
Abstract
Carbapenem-resistant Escherichia coli (CREC) is a global threat to public health; therefore, alternative treatment options are urgently needed. Bacteriophages have emerged as promising candidates for combating CREC infections. This study aimed to investigate the genetic basis of phage sensitivity in CREC by evaluating carbapenem resistance among multidrug-resistant (MDR) E. coli isolated in Daegu, South Korea and analyzing their sequence types (STs) with phage susceptibility spectra. Among the 60 MDR E. coli isolates, 80.4% were identified as CREC, with 77.0% demonstrating resistance to imipenem and 66.6% to meropenem. Moreover, 70 lytic E. coli bacteriophages were isolated from hospital sewage water and evaluated against those 60 E. coli isolates. The phages exhibited lytic activity of 33%–60%, with average titers ranging from 5.6 × 1012 to 2.4 × 1013 PFU/mL (Plaque-Forming Unit). Furthermore, multilocus sequence typing (MLST) analysis of the bacterial isolates revealed 14 distinct STs, mostly belonging to ST131, ST410, and ST648. Notably, the phage susceptibility spectra of ST73, ST13003, ST648, ST2311, ST167, ST405, ST607, ST7962, and ST131 were significantly different. Thus, the isolated phages can effectively lyse CREC isolates, particularly those with clinically dominant STs. Conversely, ST410 exhibited a 14.2%–87.14% susceptibility spectrum, whereas ST1139, ST1487, ST10, and ST206 did not lyse, suggesting the presence of more resistant STs. Future studies are warranted to identify the reasons behind this resistance and address it. Ultimately, this study will aid in developing focused treatments to address these pressing global health issues.
Introduction
The widespread use of extended-spectrum β-lactamases (ESBLs) in Enterobacteriaceae has decreased the effectiveness of third-generation cephalosporins, fluoroquinolones, and aminoglycosides in treating ESBL-positive Enterobacter infections. Consequently, carbapenems have become the last effective antimicrobial agents for controlling infections caused by multidrug-resistant Enterobacteriaceae (Chaudhary and Aggarwal, 2004; Pana and Zaoutis, 2018). However, the effectiveness of this treatment regimen is now at risk because of the emergence of carbapenem-resistant isolates. Regrettably, this resistance has given rise to various diseases caused by E. coli, including meningitis, urinary tract infections, pneumonia, sepsis, gastroenteritis, traveler’s diarrhea, neonatal meningitis, wound infections, bacteremia, and pyelonephritis (Briceño et al., 2012; Riley, 2020). The main cause of carbapenem resistance is the expression of ESBL/AmpC enzymes in combination with reduced cellular permeability because of the loss of outer membrane proteins. Isolates exhibiting this resistance mechanism frequently display varying levels of susceptibility to different carbapenems (Meletis, 2015). Furthermore, E. coli can acquire resistance genes from other bacteria and form biofilms that confer protection against antibiotics (Uruén et al., 2020). Antibiotic overuse and inappropriate use can also promote the development of antibiotic-resistant E. coli strains. In addition, exposure to suboptimal doses of antibiotics or disinfectants can lead to the emergence of antibiotic resistance in E. coli (Hogan and Kolter, 2002; Li and Webster, 2018). Isolates with carbapenems-mediated resistance are a critical clinical concern, as multi-institutional outbreaks involving such strains have been reported worldwide (Meletis, 2015). The production of carbapenems by CREC strains grants broad resistance to various antibiotics, making effective treatment significantly challenging. This situation highlights the urgent need for innovative therapeutic approaches and the development of novel antimicrobial agents to combat these infections effectively (Ramírez-Castillo et al., 2023).
Bacteriophages have emerged as a promising alternative to antibiotics for treating antibiotic-resistant E. coli infections (Alisky et al., 1998). Although the therapeutic use of phages was first documented in the early 20th century, it declined sharply with the advent of antibiotics in the 1940s (Hassan et al., 2021). Conversely, there has been renewed interest in phage therapy as a potential alternative to antibiotics in the presence of antibiotic resistance. Clinical approaches for phage therapy have been reported in the United States, Georgia, Poland, and Russia (Cisek et al., 2017; Lin et al., 2017). Phages have a high degree of specificity and selectively target the particular strain of bacteria causing the disease, which makes them more effective than broad-spectrum antibiotics (Lin et al., 2017). Bacteriophages are a natural and safe substitute for antibiotics because they do not contain synthetic substances and have a narrow therapeutic window (Ling et al., 2022). Recently, phage-resistant mutants have become a major concern. To address this issue, phage cocktails were developed to reduce resistant bacteria and maintain high efficacy (Oechslin, 2018). Nevertheless, to ensure the safety and effectiveness of phage cocktails, new bacteriophages must be isolated, and characterization data from experiments conducted both in vitro and in vivo are needed (Gibson et al., 2019).
