
Abstract
The spread of extended-spectrum beta-lactamase–producing Enterobacterales (ESBL-E) in communities has been facilitated by direct contact with humans or animal carriers and insects such as houseflies. Houseflies may have an increased risk of human exposure to antimicrobial-resistant pathogens, including ESBL-E. In this study, we determined the presence of ESBL-E in houseflies and ready-to-eat food samples from selected food vending sites in five health care facilities in Greater Accra, Ghana. Sixty (60) food vending sites were included in this study. Flies (10) were pooled, and food (5 g) was macerated and homogenized aseptically in brain-heart infusion agar. Suspensions were cultured on MacConkey agar supplemented with 4 µg/mL cefotaxime for ∼18 h. Phenotypic and molecular characterization of ESBL-E was performed using a combination disk diffusion method, multiplex polymerase chain reaction, and direct nucleotide sequencing. Of the 60 food vending sites, 62% (37) of the food vendors had flies within their premises that carried ESBL-E, and 15% (9) of the food samples collected from the vendors were contaminated with ESBL-E. The most predominant ESBL-producing Enterobacterales from housefly samples was Proteus vulgaris (54%) and that of food samples were P. vulgaris (22.5%) and Escherichia coli (22.5%). All Enterobacterales from flies and food samples harbored an ESBL gene with blaCTX-M, mostly blaCTX-M-15, being the predominant gene and gene type, respectively. Houseflies may be potential contributors to the spread of ESBL-producing Enterobacterales in Ghana.
Keywords
Introduction
Extended-spectrum beta-lactamases (ESBL) are enzymes produced by Enterobacterales that can hydrolyze most beta-lactam antibiotics, including penicillin, broad-spectrum cephalosporins, and monobactams, except for carbapenems (Bush et al., 1995; Bush and Bradford, 2020; Bush and Jacoby, 2010). Extended-spectrum beta-lactamase–producing Enterobacterales (ESBL-E) often exhibit multidrug resistance, affecting second-line antibiotic therapy for more common hospital- and community-acquired infections, resulting in increased hospital costs, morbidity, and mortality rates (Joo et al., 2018; Obeng-Nkrumah et al., 2023). The dissemination of ESBL-E within the community is facilitated by direct contact with human or animal carriers and through vectors such as houseflies (Egbule et al., 2020; Songe et al., 2017). Houseflies (Musca domestica L.), ubiquitous insects found worldwide, are carriers of antimicrobial-resistant (AMR) pathogens such as ESBL-E (Sivakumar et al., 2021). Environmental and sanitary conditions influence the prevalence of ESBL-E in houseflies in a specific location (Browne and Kerr, 1986; Fisher et al., 2017; Gupta et al., 2012; Yamamoto and Jensen, 1967). In Ghana, hospitals within the Greater Accra Region have reported ESBL production in ∼50% of Enterobacterales infections (Eibach et al., 2016; Obeng-Nkrumah et al., 2013; Owusu et al., 2023).
Literature suggests that sanitary conditions surrounding food vendors in hospitals can directly impact the prevalence of antibiotic-resistant pathogens in the hospital environment (Dela et al., 2022). Houseflies can readily move between potential contamination sources, including animal and human wastes, and food preparation areas (Goulson et al., 1999; Tufa et al., 2020). This can potentially increase the risk of spreading antibiotic-resistant organisms including ESBL-E. Despite their potential role in disease transmission, the contribution of houseflies to the spread of bacterial infections remains unclear. This study aimed to address this gap in knowledge by investigating the presence of ESBL-E in houseflies collected from food vendor sites within five health care facilities in the Greater Accra Region of Ghana.
Materials and Methods
Ethical clearance
Ethical approval was obtained from the Ethics and Protocol Review Committee of the School of Biomedical and Allied Health Sciences (SBAH) before the study was conducted (Ethics Identification Number: SBAHS—MD./10484603/AA/5A/2016–2017). Food vendors were provided with a clear explanation of the procedure, for their participation. Permission (oral consent) was also sought from food vendors at the various sites.
Study design and setting
This cross-sectional study was conducted between January 2017 and August 2017. Sixty (60) food vendor sites were selected by systematic sampling from two hospitals, Health Care Facility 1 (n = 20) and Health Care Facility 2 (n = 6), and three polyclinics, Health Care Facility 3 (n = 10), Health Care Facility 4 (n = 14), and Health Care Facility 5 (n = 10), in the Greater Accra Region, Ghana (Table 1). Only food vendors selling one type of food were included in the study to control for homogeneity. Each site was visited once for sampling food items and houseflies, with no follow-up visits. Sampling was performed at each site between 8:00 a.m. and 6:00 p.m. to minimize temporal variability in the outputs. A total of 60 days was used for sampling. Houseflies were collected within a circular area of 50 m2, centered around the food vendor site. Flies were swatted randomly at a rate of one per hour using a sterile fly swatter over a 10-h sampling period. Ten flies (regardless of species) were collected from each vendor site and pooled into a single sample. The collected flies were placed in sterile labeled containers and promptly transported to the laboratory. At each site, ∼0.5 g of food was collected every hour over 10 h and pooled to create a total sample of 5 g per site. Food samples (5 g) were placed in sterile containers and transported to the laboratory (Fig. 1).
