
Abstract
Toxoplasma gondii is a highly complex protozoan parasite that poses significant health risks to humans and livestock. Traditional inactivated vaccines have simple preparation and high safety characteristics, but the protection is insufficient. This study aimed to find a new way to prepare an inactivated vaccine and find a suitable adjuvant to evaluate the immunoprotection. Inactivated vaccine (IVAC) was prepared by a novel low-temperature inactivation method, and different adjuvants were selected to evaluate the immune response. To assess immune protection, cytokines and other quantifiable factors associated with protection were examined, and then acute and chronic immune-protected experiments were carried out. IVAC has good integrity and biosafety. Immunizing mice with the adjuvant vaccine resulted in increased immunoglobulin G antibody and interferon-gamma levels, indicating the induction of a mixed Th1/Th2 immune response. Most notably, vaccination significantly improved the survival rate of mice. The IVAC can achieve a 10% protection rate, and the protection rate with adjuvant HA201 and HA203 can reach 50% and 70%, respectively. We found a new method to prepare IVAC and identified two adjuvants that could improve survival rates by 40–60%. These results provide valuable insights for future research on the Toxoplasma vaccine.
Introduction
Toxoplasma gondii is an obligatory intracellular protist that infects almost all warm-blooded animals, including humans. It is estimated that one-third of the world’s population is infected with the parasite (Djurković-Djaković et al., 2019; Molan et al., 2019). The life cycle of T. gondii is complex, with three forms of parasite: tachyzoites, bradyzoites (in cysts), and sporozoites (in sporulated oocysts). Tachyzoites, the rapidly replicating form of T. gondii, are essential for acute infection. The RH strain, a Type I strain, is highly virulent and widely used in laboratory studies due to its rapid proliferation and inability to form cysts in vivo. In contrast, the PRU strain, a Type II strain, is less virulent and forms tissue cysts, making it a suitable model for studying chronic toxoplasmosis (Liu et al., 2015; Su et al., 2012). Bradyzoites enclosed within tissue cysts are responsible for chronic infection. In immunocompromised hosts, bradyzoites escape from the cysts, causing severe complications such as encephalitis, pneumonia, and ocular disease (Elsheikha et al., 2021). Oocysts excreted in cat feces sporulate under suitable environmental conditions, becoming infectious and capable of infecting numerous intermediate hosts (Hill and Dubey, 2002).
The treatment of toxoplasmosis has traditionally relied on a combination of sulfadiazine and pyrimethamine, which is considered the standard of care. Nonetheless, the clinical application of these drugs is limited by significant adverse effects, restricting their long-term use. Moreover, their efficacy is primarily confined to the acute phase of T. gondii infection, with minimal effectiveness against chronic toxoplasmosis (Dunay et al., 2018). Currently, the only commercially available toxoplasmosis vaccine is used to prevent miscarriages in goats and sheep, highlighting an urgent need to develop a safe and effective vaccine for broader use (Hasan and Nishikawa, 2022).
Since the 1960s, various vaccine strategies, including attenuated and recombinant subunit vaccines, have been explored to prevent Toxoplasma infection (Chu and Quan, 2021). While attenuated vaccines have shown some utility, they carry an inherent risk of reversion to virulence, leading to potential clinical infections. In contrast, vaccines utilizing inactivated pathogens retain immunogenic properties and can elicit protective immune responses when subjected to appropriate physical or chemical treatments (Jongert et al., 2009).
In a previous study, Saavedra et al. employed both physical and chemical methods to inactivate infectious pathogens. Immunization with T. gondii soluble total antigen combined with adjuvants elicited a robust Th1-type immune response in mice, characterized by elevated levels of interleukins and interferon-gamma (IFN-γ). However, despite the activation of the immune response, the level of immune protection achieved was suboptimal (Saavedra et al., 2004). In another study, Stanley et al. administered a nanoparticle-coated tachyzoite crude extract to sheep via nasal inoculation. Postimmunization evaluations demonstrated a significant upregulation of IFN-γ and antibody production in the immunized sheep. Although the immunized group exhibited a slight reduction in fever symptoms following parasitic challenge, the vaccine failed to provide complete protection (Stanley et al., 2004). These findings highlight the need for continued research into vaccines that incorporate inactivated antigens with innovative adjuvants as a promising strategy for improving immune protection.
In this investigation, we employed a novel low-temperature method to inactivate the parasites. Subsequently, adjuvants were incorporated, resulting in the formulation of an antigen-adjuvant vaccine. The objectives of this study were to assess the characteristics of the inactivated vaccine (IVAC), explore the adjuvants’ capacity to modulate immune responses, and evaluate the efficacy of the inactivated parasites as vaccine candidates. Our findings suggest a promising avenue for the advancement of T. gondii vaccine development.
Materials and Methods
Mice and ethical statements
Female Institute of Cancer Research (ICR) mice, 6–8 weeks old and weighing 18–22 g, were obtained from the Comparative Medical Center of Yangzhou University. All animal experiments were performed in accordance with the guidelines of the Animal Care and Use Committee of the College of Veterinary Medicine, Yangzhou University.
Cell culture and parasite acquisition
Human foreskin fibroblast (HFF) cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM, USA) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 µg/mL streptomycin. The cells were incubated at 37°C in a humidified atmosphere containing 5% CO2.