This study successfully isolated 70 E. coli bacteriophages from hospital sewage water and demonstrated their efficacy against CREC isolates. Through randomly amplified polymorphic DNA-polymerase chain reaction (RAPD-PCR) and MLST analysis, we identified 17 groups of bacteriophages and 14 ST types among 60 MDR E. coli isolates. By comparing the phage lytic spectra and STs results, we identified more effective bacteriophages against CREC and more resistant ST E. coli against antibiotics and bacteriophages. Overall, this study will help advance researchers’ understanding of bacteriophage therapy and present the development of new treatment options for CREC infections.
Materials and Methods
Bacterial strains and culture conditions
Sixty E. coli isolates were collected from the Kyungpook National University Hospital Culture Collection for Pathogens. The isolates were cultured by initially growing them on 5% sheep blood agar medium at 37°C for 24 h, transferred to a brain–heart infusion (BHI), and incubated at 37°C for 24 h. Finally, the isolates were preserved at −70°C using a 50% glycerol stock for future experiments.
Phage isolation, purification, and preparation
Sewage water samples were collected from Kyungpook National University Hospital in Daegu, Republic of Korea. To prepare the phage sample, the E. coli hosts were grown on blood agar. After centrifugation at 12,000 × g for 10 min, the debris and coarse matter from the sewage water were removed. The subsequent supernatant was then passed through serial membrane filters with a pore size of 0.22 μm (Whatman filters, Sigma-Aldrich, St. Louis, USA) to obtain a filtrate that was used to infect E. coli cultures in the early exponential phase (108 CFU mL−1). Infected cultures were incubated at 30°C by shaking (180 rpm) overnight and then kept at 4°C for 48 h. Subsequently, the supernatants were centrifuged again at 12,000 × g for 10 min and filtered through a 0.22mm pore membrane. Bacteriophage titers were determined by the double-layered method, which was purified and stored at −70°C in a glycerol-supplemented medium (Rahman et al., 2011).
Host range determination
The host range of the isolated phages was determined through spot testing with modifications based on the method described by Shukho Kim (Kim et al., 2022). A 15 µL of phage stock (∼1013 PFU/mL) was added to the top of each bacterial lawn representing 60 different E. coli. Following overnight incubation at 37°C, the plates were examined for the formation of a clear lytic zone. The presence of a clear zone indicated bacterial lysis by the phages (+), whereas the absence of a clear lytic zone indicated the absence of lysis (−). The tests were conducted in triplicate to ensure the accuracy of the results.
Phage DNA preparation and subtyping of isolated phages by the RAPD-PCR
Using PEG-8000 precipitation, phage DNA was extracted from a high-titer phage lysate by the guidelines provided by Dziuginta Jakočiune (2018). In addition, the RAPD analysis used short PCR primers with random sequences, typically ranging from 8 to 15 nucleotides, as described by Gutiérrez et al. (2011) (Supplementary Data S1—Procedures).
Antimicrobial susceptibility test (AST)
The antibiotic susceptibility profile of the 60 E. coli isolates was assessed against 21 distinct antibiotics across 10 families. To assess the minimum inhibitory concentration, antibiotic disks (10 µg) and culture materials were purchased from Becton, Dickinson and Company (https://www.bd.com). E. coli ATCC25922 was used as a control strain. For each test, three biological replicates were performed per strain in duplicate. Following Clinical & Laboratory Standards Institute guidelines, isolates were classified as resistant, intermediate, and susceptible (Weinstein and Lewis, 2020).
MLST
To identify different ST types of MDR E. coli, the protocols outlined by Tartof et al. (2005) were followed. To achieve this, we used a method of amplifying and sequencing seven housekeeping genes (adk, fumC, gyrB, icd, mdh, purA, and recA) from each isolate using separate PCR reactions. The PCR products were sequenced using a conventional Sanger method at Solgent Co., Ltd. (South Korea) and an ABI 3730XL DNA Analyzer (Applied Biosystems, Foster City, CA, USA). To determine the MLST type, sequence data were imported into the E. coli MLST database (https://pubmlst.org/organisms/escherichia-spp) (Clermont et al., 2011).