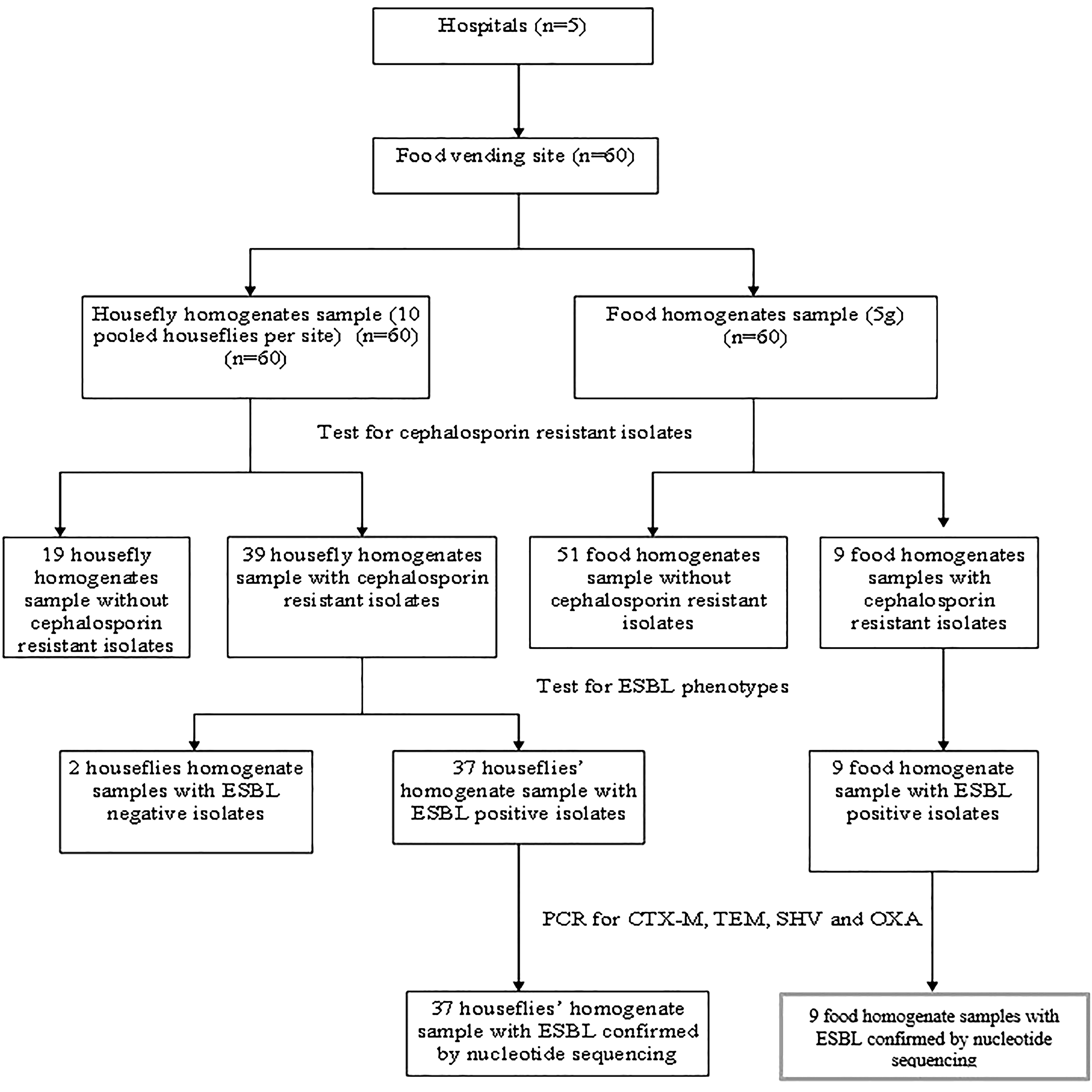
Flowchart for data collection and summary of study outcome.