The RH strain (Type I) and PRU strain (Type II) of T. gondii were maintained in our laboratory. The purified tachyzoites of the RH strain and cysts of the PRU strain were prepared according to a previously described method (Wu et al., 2009), and the cultures were subjected to centrifugation at 700 × g for 10 min. The supernatant was subsequently discarded, and the pellet containing the tachyzoites was resuspended in phosphate-buffered saline (PBS) at a pH of 7.4. The RH strain and PRU strain were collected separately in 15 mL centrifuge tubes for later use. The RH strain was used for vaccine preparation and virulent strain challenge, while the PRU strain was used for attenuated strain challenge.
Preparation of IVCA
The RH tachyzoites were inactivated by incubating at 37°C for 24 h and then maintained at 4°C for 20 days. The concentration was adjusted to 4 × 106/mL. The IVAC was stored at 4°C for future use. Adjuvants were purchased from Varnotech, China.
Evaluation of the inactivation effect of IVAC
In vitro, 6-well cell culture plates with confluent HFF cells were used, and the medium was replaced with 3 mL of DMEM containing 2% FBS. Thereafter, 25 µL of IVAC with 105 tachyzoites was added to each well of the experimental group, and 105 normal tachyzoites were added to the control group. Following a 5-day incubation period at 37°C in a CO2 incubator, the cultures were examined microscopically to detect the proliferation. Each experimental and control group was set up in duplicate.
In vivo, mice were randomly divided into two groups comprising 10 mice each. The experimental group was intraperitoneally injected with 25 µL of IVAC containing 105 tachyzoites. The control group was injected with 25 µL of PBS containing 105 normal tachyzoites. Postinjection, the mice were monitored daily for behavioral changes, disease progression, and mortality.
Immunization
Immunization process: Each mouse was immunized by intramuscular with 50 µL of IVAC containing 105 inactivated tachyzoites and an adjuvant (Fig. 3A). The ratio of antigen to adjuvant was 1:1. For the mixing process, equal amounts of adjuvant (HA) and antigen solution were drawn into two separate syringes. The two syringes were connected by a fine rubber tube, and the needle was alternately pushed back and forth until a thick emulsion was formed.
Groups: 20 mice were included in each group:
A: PBS
B: IVAC
C: IVAC + HA201 adjuvant
D: IVAC + HA203 adjuvant
Blood samples were collected after each immunization for analysis.
Preparation of toxoplasma lysate antigen
Toxoplasma lysate antigen (TLA) was prepared as previously described (Grzybowski et al., 2015). Briefly, tachyzoites obtained from the peritoneum of infected mice were washed with PBS and centrifuged at 800 × g for 10 min at 4°C. The isolated tachyzoites were disrupted by 10 cycles of freezing in liquid nitrogen and thawing at 37°C. The cell lysate was centrifuged at 10,000 × g for 20 min at 4°C. The total protein concentration was assessed by the bicinchoninic acid method.
Measurement of antibody responses
At 21- and 42-day postimmunization (dpi), ocular blood samples were collected from each group (comprising three mice per group), and antibody levels (immunoglobulin G [IgG], IgG1, IgG2a) were assessed via ELISA. Following a previously described study (Li et al., 2011). Briefly, 96-well plates were coated overnight at 4°C with the prepared TLA antigen. The plates were then washed with PBS containing 0.05% Tween 20 (PBST). Subsequently, the plates were blocked with 1% bovine serum albumin (BSA) at 37°C for 1 h. After washing with PBST, serum samples (diluted 1:200) were incubated in the wells at 37°C for 2 h. Following another wash, horseradish peroxidase-conjugated goat anti-mouse IgG (diluted 1:1000 in PBS-1% BSA) was added to the wells and incubated at 37°C for 1 h (Thermo Fisher Scientific, USA). After a final wash with PBST, the wells were incubated with tetramethylbenzidine substrate solution at 37°C for 10 min, and the reaction was terminated with 2M H2SO4. Absorbance was measured at 450 nm using a microplate reader; positive and negative samples were included in each experiment.
Determination of cytokine responses
Following a previously described study (Yang et al., 2020). The retrieved spleens were washed with RPMI-1640, gently ground on a 200-mesh nylon mesh to obtain a cell suspension, and centrifuged at 1500 × g for 10 min to remove cellular debris. All separated splenic cells were immersed in red blood cell lysate for 3 min to obtain a homogeneous splenic cell suspension. The suspension was then resuspended in RPMI-1640 culture medium supplemented with 10% FBS. The live cell count was determined using the trypan blue exclusion method to ensure >95% cell viability for subsequent analysis. The concentration of splenic lymphocytes was adjusted to 3 × 106 cells/mL and dispensed into individual wells. The splenocytes were then stimulated with TLA at a final concentration of 10 µg/mL to elicit an immune response. Following stimulation, the supernatants were harvested at specific time intervals: 96 h for IFN-γ quantification (mlbio, China). The collected supernatants were used to measure cytokine levels following the recommendations of the aforementioned kits; positive (ConA) and negative (PBS) controls were included in each experiment. Each sample was prepared in triplicate.
Protection against acute and chronic infection
At 42 dpi, 10 mice from each group were challenged with 1 × 102 RH tachyzoites to evaluate protection for acute infection. Another 10 mice from each group were challenged with 105 PRU tachyzoites to evaluate protection for chronic infection. The infection status and survival of these mice were monitored daily. Alongside daily survival monitoring, these mice were scheduled for euthanasia at 28 days postchallenge to assess cerebral cyst burden.