Electron microscopy analysis of bacteriophages
Bacteriophage morphology was examined by transmission electron microscopy (TEM; Zeiss EM902) at 75 kV. Virus samples were prepared for electron microscopy using carbon-shadowed palladium-coated grids. For 2 min, samples were stained with 2% uranyl acetate (pH 4.0) (Ackermann, 2012).
Results
Antibiotic resistance profile of clinically isolated E. coli strains
As shown in Supplementary Table S1, the AST results revealed that all 60 E. coli isolates exhibited a multidrug resistance, indicating an alarming 100% prevalence of MDR (n = 60). Among these, 48 (80.4%) isolates were identified as CREC, highlighting the extent of carbapenem resistance. The antibiotic resistance and susceptibility patterns of 48 CREC isolates were investigated, and the results revealed significant resistance to cephalosporins, specifically cefazolin (100%, 33/33), ceftazidime (90.4%, 38/42), cefoxitin (86.1%, 31/36), ceftriaxone (100%, 8/8), cefotaxime (96.8%, 31/32), and cefepime (84.0%, 37/44). The monobactam family also exhibited notable resistance, with aztreonam showing 78.5% resistance (33/42). Aminopenicillins also showed resistance, with amoxicillin (92%, 23/25) and ampicillin (100%, 33/33) demonstrating high resistance rates. Carbapenem-resistant strains were observed in imipenem (78.7%, 37/47) and meropenem (66.6%, 32/48). The isolates also showed high fluoroquinolone resistance, with 85.3% (33/41) to ciprofloxacin and 93.3% (14/15) to levofloxacin. The aminoglycoside family showed resistance to amikacin (17%, 7/41), tobramycin (100%, 1/1), and gentamicin (56%, 23/41). In addition, the penicillin family resisted piperacillin (100%, 1/1), and the sulphonamide-trimethoprim family resisted trimethoprim (88.4%, 23/26). ESBL was detected in 28% of the isolates (8/28). Finally, E. coli isolates also resisted the tetracycline and oxazolidinone (linezolid) (Fig. 1).
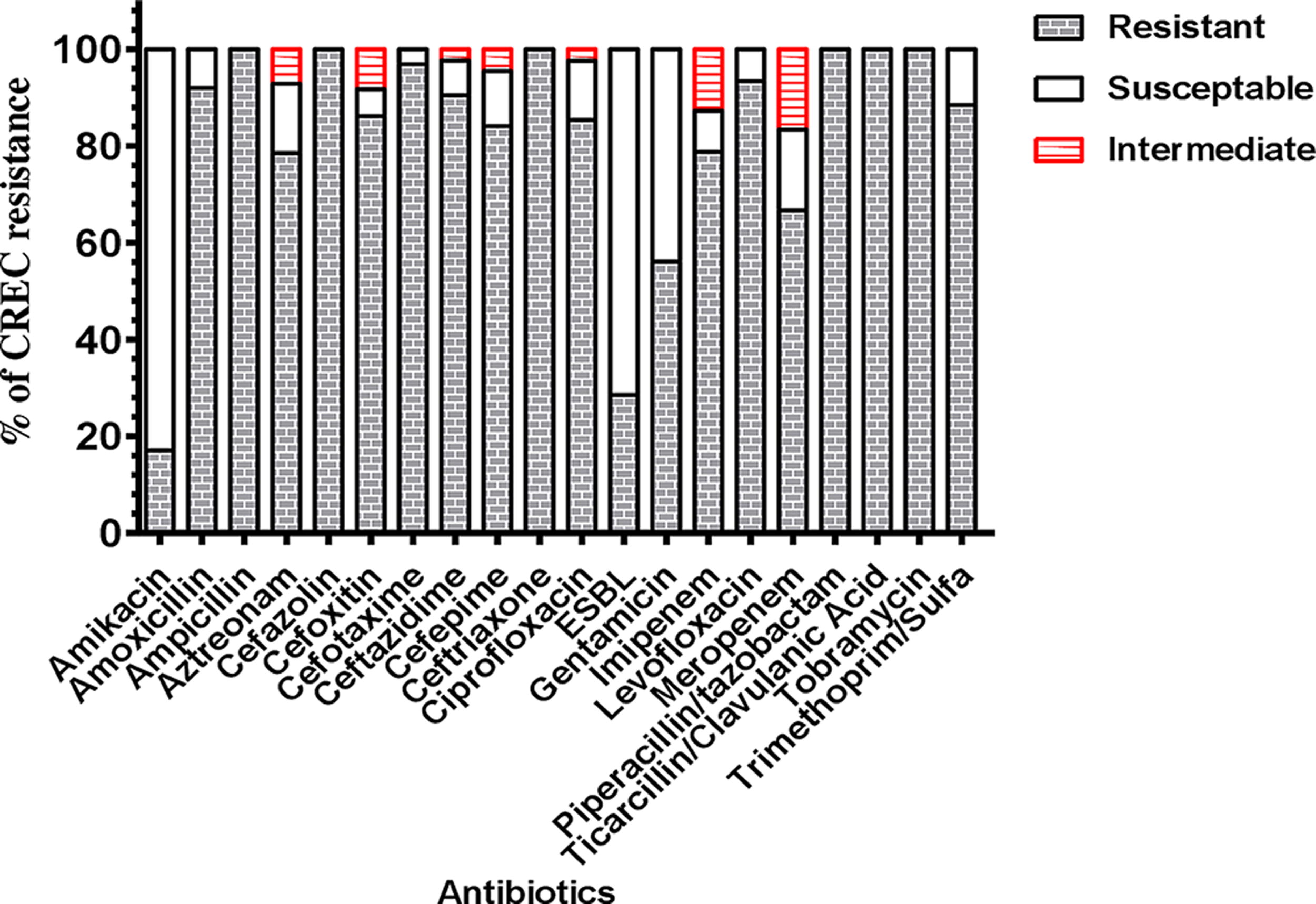
The antibiotic % resistance ratios of 48 carbapenem-resistant E. coli reveal varying resistance levels. % Resistance indicates the proportion of isolates that resist each antibiotic.
Bacteriophage isolation, purification, host range determination, and susceptibility testing