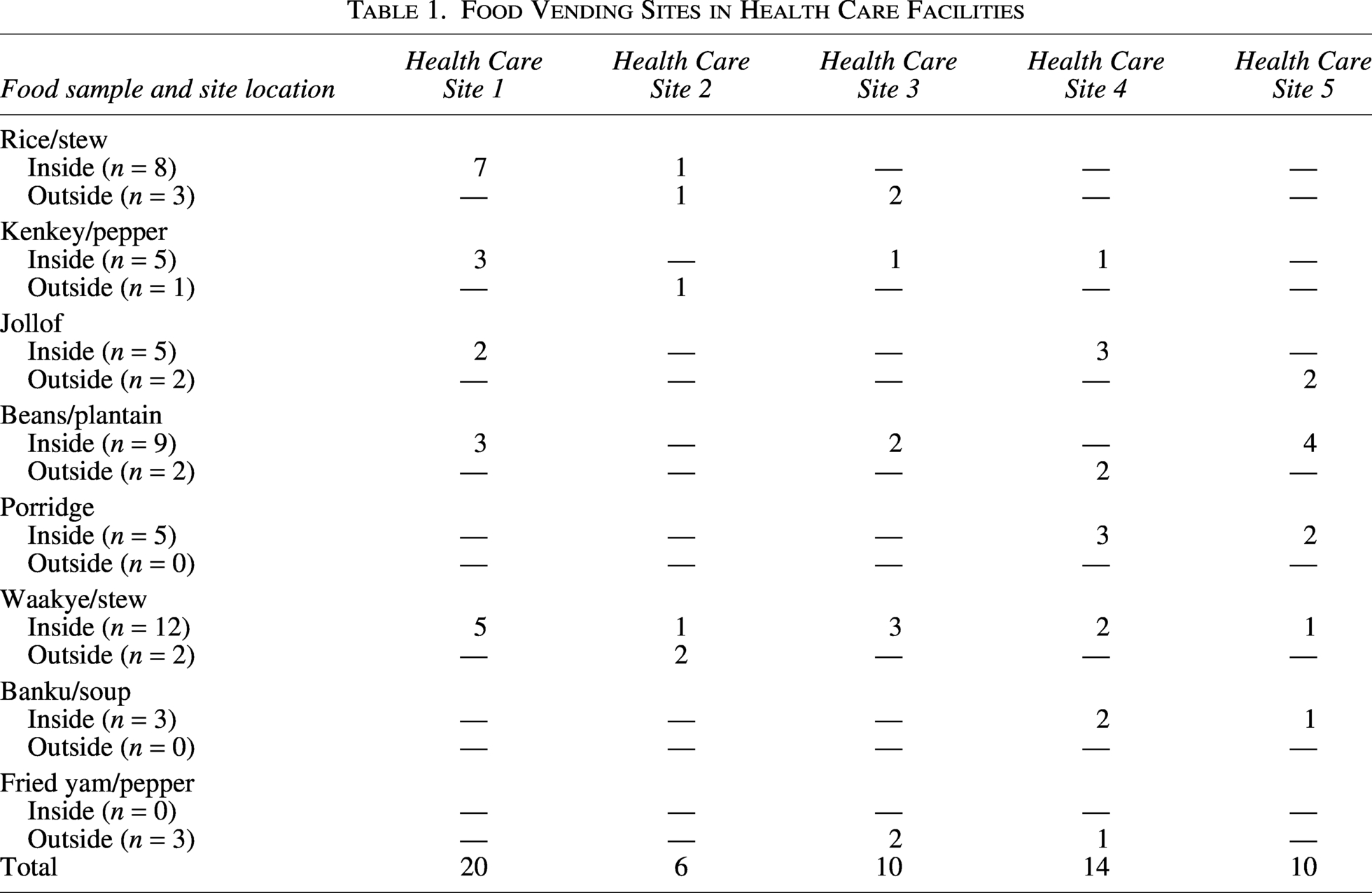
Food Vending Sites in Health Care Facilities
Sample processing, culturing, and screening
Each sample, 10 houseflies per food vendor site, and 5 g of each food sample collected were soaked in 5 mL and 20 mL, respectively, of brain-heart infusion (BHI) broth in sterile containers, vortexed vigorously for 5 min, and incubated for ∼18–24 h at 35–37°C. Subsequently, insects and food samples were macerated using a sterile rod and vortexed vigorously for 5 min. The resulting suspension of homogenized houseflies in 5 mL of BHI broth and a homogenized food sample in 20 mL of BHI broth was taken as different whole homogenate samples. A loop full of the suspension from each sample was screened for third-generation cephalosporin-resistant Enterobacterales (CrE) by inoculating onto MacConkey agar plates supplemented with cefotaxime (CTX, 4 µg/mL) (MAST) and incubated overnight at 35–37°C for 18–24 h. From each culture plate, distinct morphological phenotypes of Enterobacterales colonies on agar media—identified based on colonial morphology and biochemical tests—were considered positive for CrE. Single discrete colonies were subcultured on MAST for purity.
Identification of Enterobacterales
Each distinct morphological phenotype (representative colonies of pure isolates) was identified to the species level using biochemical test kits (Minibact-E®; SSI Diagnostica, Hillerød, Denmark) according to the manufacturer’s guidelines, and six colonies of each species were tested against 30 µg cefotaxime Kirby-Bauer disk diffusion to confirm the CrE status.
Characterization of ESBL phenotypes
Enterobacterales that were positive for CrE were phenotypically confirmed to produce ESBL. ESBL production was detected using the combination disk diffusion method with cefotaxime (30 μg) and ceftazidime (30 μg) alone and in combination with clavulanic acid (10 μg) using Neo-Sensitabs (Rosco Diagnostica, Taastrup, Denmark), according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI, 2017). Isolates that demonstrated a clavulanic acid effect, as defined by a zone difference of ≥5 mm between the single and combination disks for cefotaxime and/or ceftazidime were regarded as positive for ESBL production. Klebsiella pneumoniae (ATCC 700603) was used as a positive control for ESBL production. Escherichia coli (ATCC 25922) was used as a negative control.
