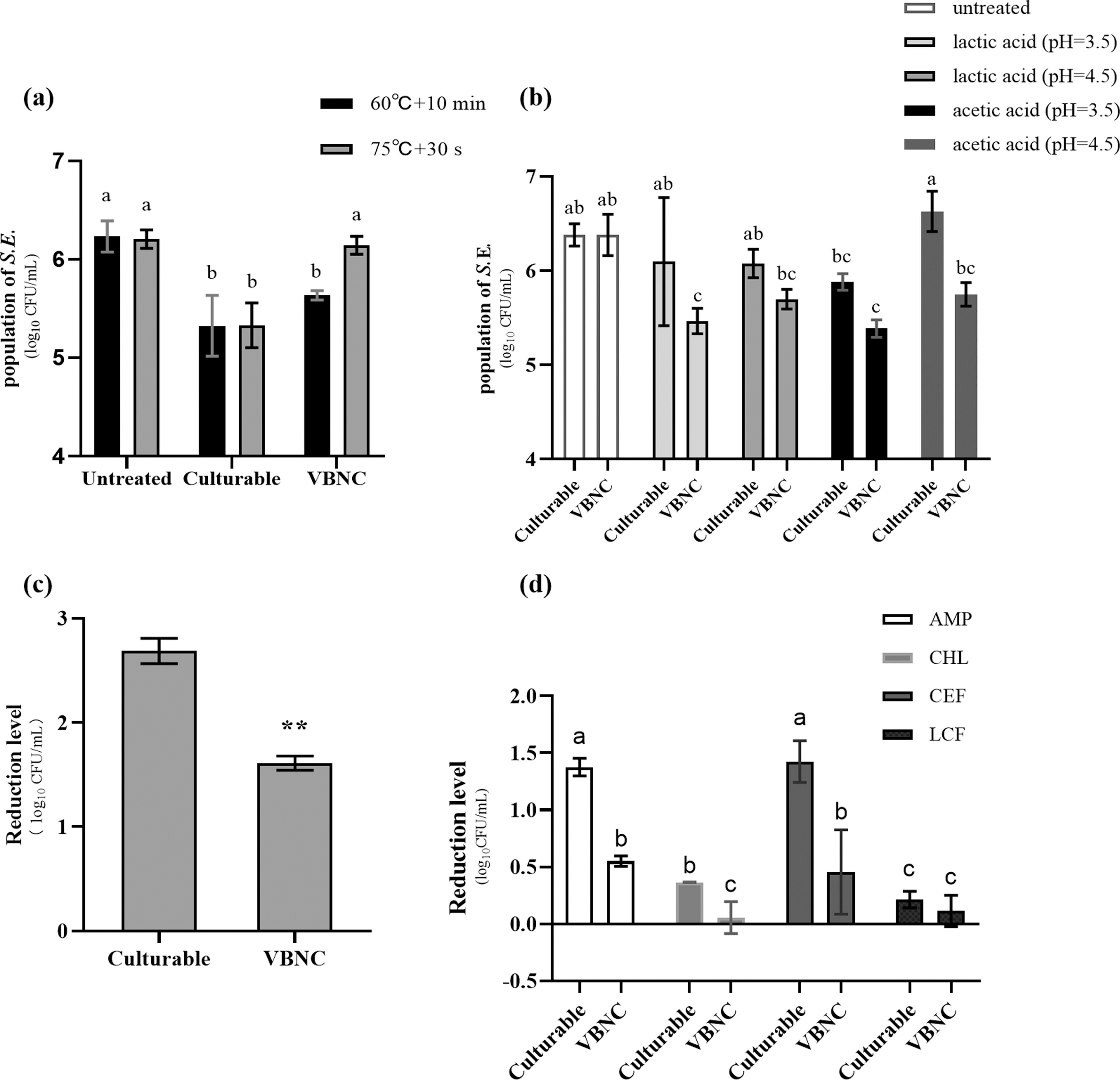
Profiles of resistance between culturable and VBNC S. Enteritidis induced by TC.