Based on the previously described method for detecting Toxoplasma cysts in the brain (Abdallah et al., 2022). Each brain hemisphere was thoroughly ground using a mortar and pestle. The resulting tissue was homogenized by mixing with 1 mL of 10% formalin. A 10 µL sample of the homogenized brain solution was transferred onto a glass slide and observed under a light microscope.
Statistical analysis
In this study, statistical analyses were performed using the GraphPad Prism software package, version 8.0.1 (GraphPad Software, San Diego, CA, USA). Differences among the groups, such as those of antibody responses and cytokine levels, were assessed using a one-way analysis of variance. A p-value of <0.05 was considered statistically significant. The levels of significance were indicated as follows: *p < 0.05, **p < 0.01, ***p < 0.001, and “ns” indicating a lack of significance.
Results
Vaccine characterization
The morphology of IVAC and normal tachyzoites was observed under a microscope. The results showed no differences between IVAC and normal tachyzoites (Fig. 1).
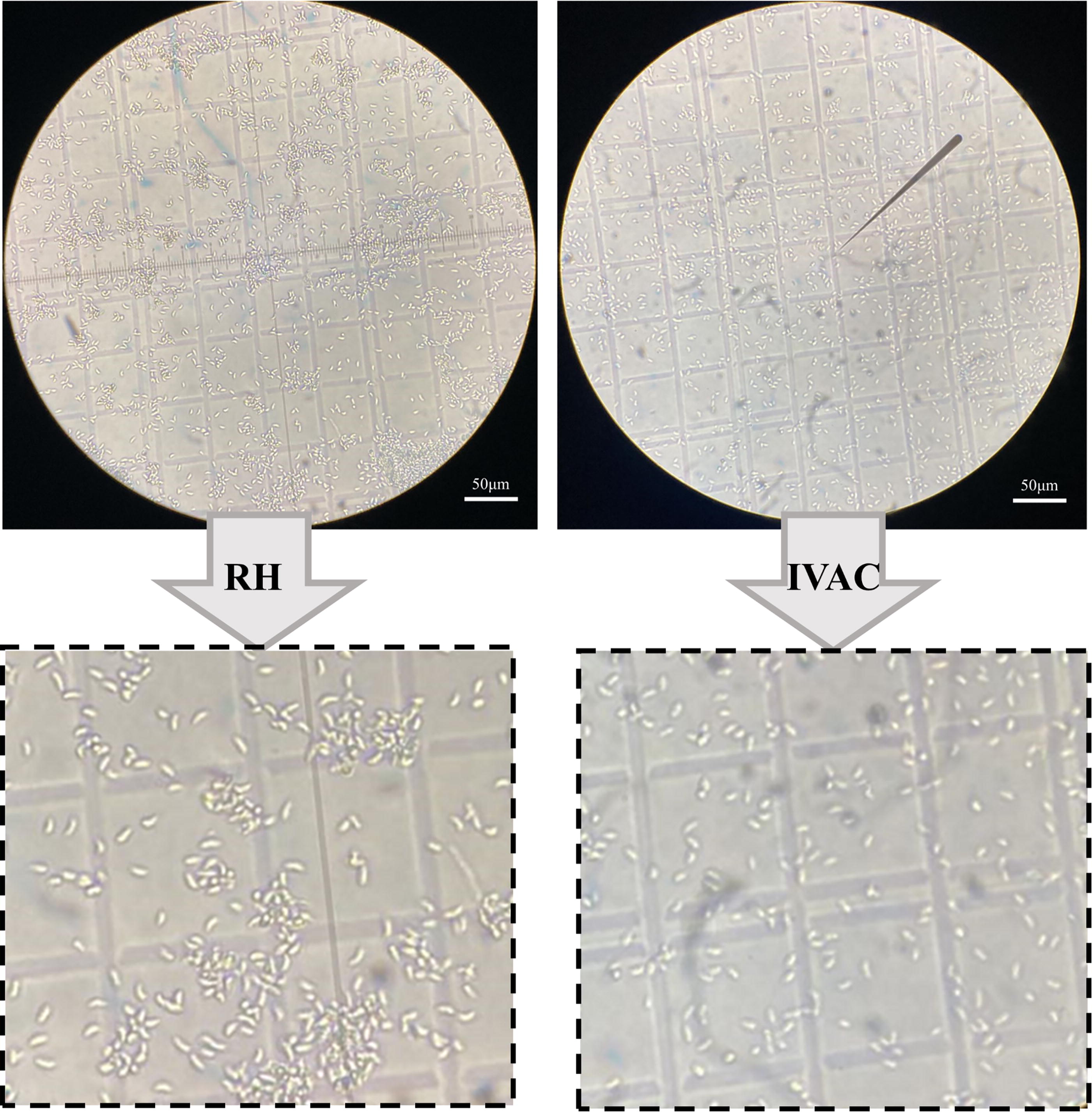
Characterization of IVAC. Light microscopy images of inactivated tachyzoites (left) and RH tachyzoites (right). IVAC, inactivated vaccine.
Evaluation of the inactivation effect of IVAC
From the results of cell culture experiments, we found that IVAC lost its proliferative ability in HFF cells compared to the control group, suggesting that the inactivation process was effective (Fig. 2B). Mice inoculated with IVAC maintained normal behavior, with no signs of illness or mortality observed. Conversely, in the control group, mice died on days 7 and 10 postinfection (Fig. 2A).