Identification of ESBL gene
Phenotypically confirmed ESBL-E were analyzed for the presence of genes encoding CTX-M-1, CTX-M-2, CTX-M-9, OXA-2, OXA-10, TEM, and SHV by multiplex polymerase chain reaction (PCR). Refer to Supplementary Table S1 for amplification primers and conditions. A loop full of pure culture on Mueller Hinton agar was suspended in 300 μL Milli-Q® water, heated for 10 min at 98°C, and subsequently centrifuged for 5 min at 4°C and 20,000 × g. The DNA extracts were stored at −20°C. For amplification of genes, 2 μL of template DNA, 12.5 μL of 2× Multiplex Master Mix (Qiagen), 2.5 μL of 10× reverse and forward primers of each gene type, and 7.5 μL of DNAse/RNAse-free water (Qiagen) were amplified using T100™ Thermal Cycler (Bio-Rad Laboratories). The amplified products were run on a 2% agarose gel tinged with 5 μL of 0.5 mg/mL ethidium bromide in a dark room. A constant voltage of 100 V for 30 min was used for separation. A 1× Tris-acetate-ethylenediaminetetraacetic acid (TAE) was used as a buffer. A 100-bp ladder molecular weight marker was used to measure the molecular weight of the amplified products. All PCR products with amplified genes for TEM, SHV, CTX-M-1, CTX-M-2, and CTX-M-9, as well as OXA-2, and OXA-10 were sent to Macrogen (Seoul, Korea) for nucleotide sequencing using Illumina’s next-generation sequencing technology. Additional internal primers (Supplementary Table S1) were used for sequencing CTX-M-1, CTX-M-9, SHV, and TEM genes. The contigs were examined using the ColorCode® analyzer (CodonCode Corporation), and the aligned nucleotide sequences were compared with sequences in the NCBI database (http://www.ncbi.nlm.nih.gov/BLAST). All CTX-M, TEM, and SHV β-lactamase sequences were compared to the wild-type E. coli CTX-M, TEM-1, and SHV-1 sequences (GenBank accession number AY536259.1, AF427133.1, and AF148850, respectively) at http://www.lahey.org/studies.
Data analysis
The study data were entered into Microsoft Excel for editing and analysis. Data were analyzed using STATA IC-12. The CrE isolates were compared in proportions and percentages. Similarly, ESBL-producing isolates were described using relative numbers and, where appropriate, proportions or percentages.
Results
Third-generation CrEs and ESBL phenotype
Housefly samples