Seventy bacteriophages were isolated from the hospital sewage water using various E. coli as host strains. Following purification and propagation, the phage titers ranged from 1012 to 1013 PFU/mL. The host range of each bacteriophage was determined against 60 MDR E. coli isolates (Fig. 2), and the susceptibility percentage of each bacteriophage against these isolates was determined to range from 33% to 60% (Supplementary Fig. S1). This study found that φ = EC.W1-1, φ = EC.W2-1, φ = EC. W2-6, φ = EC.W8-1, and φ = EC.W15-4 showed lytic activity against Salmonella enterica, whereas φ = EC.W1-9, φ = EC.W2-1, φ = EC.W13-3, and φ = W15-4 displayed lytic activity against Klebsiella pneumonia (Supplementary Table S2).

Correlation between sequence types and phage lytic spectra in 60 multidrug-resistant E. coli. This study examined lytic spectrum of bacteriophages lysing E. coli isolates. Seventy phages were isolated and purified using E. coli isolates as hosts (KBN10P04004, KBN10P03040, KBN10P07288, and ATCC25922). The RAPD-PCR profiles of the phages were analyzed, excluding those with identical sizes. This analysis resulted in the identification of 17 phage classes and subclasses with varying sizes. The black columns in the figure indicate confirmed lysis, signifying effective kill of E. coli strains by these phages. The term “lytic spectrum” of a bacteriophage refers to its range of lytic abilities against different E. coli isolates. For MLST sequencing, seven housekeeping genes (adk, fumC, gyrB, icd, mdh, purA, and recA) were used, and ST were determined using the PubMLST database (https://pubmlst.org). Finally, a comparative analysis was conducted between the phage lytic spectra and ST types of E. coli.
RAPD-PCR
RAPD-PCR revealed that the 70 isolated bacteriophages were arranged into 17 different groups based on their band patterns. Each group showed distinct band patterns between 100 and 2,000 bp, revealing significant genetic diversity was found among them. To obtain more accurate phage groups, 70 bacteriophages were analyzed by RAPD-PCR and host spectrum results, and 12 groups were selected for further study (Supplementary Fig. S2).