Gastrointestinal simulation and cell infectious assay
There was no significant reduction of viable population among culturable, VBNC, and resuscitated S. Enteritidis in the gastric phase (Fig. 3). After a 2-h digestion in the intestinal phase, viable population of VBNC and resuscitated S. Enteritidis was found to decrease by 1.30 ± 0.29 and 1.24 ± 0.48 log10CFU/mL, respectively (p < 0.05).
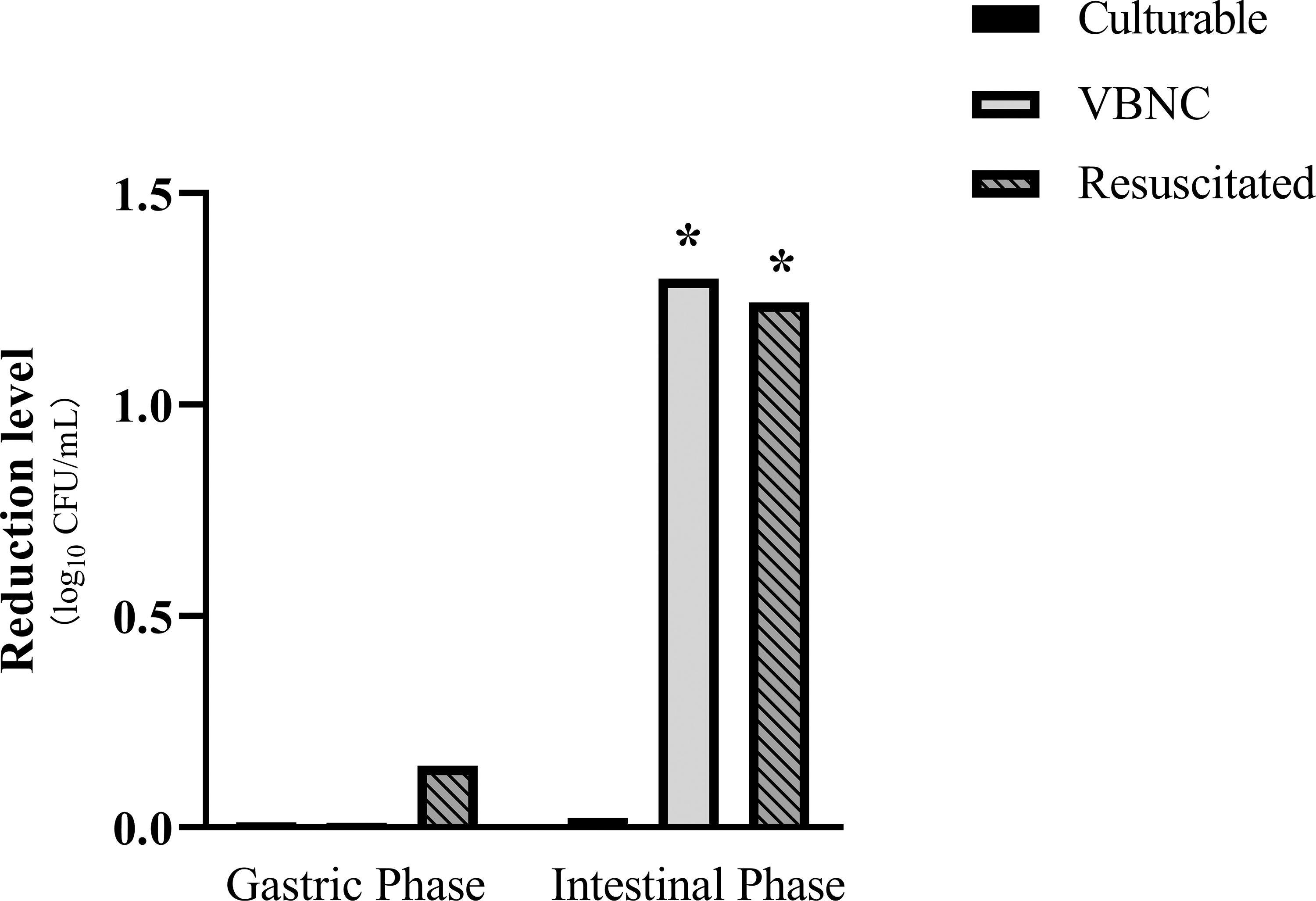
Reduction in levels of viable population in culturable, VBNC, and resuscitated S. Enteritidis under simulated gastrointestinal environment. “*” indicates statistical significance in population reduction (p < 0.05).