Overall, third-generation CrE were cultured from 65% (n = 39/60) of housefly samples (Table 2). These included Proteus vulgaris (54%; n = 21/39), Klebsiella oxytoca (33%; n = 13/39), K. pneumoniae (8%; n = 3/39), and E. coli (5%; n = 2/39). Of these, 62% (n = 37/60) of CrEs were ESBL-positive and two were ESBL-negative. The 37 ESBL-positive Enterobacterales included 19 P. vulgaris, 13 K. oxytoca, 3 K. pneumoniae, and 2 E. coli. All 37 ESBL-positive isolates harbored an ESBL gene.
Homogenate Samples with Cephalosporins-Resistant and Extended-Spectrum Beta-Lactamase–Positive Enterobacterales
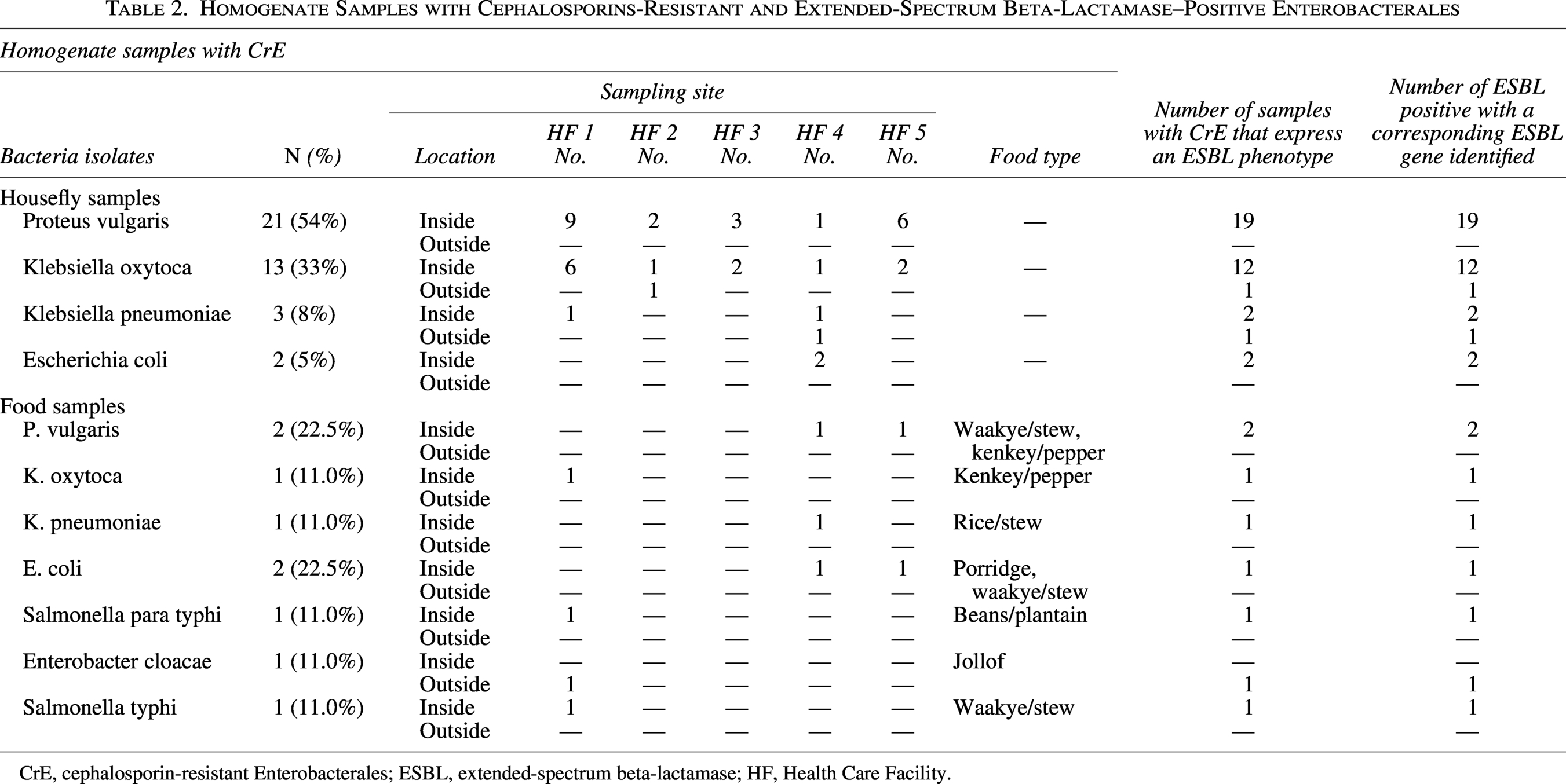
CrE, cephalosporin-resistant Enterobacterales; ESBL, extended-spectrum beta-lactamase; HF, Health Care Facility.
Food samples
Overall, 60 homogenate food samples were cultured, and nine (15%; n = 9/60) CrEs were recovered from nine homogenate samples (Table 2). This comprised Salmonella typhi (11%; n = 1/9), Salmonella para typhi (11%; n = 1/9), K. pneumoniae (11%; n = 1/9), K. oxytoca (11%; n = 1/9), Enterobacter cloacae (11%; n = 1/9), E. coli (22.5%; n = 2/9), and P. vulgaris (22.5%; n = 2/9). All nine CrEs were phenotypically confirmed for ESBL production and were identified to harbor the ESBL gene. This included two P. vulgaris, one K. pneumoniae, one K. oxytoca, one S. para typhi, one S. typhi, two E. coli, and one E. cloacae.
Molecular characterization of ESBL genes in CrE recovered from housefly
CTX-M was the most common ESBL (100%, n = 37/37). All ESBL producers harbored blaCTX-M alone or in combination with TEM, SHV, and/or OXA ESBL (Table 3). None of the isolates harbored only TEM, SHV, or OXA ESBLs. Overall, seven isolates harbored only a single gene of ESBL, which included four blaCTX-M-15, one blaCTX-M-28, and two blaCTX-M-128, while 20 isolates harbored a combination of the two ESBL genes (three blaCTX-M-15/TEM-3, one blaCTX-M-15/TEM-29, three blaCTX-M-15/SHV-2, one blaCTX-M-15/SHV-4, two blaCTX-M-15/SHV-31, three blaCTX-M-28/SHV-2, one blaCTX-M-101/SHV-2, one blaCTX-M-183/SHV-2, one blaCTX-M-183/TEM-3, two blaCTX-M-186/SHV-2, one blaCTX-M-186/SHV-5, and one blaCTX-M-186/SHV-24). Overall, 23 ESBL gene sequence types were identified. The highest diversity in ESBL sequence type was found for blaCTX-M (six different sequence types), blaSHV (five different sequence types), blaTEM (three different sequence types), and blaOXA (two different sequence types). The predominant ESBL sequence type was blaCTX-M-15 (n = 22), followed by blaSHV-2 (n = 13), blaCTX-M-186 (n = 8), blaTEM-3 (n = 7), blaCTX-M-28 (n = 4), blaSHV-24, and blaSHV-26 (n = 3).