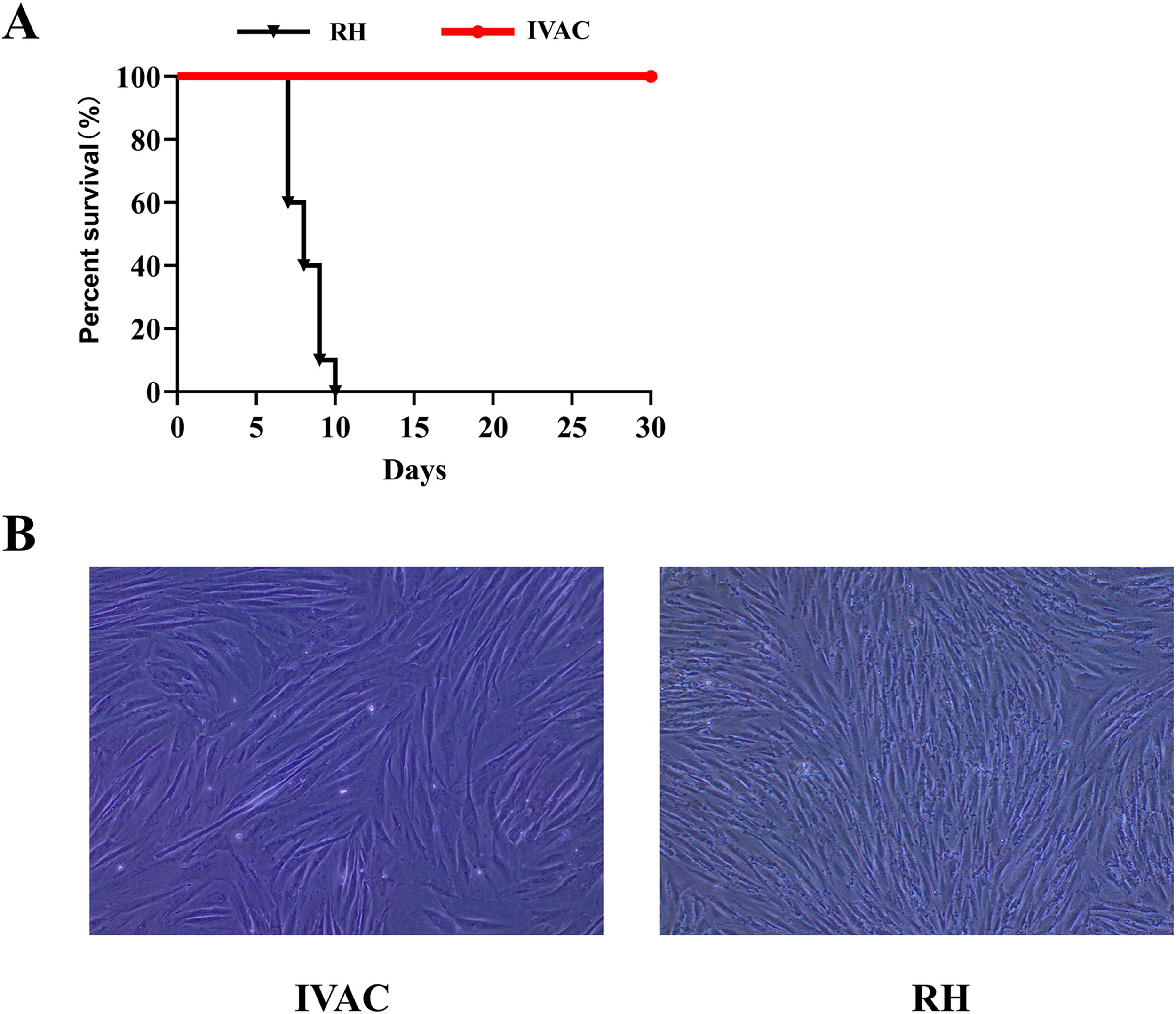
Safety evaluation of IVAC in mice.
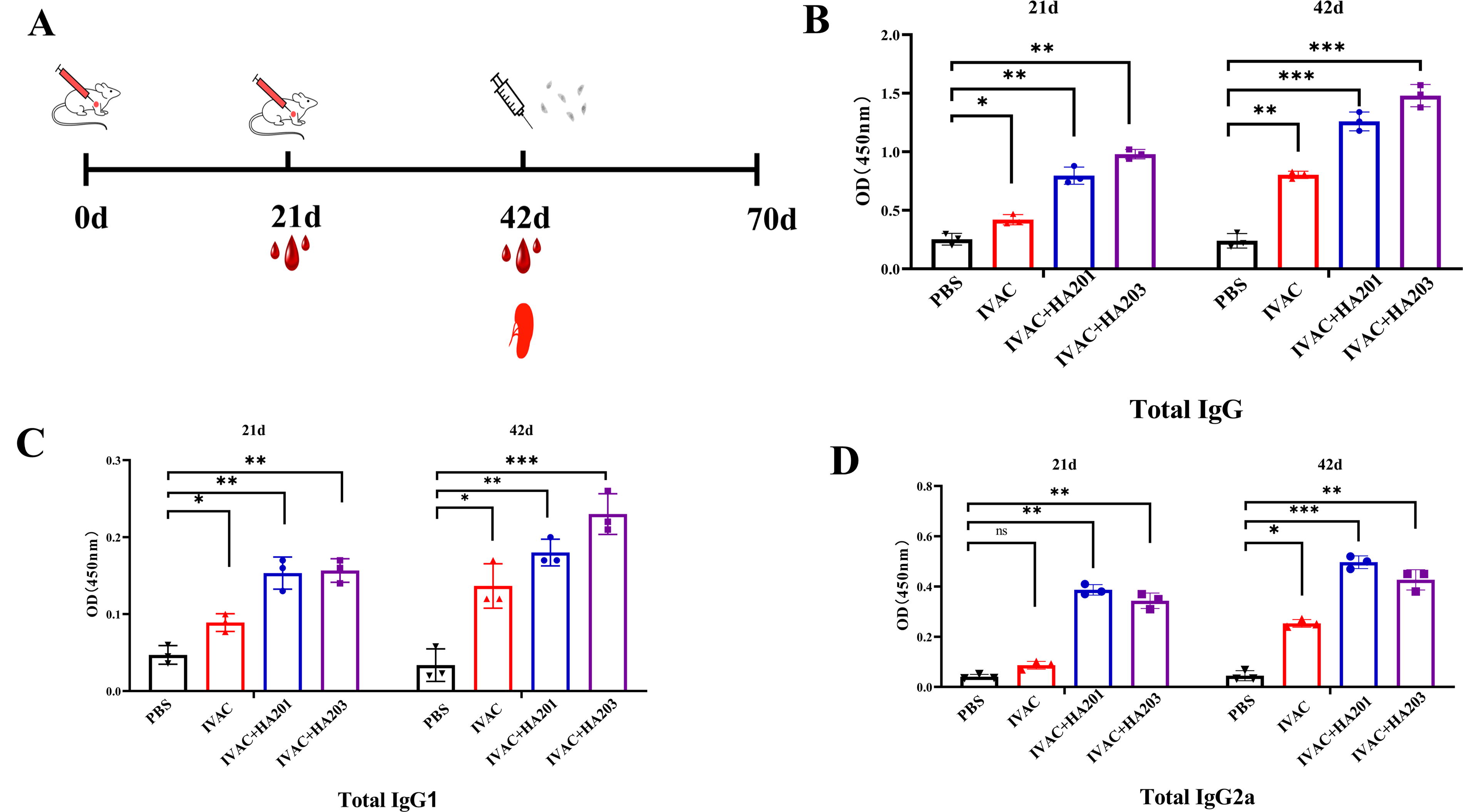
Immunization program and serum antibody levels in different groups.
Both in vivo and in vitro experimental results indicated that T. gondii was completely inactivated by the new inactivation method.
Humoral immune response induced by vaccination
To assess the level of specific humoral responses, the T. gondii IgG antibody levels and IgG subtypes were analyzed by ELISA at 21 and 42 dpi (Fig. 3A). The results indicated that all immunized mice exhibited elevated levels of T. gondii-specific IgG antibodies in their serum compared to non-immunized mice at 21 dpi, with IgG levels remaining high at 42 dpi (Fig. 3B). Notably, the IgG levels in mice immunized with IVAC + HA201/HA203 were significantly higher than those immunized with IVAC alone, suggesting that IVAC + HA201 and IVAC + HA203 elicited robust humoral immune responses.