Moreover, it showed that VBNC S. Enteritidis maintained adhesive and invasive capacities to Caco-2 cells (Table 4). Infectious capacity of resuscitated S. Enteritidis was significantly weaker than those of the culturable and VBNC cells (p < 0.05).
Adhesion and Invasion Efficiency of S. Enteritidis
Different letters indicate significant differences in means ± standard deviations between treatments by one-way ANOVA (p < 0.05).
Gene expression
Relative expression levels of genes involving antioxidation (katG and sodA), (p)ppGpp synthesis (relA, lon, spoT, and ppx), and protein aggregation (dnaK and grpE) were measured (Fig. 4). All of these genes significantly upregulated with relative fold changes ranging from 12.52–1876.86 folds compared to log-phase S. Enteritidis. Expression of grpE achieved the highest upregulation level with an average fold change of 1876.86. Genes sodA and katG also exhibited high expression levels (217.14 and 53.01 folds, respectively). Relatively low expression levels of relA and spoT were observed in VBNC S. Enteritidis (12.52 folds and 18.37 folds, respectively).
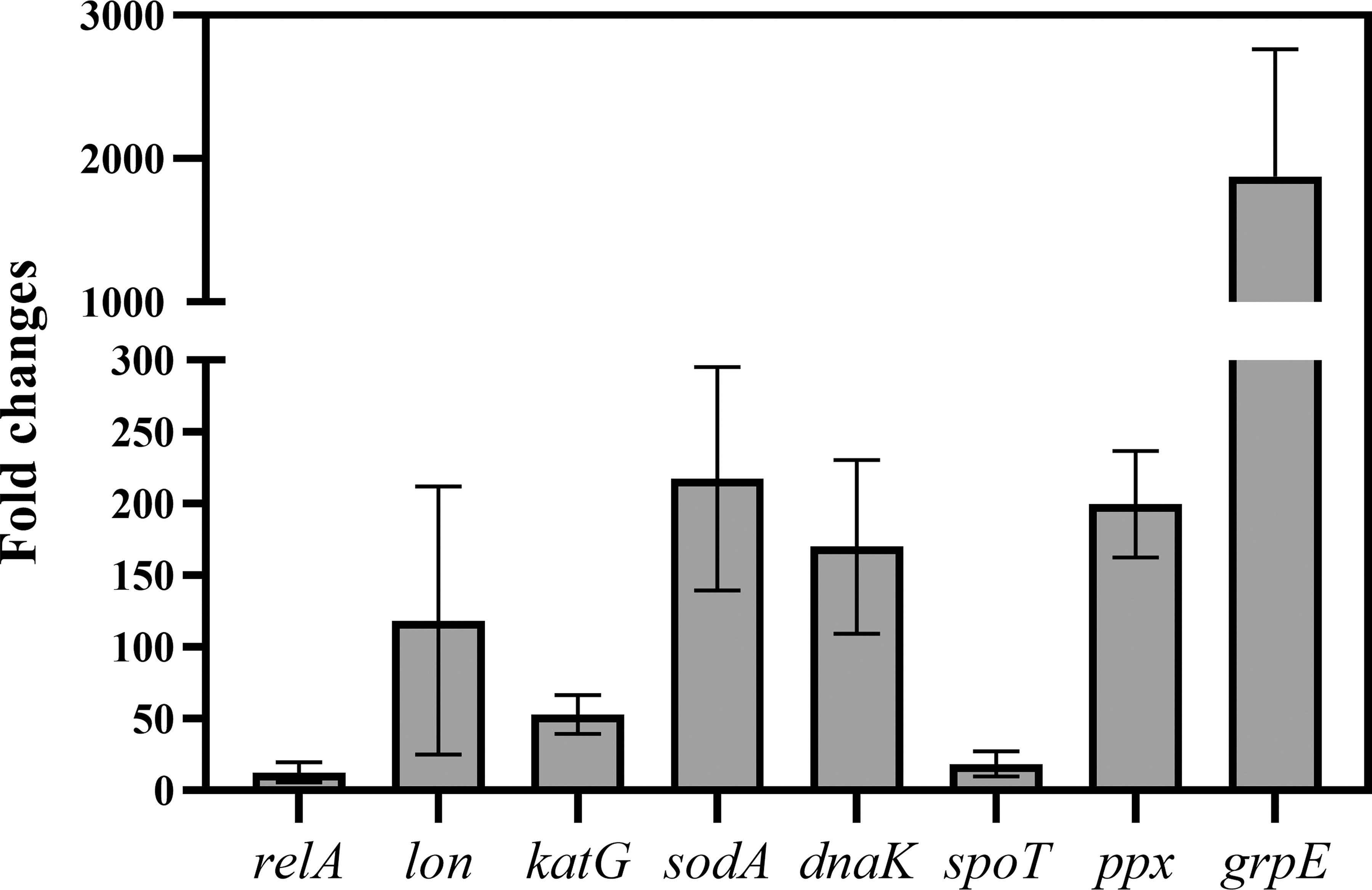
Expression levels of stress-related genes involving heat shock proteins, antioxidation, and
Morphology of S. Enteritidis
In culturable S. Enteritidis, intracellular cytoplasm was found clinging to the cell wall (Fig. 5a and 5b). In contrast, the morphology of VBNC S. Enteritidis transformed to coccoids (Fig. 5c and 5d). The isolation of cytoplasm from cell wall and presence of dark block mass were observed in VBNC S. Enteritidis. Besides, a number of S. Enteritidis cells became fractured after 24-h treatment with TC, while some of them remained intact (Fig. 5c).
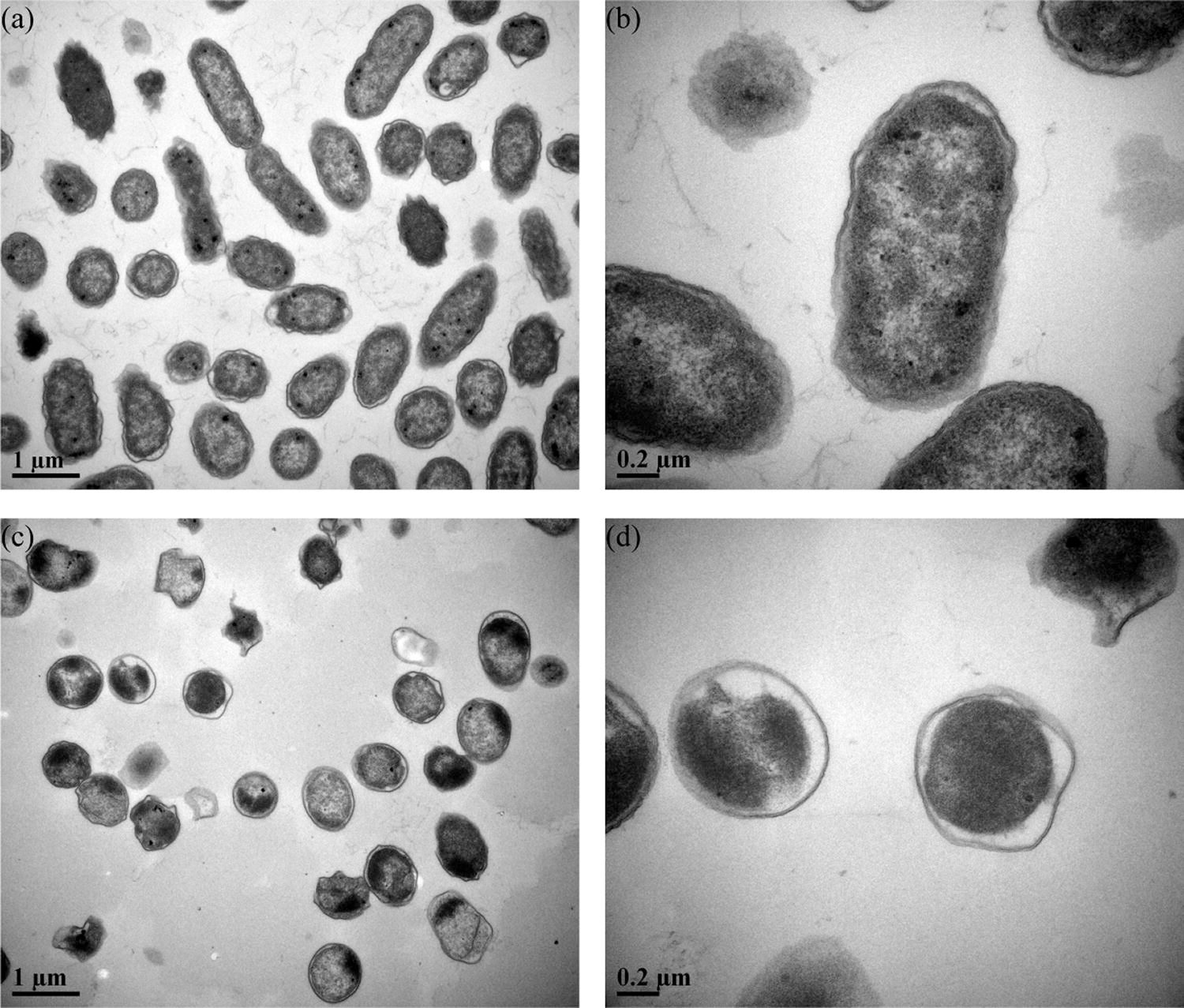
Transmission electron microscope images of culturable S. Enteritidis