Molecular Characterization of Extended-Spectrum Beta-Lactamase Genes in Cephalosporin-Resistant Enterobacterales Recovered from Housefly
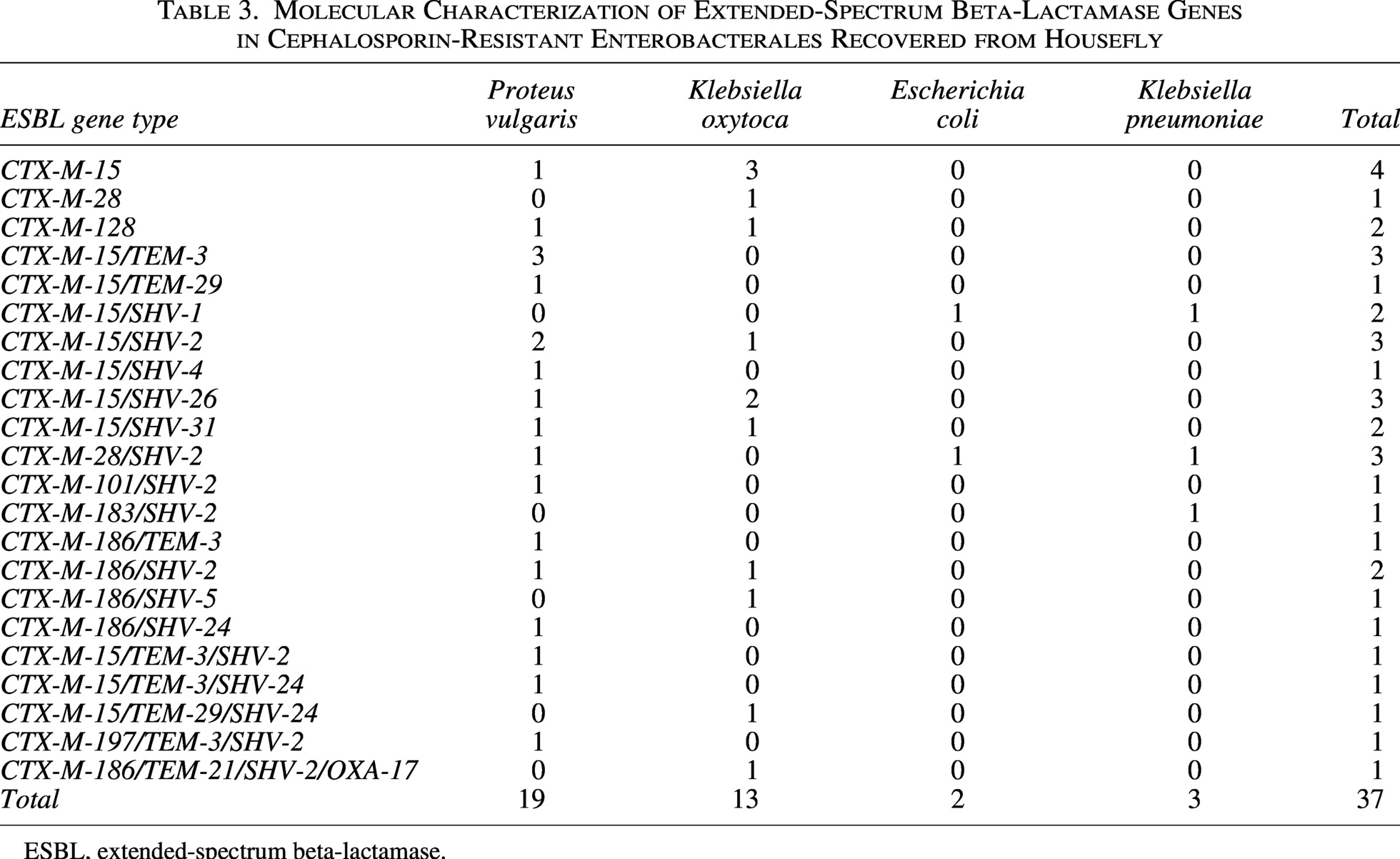
ESBL, extended-spectrum beta-lactamase.
Molecular characterization of ESBL gene in CrE recovered from food
All ESBL producers harbored blaCTX-M alone or in combination with TEM, SHV, and/or OXA ESBL (Table 4). None of the isolates harbored only TEM, SHV, or OXA ESBLs (Table 4). In total, five isolates harbored only one ESBL type (four blaCTX-M-15 and one blaCTX-M-18), three isolates harbored two ESBL sequence types (two blaCTX-M-15/TEM-21 and one blaCTX-M-101/SHV-2), and one isolate harbored three ESBL sequence types (blaCTX-M-194/SHV-18/OXA-17). None of the isolates obtained from food samples harbored the ESBL gene or another broad-spectrum gene. The most predominant ESBL gene type was blaCTX-M-15 (n = 6), followed by blaTEM-21 (n = 2), blaCTX-M-101 (n = 1), blaCTX-M-186 (n = 1), blaCTX-M-194 (n = 1), blaOXA-17 (n = 1), and blaSHV-18 (n = 1).