At 21 dpi, the levels of IgG1 and IgG2a in mice immunized with IVAC + HA201 and IVAC + HA203 were significantly higher than those in the control group and the IVAC group. No significant differences were observed in the levels of IgG1 and IgG2a between the two adjuvant groups. At 42 dpi, IgG1 and IgG2a levels remained significantly elevated in vaccinated mice compared to controls (Fig. 3C, D).
Cellular immune response induced by vaccination
Splenocytes were isolated at 42 dpi to evaluate the cellular immune response elicited in mice by measuring cytokine production in the supernatant of splenocytes stimulated with TLA. A slight increase in IFN-γ levels was observed in splenocytes from IVAC-immunized mice, while a significant elevation in IFN-γ levels was detected in the IVAC + HA201 and IVAC + HA203 groups (Fig. 4).
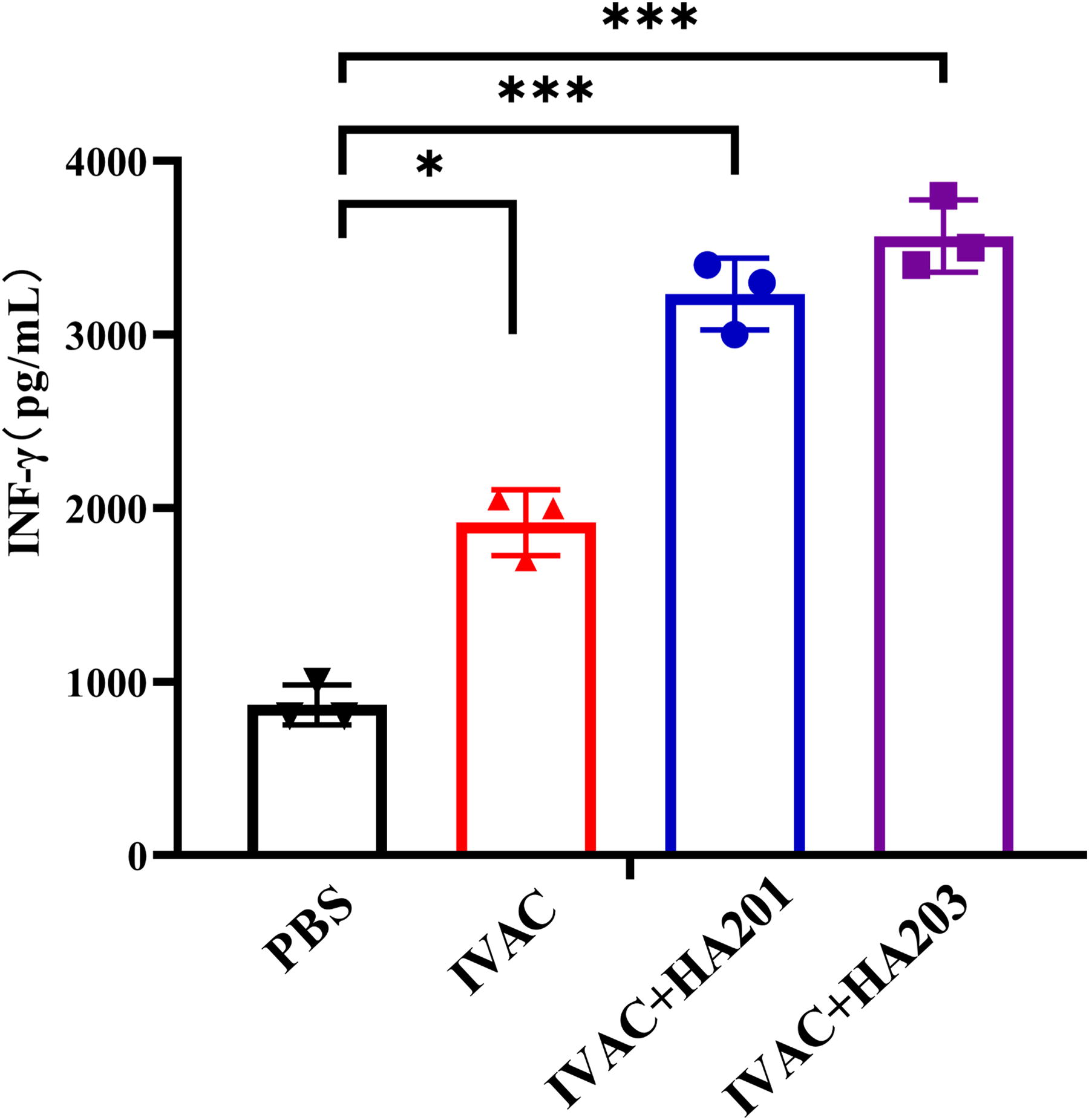
IFN-γ response levels in splenocytes of different groups. At 42 dpi, splenocytes from three mice in each group were randomly collected and stimulated with 10 