Morphological analysis of phages
Phage plaques were assessed by culturing them on a BHA plate covered with an E. coli bacterial lawn. After 24 h of incubation at 37°C, transparent halos of different sizes ranging from 1 to 10 mm were observed. To further investigate the 12 phages, TEM was used (Fig. 3). Based on this analysis, all may belong to the order Caudovirales with two morphotypes. The majority, accounting for 66.6% (8/12), looked like under the Straboviridae family, whereas 33.3% (4/12) were observed as Podovirdae. Straboviridae isolates showed different tail lengths varying between 84 ± 3 nm and 120 ± 5 nm and head widths between 100 ± 4 and 115± 5 nm. Furthermore, Podoviridae heads have a diameter of 58 ± 5 to 60 ± 4 nm, and the tail has a dimension of 5 ± ∼10 ± 2 nm (Table 1).
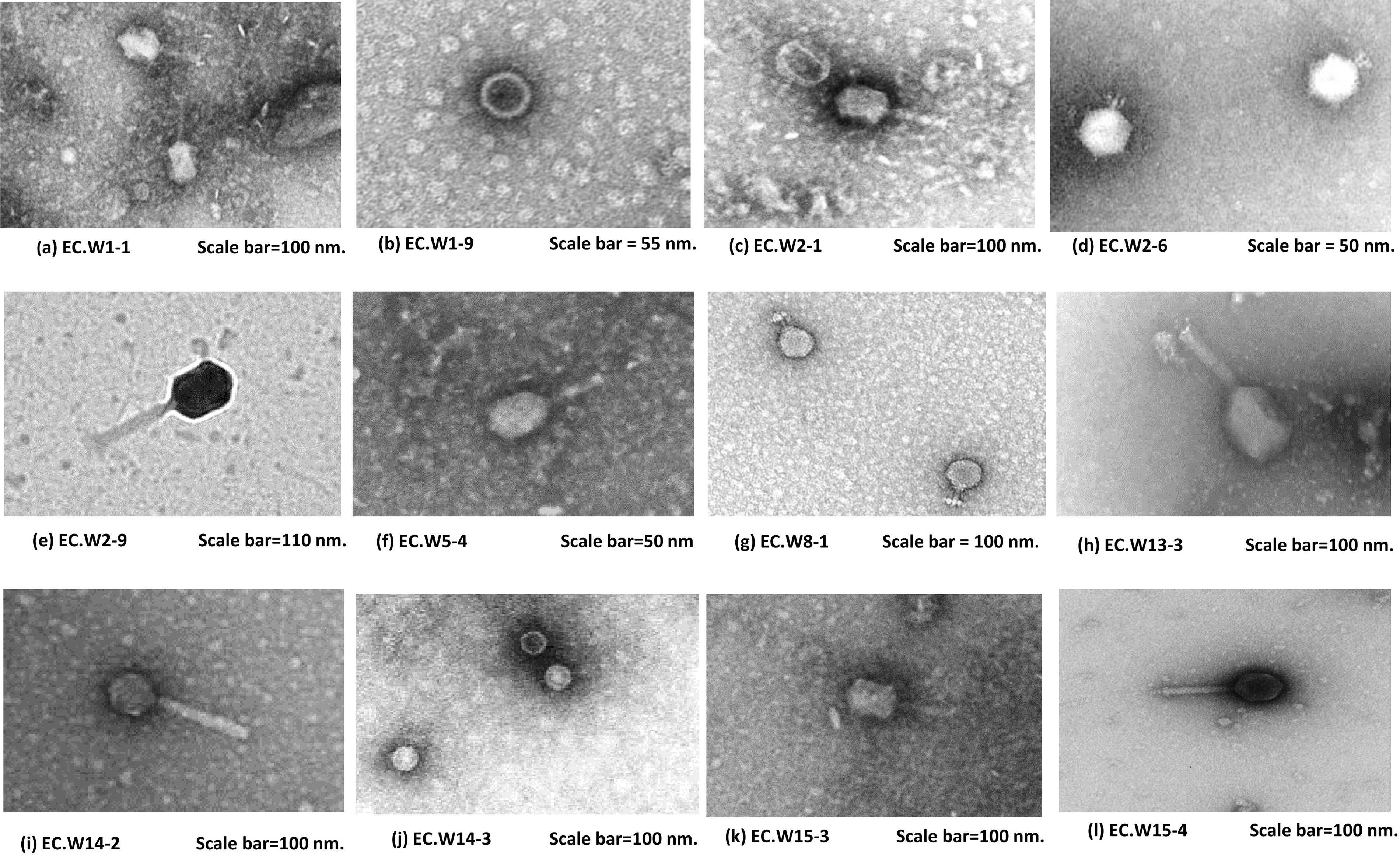
Transmission electron microscopy images of Escherichia coli phages showing the following classifications based on their distinct morphologies: (1) Podoviridae morphology: b, d, g, and j; and (2) Straboviridae morphology: a, c, e, f, h, i, k and l.