Molecular Characterization of Extended-Spectrum Beta-Lactamase Genes in Cephalosporin-Resistant Enterobacterales Recovered from Food
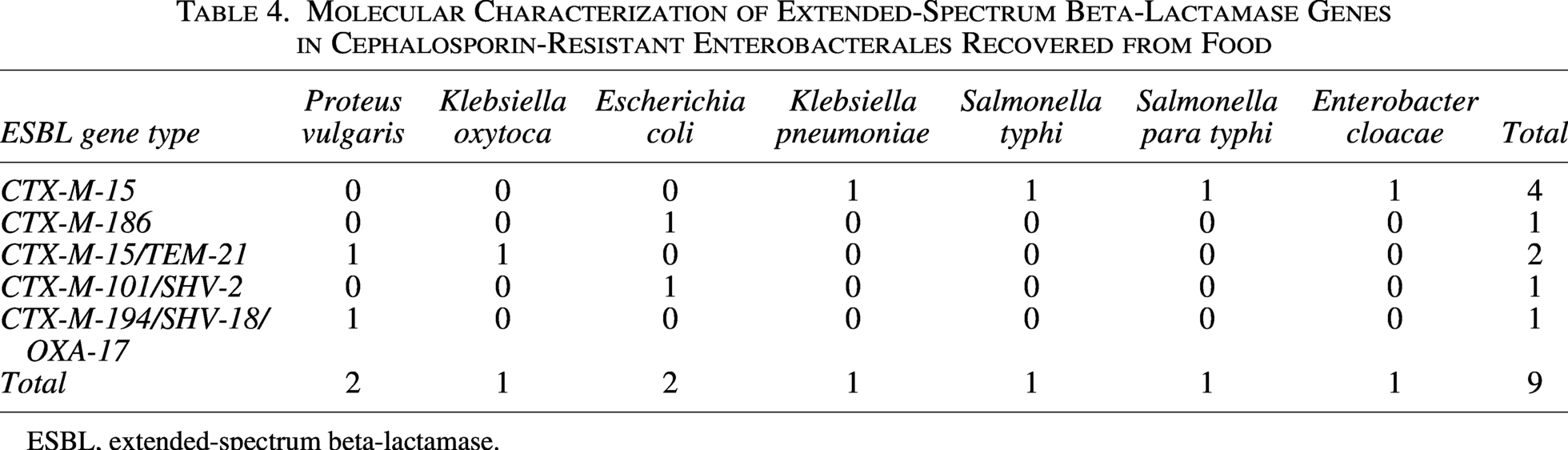
ESBL, extended-spectrum beta-lactamase.
Vendors with ESBL-E isolated from both housefly and food samples
Of the 60 food vending sites, nine had their site confirmed with ESBL phenotypic Enterobacterales from both houseflies’ samples and food samples (Table 5). Of the nine food vending sites, three had their sites harboring the same isolate from the housefly and their food sample, while the other six had different isolates from both the food sample and housefly. In addition, five food vending sites had ESBL-producing Enterobacterales isolated from both houseflies and food that harbored the same ESBL gene type. Among the nine food vending sites was blaCTX-M-15.
Food Vending Site with Extended-Spectrum Beta-Lactamase–Producing Enterobacterales from Both Housefly and Food Sample

CrE, cephalosporin-resistant Enterobacterales; ESBL, extended-spectrum beta-lactamase; HF, Health Care Facility.
We employed univariate analysis with odds ratios to assess associations between the predictor variables (sites 1, 2, 3, 4, and 5; and months 1 and 2 of sampling) and binary outcomes (presence or absence of ESBL producers in houseflies and food). None of the univariate analyses showed a significant association (data not shown).
Discussion
Houseflies (Musca domestica) can serve as reservoirs and vectors for human and animal pathogens because of their contact with animal manure, food, and humans (Rybaříková et al., 2010; Wang and Shelomi, 2017). This study sought to determine the carriage of ESBL-E at food vending sites in major hospitals in the Greater Accra Region. Four major findings of this study merit further attention. First, 62% of the sampled food vending sites had houseflies within their premises that carried ESBL-E. Second, 15% of the sampled food items were contaminated with ESBL-E. Third, ESBL-E was detected among diverse bacterial species in the order Enterobacterales. Last, despite the wide geographical dispersion of the sampling sites, CTX-M ESBL, mostly CTX-M-15, was the predominant ESBL gene type across all sampling points.
The finding that 62% of the vending sites had houseflies within their premises that carried ESBL-E is worrying. In Ghana, antibiotic resistance is high, and the prevalence of ESBL-producing pathogens is increasing (Donkor et al., 2019; Obeng-Nkrumah et al., 2013; Oduro-Mensah et al., 2016; Owusu et al., 2023). ESBL-E has been recovered from the environment, including from open water surfaces and fecal matter (Behrens et al., 2023; Poudel et al., 2019). The presence of ESBL-E in houseflies exacerbates this problem and could serve as a conduit for transferring AMR pathogens between hot spots and humans. A few studies with comparable methodologies have reported ESBL-E in houseflies among food vending sites. In Ethiopia, 67% of flies were captured from a food butchery site and a hospital carrying ESBL-E (Tufa et al., 2020). A study conducted in Zambia reported a lower prevalence (17.2%) of ESBL-producing E. coli in houseflies infesting fish in food markets (Songe et al., 2017). The observed difference in prevalence between the aforementioned studies and the current report may be associated with several factors, including variations in the sanitary conditions of the environment, geographical settings, and the frequent misuse of third-generation cephalosporin antibiotics in humans and the environment. Despite the paucity of data on the presence of ESBL-E in houseflies among food vending sites, some studies have also reported the carriage of other AMR pathogens such as methicillin-resistant Staphylococcus aureus and carbapenem-resistant Enterobacterales among food vending sites and animal farms (Seetswane et al., 2022; Solà-Ginés et al., 2015). Indeed, the current study supports the evidence that a housefly may be able to spread AMR pathogens across different settings (Madec et al., 2017; Tufa et al., 2020).