µg/mL TLA in vitro. The cell culture supernatants were then collected for cytokine detection (*p < 0.05, **p < 0.01, ***p < 0.001). IFN-γ, interferon-gamma.
Protection against acute and chronic infection
At 42 dpi, 10 mice from four distinct groups were intraperitoneally injected with 1 × 102 RH tachyzoites. The survival curves for each group are depicted (Fig. 5). Mice in the IVAC group exhibited a survival rate of 10%. Notably, the survival rates for the mice in the IVAC + HA201 and IVAC + HA203 groups were 50% and 70%, respectively. Mice in the control group died within 10 days postinfection. These results suggest that the IVAC confers a degree of protection against acute infection, and adding adjuvants to the vaccine significantly improves the survival rate of mice.
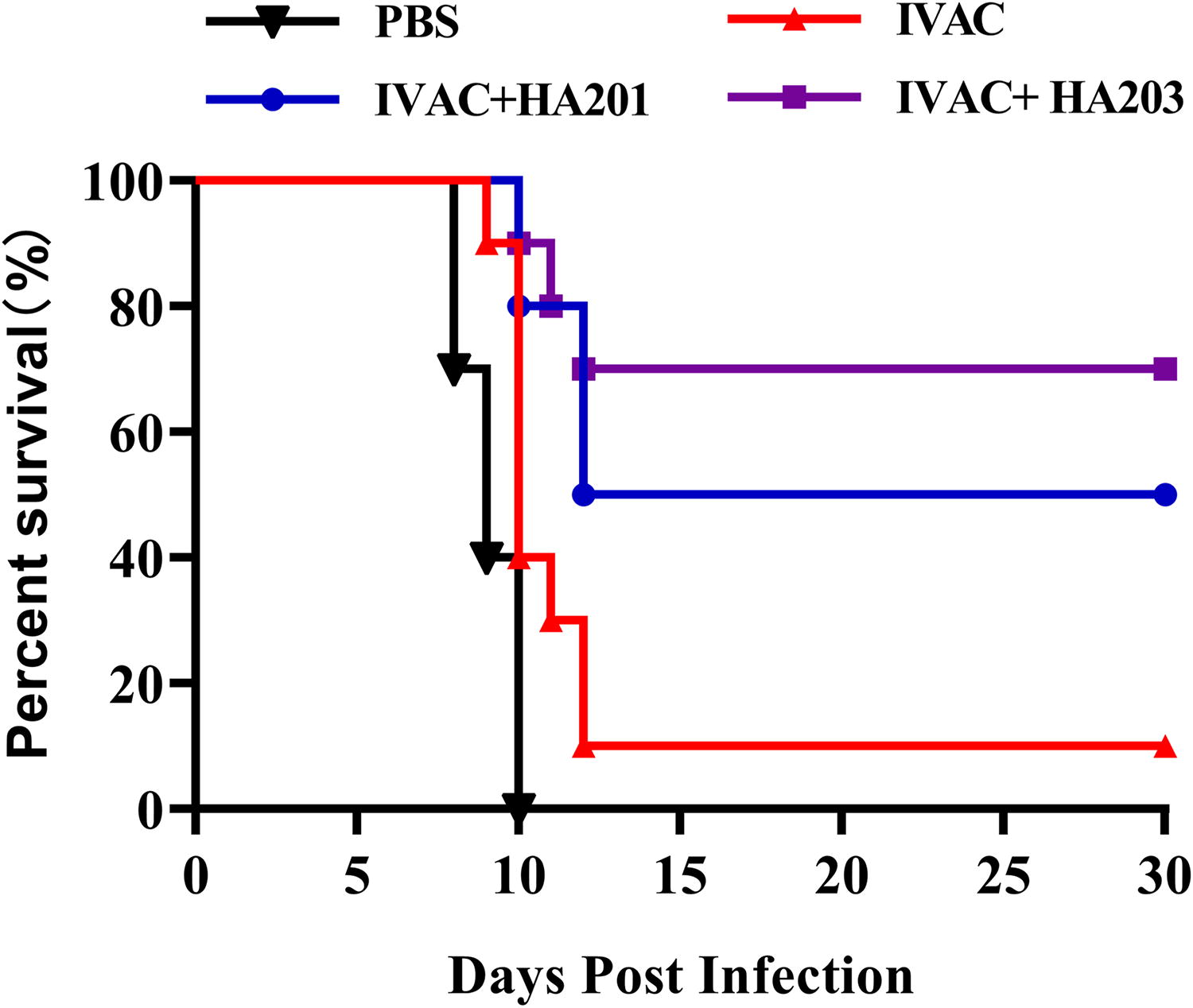
Protection against acute infection. Survival curves of different groups that were challenged with 1 × 102 tachyzoites of the RH strain at 42 dpi (n = 10 mice per group).