The Morphology of Escherichia coli Phages via Transmission Electron Microscopy. The Phages Are Classified into Three Families Based on Their Distinct Morphologies
MLST analysis of E. coli isolates
The 60 E. coli isolates contained a diverse range of STs. Fourteen distinct STs were detected with varying prevalence. The most prevalent STs were ST131 (30%, 18/60) and ST410 (42.3%, 26/60) (Supplementary Fig. S4). Interestingly, some STs were significantly carbapenem-resistant. ST2311, ST167, ST607, ST13003, ST1139, ST1487, ST10, and ST206 each harbored 100% CREC isolates within their respective groups. Further analysis revealed that ST131 contained 77.7% (14/18) of CREC, whereas ST410 exhibited a high proportion of CREC, with 96.15% (25/26) displaying carbapenem resistance. ST648 and ST7962 were intermediate, with approximately 50% of the isolates in each ST demonstrating carbapenem resistance (Table 2).
Multilocus Sequence Types and % Carbapenem Antibiotic Resistance Among 60 E. coli Isolates and % Lysis Activity of 70 Isolated E. coli Bacteriophages in Each ST Type Group
CREC, carbapenem-resistant Escherichia coli; E. coli, Escherichia coli; MDR, multidrug resistant; ST, sequence type.
Discussion
The resistant of CREC to many commonly used antibiotics is a growing issue because of the emergence of the blaNDM-5 gene in E. coli strains (Duong et al., 2022). It can spread quickly through travel or local transmission, causing concern for future infections that could resist last-line treatments. It is an urgent issue in many countries, causing deaths because of antimicrobial resistance. Promoting responsible antibiotic use and effective infection control is crucial to preserving existing antibiotic efficacy (Xu et al., 2016; Mills and Lee, 2019). The study investigated the use of bacteriophages as a potential therapeutic approach against CREC strains, focusing on the correlation between high-risk STs and the effectiveness of phage-mediated lysis. 70 E. coli phages were isolated from hospital sewage water, and 17 classes were selected based on their lytic efficiency and host range. A specific set of MDR E. coli isolates were collected, and 81.6% CREC was identified from 60 MDR isolates. The phages were characterized and tested for their ability to lyse clinical isolates, RAPD-PCR results, and TEM analysis.
Most CRECs were found in ST410, ST167 ST648, and ST131 (Bhattacharjee et al., 2023). The majority of CREC from South Korea were ST131 and ST410, which is concerning because they have been widely distributed (Kim et al., 2016; Baek et al., 2019). In this study, the MLST sequence revealed that most of the E. coli isolates contain ST410 (42.33%, 26/60) and ST131 (30%, 18/60) among the 60 E. coli isolates. ST410 and ST131 are STs of E. coli strains that have been associated with a high degree of pathogenicity and antibiotic resistance (Adler et al., 2015). ST410 is particularly concerning because of its ability to cause infections in humans and animals, leading to potential cross-species transmission.ST410 E. coli strains are frequently associated with urinary tract infections, sepsis, and meningitis, with a high mortality risk among infected individuals (Mahazu et al., 2021). Similarly, ST131 E. coli strains are known for their high levels of antibiotic resistance and pathogenicity, particularly in urinary tract infections and bloodstream infections. These strains have been found to harbor the CTX-M-15 extended-spectrum beta-lactamase gene, which confers resistance to many beta-lactam antibiotics (Ogbolu et al., 2018). The ability of ST410 and ST131 E. coli strains to develop antibiotic resistance is a major concern because this can limit treatment options for infected individuals. In this study, ST410 (96.1%) and ST131 (77.7%) identified carbapenem resistance. The study evaluated the effectiveness of phage therapy against antibiotic-resistant E. coli isolates, including ST410 and ST131. Seventy isolated bacteriophages were applied against 26 ST410 E. coli and 18 ST131 E. coli isolates. Consequently, 50% (13/26) of the ST410 E. coli were susceptible to lysis by the phages, whereas 100% (18/18) of the ST131 E. coli were lysed. The individual phage lysis ability was 14.2%–87.4% for ST410 and 77.7% for ST131. Phage therapy, a selective use of bacteriophages to destroy CREC bacteria, has been shown to be a valuable solution to combat antibiotic resistance (Broncano-Lavado et al., 2021). In this study, the antibiotic-resistant patterns of all STs were almost MDR. Phage non-susceptible pattern of ST410 exhibited 50% lysis to the isolated bacteriophages. Moreover, other isolated bacteriophages were ineffective against ST10 (1/1), ST206 (1/1), ST1139 (1/1), and ST1487 (1/1), which exhibited lysis. By contrast, this study observed complete lysis (100%) of ST131 (18/18), ST73 (1/1), STs648 (4/4), ST405 (1/1), ST607 (1/1), ST13003 (1/1), ST2311 (1/1), ST7962 (1/1), and ST167 (1/1) (Fig. 2 and Supplementary Fig. S4). Despite having multiple hosts, phages were successfully isolated from only isolates ST73, ST648, and ST131 of E. coli. Moreover, the phage lytic spectrum has been correlated with 14 distinct STs, including 60 isolates of E. coli. Interestingly, phages isolated from ST73, ST131, and ST648 demonstrated more significant lytic activity against the same ST than phages obtained from different STs. Specifically, if a phage can lyse an isolate belonging to a specific ST, it may have the potential to lyse other isolates of the same ST (Yamamura et al., 2022).