Isolation of ESBL-E from ready-to-eat food poses a public health concern. Earlier studies have reported that food can serve as a transmission vehicle for ESBL-producing pathogens, likely originating from reservoirs such as flies, food animals, and food handlers. Once an infection occurs, these pathogens have the potential to cause foodborne disease outbreaks (Madec et al., 2017; Sivakumar et al., 2021). Some studies with similar findings from Ghana, India, and Ethiopia have reported an estimated ESBL-E with food contamination prevalence of 4%, 15.2%, and 23.8% among food vending sites (Dela et al., 2023; Sivakumar et al., 2021; Tufa et al., 2020). The lower prevalence of ESBL-E recovered from food samples may be attributed to the rigorous exposure of these ready-to-eat food items to heat (boiling) during the preparation process, which can eliminate ESBL pathogens possibly contaminating food (Dela et al., 2023).
This study identified ESBL-E among diverse bacteria species of the order Enterobacterales, with P. vulgaris and E. coli being the predominant Enterobacterales recovered from houseflies and food samples, respectively. P. vulgaris is a gastrointestinal commensal known to be an opportunistic pathogen associated with nosocomial infections, including urinary tract infections, septicemia, and wound infections (Armbruster et al., 2018; Marcon et al., 2019). The recovery of Salmonella spp. from food samples is significant as salmonellosis is one of the most prevalent foodborne pathogens globally, with rising incidence rates in industrialized and developing countries. Salmonella spp. can infect humans and animals and is a major cause of diarrheal diseases with high morbidity and mortality rates worldwide (Songe et al., 2016). Contrary to our findings, some studies conducted in Ghana have reported E. cloacae as a common fecal-orally transmitted pathogen mostly recovered from ready-to-eat food (Dela et al., 2023; Yeboah-Man et al., 2010; Yeleliere et al., 2017).
The most predominant ESBL gene harbored by these ESBL-E isolates from both houseflies and food samples was blaCTX-M, mostly blaCTX-M-15 (59%, n = 22/37). The high prevalence of CTX-M-15 is concerning for several reasons. CTX-M-15 is mostly found on large plasmids with a high conjugation frequency per pathogen and can coexist with other resistance genes, such as blaAmpC, quinolone-resistance genes, or aminoglycoside-resistance genes, resulting in multidrug resistance (Chouchani et al., 2012; Sallem et al., 2015). Some studies have reported a varied prevalence of CTX-M-producing enzymes over the past decade, particularly in certain African countries, including Ghana (Bevan et al., 2017; Bonnet, 2004; Cantón and Coque, 2006). For example, the prevalence of CTX-M-15 in our current study was lower compared to the 80% CTX-M-15 prevalence reported for other hosts in Ghana by Obeng-Nkrumah et al. (2023) and 75% in a study conducted by Moremi et al. (2017) in Tanzania. In contrast, a study conducted in Nigeria by Fortini et al. (2015) reported a 32% prevalence rate of CTX-M-15. The high levels of CTX-M-15 reported in this study and elsewhere across Ghana highlight the urgent need for AMR control measures to limit the spread of such AMR pathogens.
The potential limitations of this study must be acknowledged. It is noteworthy that although this study focused on ESBL-E recovered from flies and meals at food vending sites, it does not establish the direct transmissibility of ESBL pathogens between flies and food samples. Although colonization with ESBL pathogens is common in flies, the study design does not clarify whether the external organs, such as the legs and mouth, or the gut of flies are more involved in carrying ESBL-producing pathogens, which could affect the transmission of the bacteria. Therefore, the findings of this study should be interpreted with caution. This study was limited to a few hospitals in the Greater Accra Region. As such, the data are less likely to be a true representation of the national sample. We were unable to collect data on food vendor behavior and hygiene practices. Such information would have provided better insights into the factors contributing to the occurrence and spread of ESBL pathogens within the sampling framework. We focused our investigations on the TEM, SHV, CTX-M, and OXA types because they are the most prevalent and diverse groups of ESBLs reported globally and in Ghana, constituting over 95% of ESBL genes identified worldwide. We specifically amplified CTX-M-1, CTX-M-2, and CTX-M-9 genotypes, as these moderate-sized parent gene blocks encompass all CTX-M nucleotide variants. Similarly, the OXA-2 and OXA-10 clusters cover all OXA ESBL gene nucleotide sequences.
Conclusions
The study identified a high prevalence of ESBL-E in houseflies and ready-to-eat foods at food vending sites in the Greater Accra Region. CTX-M, mostly CTX-M-15, was the predominant ESBL gene identified in this study. This finding emphasizes the need for strict adherence to proper sanitary conditions and proper personal hygiene.
Authors’ Contributions
N.O.-N., P.K., and A.-K.L. conceptualized the study, participated in its design, and acquisition of data. P.K., N.O.-N., A.-K.L., E.O., G.D.T.-A., S.S., D.B., D.G.J., and F.N.A.D. helped to draft the article and revise it critically for important intellectual content. P.K., N.O.-N., and A.-K.L. performed the data analysis. All authors have approved the final article.
Footnotes
Acknowledgments
We are grateful to God Almighty, and to the food vendors at the various health care facilities, for their voluntary participation in this study.
Funding Information
This study was self-funded.
Data Availability
The data are available from the corresponding author upon request.
Ethical Approval
Ethical approval was sought from the ethics, and protocol review committee of the SBAH before the study was carried out (Ethics Identification Number: SBAHS—MD./10484603/AA/5A/2016–2017). Permission was also sought from food vendors at the various sites.
Disclosure Statement
No competing financial interests exist.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.