The survival rates and brain cyst burdens were assessed to evaluate the efficacy against chronic infection. All immunized groups exhibited a 100% survival rate, whereas only 80% of the nonvaccinated mice survived (Fig. 6A). At 28 days postinfection, the number of brain cysts in vaccinated mice was significantly lower compared to the nonvaccinated mice (Fig. 6B). The cyst counts in the IVAC + HA201 and IVAC + HA203 groups were reduced by 60% and 50% compared to the control group, respectively.
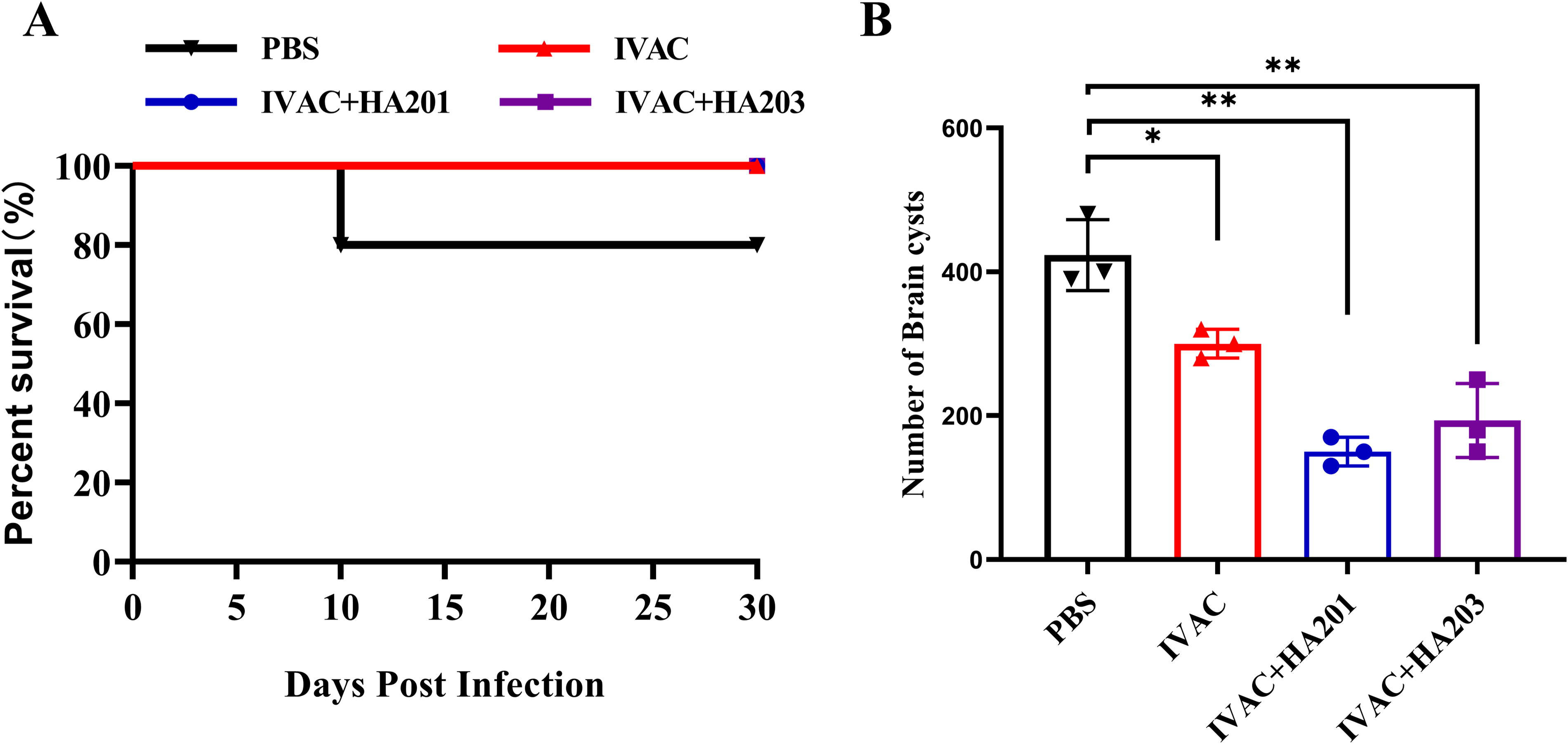
Protection against chronic infection.