Understanding why some STs of E. coli are not lysed by these bacteriophages may require further research. However, these isolates may have evolved strategies to resist bacteriophages, including changes in surface receptors, phage-degrading enzymes, or immune defense activation (Egido et al., 2022). The effectiveness of phages against specific strains of E. coli depends on their ability to attach to their surface receptors. (Teklemariam et al., 2023). In addition, phage adsorption can be inhibited by differences in bacterial surface structures such as lipopolysaccharides (Chapartegui-González et al., 2020). Conversely, bacteriophages may become inactive because of factors such as environmental conditions, antibiotic exposure, or host defense (Torres-Barceló, 2018). In addition, some bacteriophages have a limited host range and can only infect certain STs or species of bacteria. If the ST of E. coli is not within the host range of the phage, it will not be more effective. The effectiveness of phages is also dependent on their concentration or titer. If the titer is too low, they may not be able to infect or lyse the bacteria effectively (Torres-Barceló, 2018; Hyman, 2019). However, we carefully determined the host ranges of bacteriophages using plaque and spot tests. The ranges were between 1013 and 1015 (PFU/ml). Furthermore, host bacteria can have defense mechanisms that prevent phages from penetrating their cell walls or membranes, such as efflux pumps or restriction-modification systems (Smith et al., 2023).
In this study, by comparing RAPD-PCR and host spectrum results, we classified 70 bacteriophages into 17 categories and reduced their numbers. In contrast, PubMLST is a bacterial molecular-typing database with sequence data for 100+ microorganisms, which are crucial for clinical epidemiology (Jolley et al., 2018). Considering these circumstances, classifying strains and estimating endemic clones using the method described in this study and not by the receptor recognized by phages is reasonable. Future studies should consider the whole genome sequence of phages and identify their genomic information with phage lysis experiments using phage cocktails and phage antibiotic synergistic methods for phage-resistant ST E. coli. Finally, MLST data with additional typing schemes for index receptors recognized by phages are needed.
Conclusion
This study highlights the urgent need for alternative treatment options for CREC, a global public health threat. To provide valuable understanding for the development of CREC therapies, this study extensively isolated 70 lytic E. coli bacteriophages and identified distinct E. coli STs that are predominant in Korea. The varying levels of phage resistance observed among STs highlight the importance of understanding the genetic diversity and resistance patterns of CREC isolates. Although bacteriophage therapy is still experimental, further research is crucial to determine its safety and efficacy in human patients. These significant findings pave the way for potential advancements in treating CREC infections, offering hope for improved patient outcomes and public health preservation.
Footnotes
Disclosure Statement
The authors declare no conflict of interest.
Authors’ Contributions
M.S. and S.K. conceived and designed the experiments. M.S. conducted all the experiments. S.K., M.S.S., and J.K. interpreted the data. J.K. coordinated and supervised this study. M.S., S.K., Y.J.C., B.K., R.H.D., M.S.S., and J.K. analyzed the data and prepared the article. All authors contributed to the article and approved the submitted version.
Funding Information
This research was supported by a grant from the Korea Disease Control and Prevention Agency (Grant No. 2022-ER2202-00).
Supplementary Material
Supplementary Data S1
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.