Protein aggregation of S. Enteritidis
The SDS-PAGE assay illustrated that protein aggregates in VBNC cells were greater than that of culturable and resuscitated cells (Fig. 6a). Integrated density of protein aggregates in VBNC cells was statistically higher than that of culturable cells and resuscitated cells (Fig. 6b).
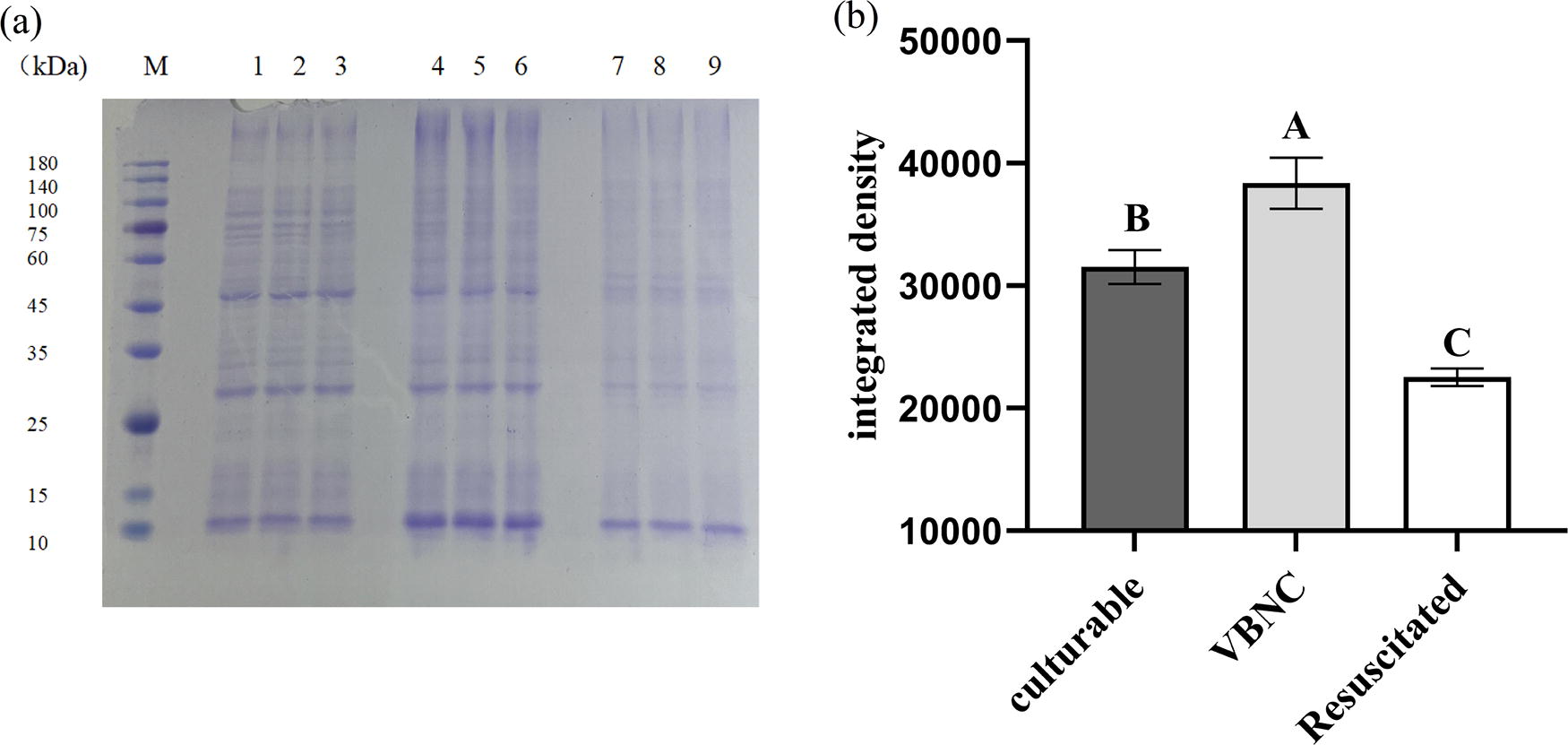
Variation of protein aggregation in culturable, VBNC, and resuscitated S. Enteritidis.
Discussion
Cinnamaldehyde has been effectively applied to inhibit the growth of Salmonella in various types of food products such as chicken (Wang et al., 2023a) and apple jam (Pereira Batista et al., 2023). In this study, we further found that the TC rendered S. Enteritidis into VBNC state, which survived for at least 22 days under TC treatment.
Consistent with studies reporting stress resistance of VBNC bacterial cells (Guillén et al., 2021, Oliveira et al., 2021), VBNC S. Enteritidis induced by TC also presented enhanced heat tolerance and H2O2 resistance (Fig. 2a and 2c). This is also supported by evidence that the stress-related genes were all upregulated in this study (Fig. 4). Heat shock genes (dnaK and groEL) played an important role in protein folding and promotion of proteolysis of misfolded proteins (Kang et al., 2018), which may provide protection against heat stress in S. Enteritidis.
VBNC S. Enteritidis showed increased tolerance to ampicillin, ceftriaxone sodium, and chloramphenicol (Fig. 2d). Enhanced antibiotic resistances have been reported in VBNC pathogenic bacteria such as Mycobacterium smegmatis (Anuchin et al., 2009) and Staphylococcus aureus (Liao et al., 2021), which may be associated with low metabolic activity and accumulation of ROS (Liao et al., 2021), as well as enhanced efflux activity and protein aggregates (Pu et al., 2019, Pu et al., 2016).