Discussion
T. gondii is an obligate intracellular protozoan parasite, and toxoplasmosis represents a globally distributed zoonotic disease of significant concern (Cenci-Goga et al., 2011; Montoya and Liesenfeld, 2004). To date, a single commercial vaccine has been authorized for use in ovine populations to mitigate the incidence of Toxoplasma-induced abortion (Stanley et al., 2004). However, this vaccine faces limitations, including a short shelf life and restricted protective efficacy. While attenuated live vaccines generated through targeted gene deletion via genetic engineering hold promise, they carry inherent risks of virulence reversion and mutation. Consequently, the IVAC is considered a safer alternative; however, its protective efficacy remains a significant challenge that needs to be addressed (Jongert et al., 2009).
Studies have elucidated that parasites secrete and excrete large amounts of excretory–secretory (ES) antigens into the host organism, which directly interact with the host immune system, eliciting both humoral and cellular immune responses (Ezz Eldin et al., 2015; Saraav et al., 2019). During the inactivation process, a 37°C incubation for 24 h was included; the tachyzoites released substantial quantities of ES antigens, which function synergistically with the intact inactivated tachyzoites to exert their effects. Capitalizing on the unique characteristics of T. gondii, we developed a method for preparing IVAC.
The virulence and proliferation of IVAC were assessed both in vitro and in vivo. No proliferation of IVAC was observed in HFF cells, and all mice survived after intraperitoneal injection with 1 × 105 IVAC. These results suggest that the new inactivation method can completely inactivate tachyzoites, and the biosafety of IVAC is adequate for use as an IVAC.
Specific IgG antibodies also play a crucial role in controlling T. gondii infection, as they can facilitate the destruction of the parasite through macrophage activation and complement-mediated lysis (Liyanage et al., 2021; Zorgi et al., 2011). In this study, the groups with adjuvants exhibited significantly higher IgG levels compared to the control group. Analysis of IgG1 and IgG2a levels indicated that mixed Th1/Th2 immune responses were induced in each immune group, while the levels of response were significantly higher in the IVAC + HA203 group compared to other groups.
It is widely recognized that Th1 immune responses are crucial for controlling T. gondii infection (Denkers and Gazzinelli, 1998; Sasai et al., 2018). Consequently, an adjuvant with the ability to elicit a Th1 response was important for a vaccine against T. gondii. The HA series of adjuvants meets this requirement, as it activates and induces Th1 cell-mediated immunity. IFN-γ is associated with Th1 immune responses and plays a pivotal role in controlling T. gondii infection. It mediates the host’s immune defense against the parasite through multiple mechanisms.
To evaluate the protective efficacy of our candidate vaccines, we established an immunization model and monitored the survival rates to assess the immune protection. Our findings indicated that mice immunized with IVAC alone exhibited a survival rate of 10%, whereas those receiving the adjuvant-enhanced vaccine demonstrated a substantial increase in protection, with a survival rate of 70%, indicating effective immune protection. A study demonstrated that TLA combined with CpG-ODN adjuvant could increase the survival rate of mice by 20% (Saavedra et al., 2004). In this study, HA201 could increase the survival rate of mice by 40%, and HA203 could increase it by 60%. These results indicated that HA201 and HA203 had a good adjuvant effect, especially in T. gondii. During chronic infection, all immunized groups successfully survived. Although the formation of tissue cysts was not completely inhibited, mice vaccinated with IVAC + HA201 and IVAC + HA203 successfully survived, and a reduced number of cysts was observed compared to the control group. The preparation of TLA is straightforward, but its immune protection is suboptimal (El-Malky et al., 2014; Saavedra et al., 2004). In contrast, IVAC requires a longer preparation time and is not immediately usable; however, the addition of HA201/HA203 to IVAC has proven to be both safe and effective.
Our research findings demonstrated that the addition of adjuvants to the vaccine induced a robust humoral and cellular immune response in immunized mice, significantly improving their survival rate. This finding is important for obtaining a safe and effective toxoplasmosis vaccine to control toxoplasmosis.
Conclusion
Our study found a novel low-temperature inactivated T. gondii method, which was safe and effective. Two good adjuvants (HA201, HA203) were found; HA201 and HA203 could increase the survival rate of mice by 50% and 70%, respectively. These results lay the foundation for the development of T. gondii vaccines.
Footnotes
Acknowledgment
The authors would like to thank Yi-min Fan for technical assistance.
Authors’ Contributions
S.Y.H.: Funding acquisition, conceptualization, methodology, formal analysis, investigation, and writing—review and editing. S.Y.B.: Conceptualization, methodology, formal analysis, writing—original draft, validation, and project administration. G.Q.Z.: Conceptualization, methodology, writing—original draft, and project administration. Y.Y.L.: Conceptualization, investigation, and writing—original draft. M.P.: Conceptualization and writing—review and editing. L.F.: Methodology and Writing—review and editing.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This work was supported by the Strategic Priority Research Program of the National Center of Technology Innovation for Pigs (NCTIP-XD/B19), Chongqing Municipal Financial Fund Projects (24535C), and the Priority Academic Program Development of Jiangsu Higher Education Institutions (Veterinary Medicine).