It is worth noting that VBNC S. Enteritidis displayed increased susceptibility to acids (Fig. 2b), which seemed inconsistent with previous studies reporting increased tolerance to acid in VBNC S. enterica (Oliveira et al., 2021, Xu et al., 2008). The inconsistency may be due to the accumulation of protein aggregates in VBNC cells, as well as the prolonged exposure time of acid treatment. Prolonged acid treatment may damage proteins through covalent modification and cause misfolding that leads to irreversible aggregation (Schramm et al., 2019). Besides, as cinnamaldehyde can permeate the cell wall and disrupt the cytoplasmic membrane structure (Duan et al., 2022), a synergistic effect against VBNC S. Enteritidis may have occurred when TC was combined with acid treatment, resulting in damage to cell membranes (Huang et al., 2021). These findings indicate that acid treatment can be potentially used as an effective control strategy against VBNC pathogenic bacteria.
Gastrointestinal simulation assays showed that VBNC and resuscitated cells had significantly higher susceptibility to simulated gastrointestinal environment (Fig. 3), which may be attributed to the antimicrobial action of bile salts (Su et al., 2013), as well as the aforementioned acid-induced inhibition effects. Nevertheless, VBNC S. Enteritidis retained adhesive and invasive capacities to Caco-2 cells (Table 4). Similar results were also reported in stressed Salmonella Typhimurium (Chaisowwong et al., 2012).
In this study, successful resuscitation only occurred with treatment of sodium pyruvate, which could resuscitate VBNC Salmonella by scavenging free radical (Liao et al., 2018). Reduced ROS levels of resuscitated S. Enteritidis (Table 3) demonstrated that ROS elimination was of importance for resuscitation. Unexpectedly, catalase and CFS failed to recover the growth of VBNC S. Enteritidis, while successful resuscitation with catalase and CFS has been reported in previous studies (Ayrapetyan et al., 2014; Hamabata et al., 2021).
The accumulation of protein aggregates and increase in ROS levels are signs of deep dormancy in VBNC cells (Fu et al., 2020). The accumulated ROS might lead to instability of protein structures, further causing the aggregates of destroyed proteins; furthermore, intracellular ATP might be exhausted by ROS, hampering protein disaggregation that was regulated by ATP-motivated DnaK and GroEL (Kohanski et al., 2007, Rowe et al., 2020). Instable proteins may hinder the processes of vital metabolic pathways, which may eventually induce the bacteria into a state of dormancy (Bollen et al., 2021, Dewachter et al., 2021). Clearance of cellular protein aggregates may favor recovery of cells from VBNC state (Pu et al., 2019), which was supported by our finding that resuscitated S. Enteritidis presented a significantly decreased level of protein aggregation (Fig. 6).
Conclusion
In this study, VBNC S. Enteritidis induced by TC showed enhanced resistance to heat, antibiotics, and H2O2, while weaker resistance to acid and simulated gastrointestinal environment. A synergistic effect against VBNC S. Enteritidis between TC and acid treatment was observed. The entry of S. Enteritidis into the VBNC state and its resuscitation were accompanied with the accumulation and clearance of ROS and protein aggregates. This research provides a better understanding of the mechanism of VBNC state induced by TC.
Footnotes
Authors’ Contributions
Z.L.: Experiment designing and performing and original article writing. T.X. and Z.Z.: Experiment performing, and validation. Z.L., D.H., Z.M., S.C., and Y.Y.: Data analyzing and writing—review and editing. S.L.: Experiment designing and article editing and writing. H.Z.: Methodology and project supervision. All the authors read and approved the article.
Disclosure Statement
No competing financial interests exist.
Funding Information
This work was financially supported by the National Natural Science Foundation of China (No. 31972044 and No. 32202186).