
Abstract
Cyclospora spp. is a globally distributed foodborne intestinal parasite with potential zoonotic risk. The Tibetan antelope (Pantholops hodgsonii), an iconic wildlife species in China, is classified as a national first-class protected animal. However, data on Cyclospora spp. infections in Tibetan antelope remain limited. This study employed polymerase chain reaction–restriction fragment length polymorphism analysis targeting the small subunit ribosomal RNA gene of Cyclospora spp. on 503 fecal samples collected from Tibetan antelope in the Tibet Autonomous Region, Xinjiang Uygur Autonomous Region, and Qinghai Province. The overall infection rate was 1.2%, with the highest rates observed in Qumarlêb County (4.4%), followed by Qiemo County (3.3%) and Shuanghu County (0.5%). No infections were detected in Shenza, Nima, and Ruoqiang counties. Infection rates were highest in summer (3.3%), followed by autumn (1.2%), with no infections detected in spring. Across sampling years, the highest infection rate was recorded in 2024 (3.3%), followed by 2020 (0.3%), with no infections detected in 2023. Statistical analysis identified sampling region and year as significant factors influencing infection rates. Additionally, six isolates were identified as Cyclospora spp. This study provides the first evidence of Cyclospora spp. infection in Tibetan antelope, expanding its known host range and geographic distribution.
Introduction
Cyclospora spp., an intestinal parasitic protozoan, belongs to the subkingdom Protozoa, subphylum Apicomplexa, subclass Coccidiasina, order Eucoccidiorida, family Eimeriidae, and genus Cyclospora (Shields and Olson, 2003). To date, 22 species of Cyclospora spp. have been identified in humans and animals based on morphological and molecular data, with Cyclospora cayetanensis being the only species known to infect humans (Giangaspero and Gasser, 2019; Li et al., 2020; McAllister et al., 2018). Humans are believed to become infected by consuming food or water contaminated with oocysts (Dubey et al., 2022). Contaminated fruits, vegetables, water, and soil are the primary environmental sources of human infection with C. cayetanensis (Onstad et al., 2019). Symptoms of infection include severe watery diarrhea, abdominal cramps, mild fever, nausea, anorexia, weight loss, and fatigue (Connor et al., 1999; Ortega et al., 1997). Several Cyclospora spp. species have been identified in animals, including C. cayetanensis, C. cercopithi, C. colobi, C. duszynskii, C. macacae, C. papionis, and C. yatesi (Almeria et al., 2019; Eberhard et al., 1999; McAllister et al., 2018; Totton et al., 2021). Notably, contact with animals has been identified as a risk factor for human Cyclospora spp. infection (Almeria et al., 2019; Chacín-Bonilla, 2010).
The prevalence of human C. cayetanensis infection in China was reported to be 0.8% (214/25,825) (Chen et al., 2024). The prevalence rate of C. cayetanensis in China showed regional differences among provinces. For example, the prevalence rate was 0.7% (81/11,554) in Henan (Zhou et al., 2011), whereas it was 1.7% (5/291) in Shanghai (Jiang et al., 2018). Among animals, the prevalence of Cyclospora spp. in China was 1.3% (11/862) in fur-bearing animals (Xue et al., 2025) and 2.7% (22/812) in snakes (Zhang et al., 2025). The prevalence of Cyclospora spp. in cattle was 2.1% (16/761) and 2.5% (13/524) in Shanxi and Yunnan Provinces, respectively (Wu et al., 2024; Yang et al., 2023). The prevalence of C. cayetanensis in Mongolian gazelles was 30.00% (36/120) (Gao et al., 2024).
The Tibetan antelope (Pantholops hodgsonii) belongs to the genus Pantholops, family Bovidae, and order Cetartiodactyla. Tibetan antelope is among the rarest and most endangered wild animals. An estimated 100,000–150,000 Tibetan antelopes remain in India and China (IUCN SSC Antelope Specialist Group, 2016). Tibetan antelopes can carry various pathogens, including Mycoplasma capricolum subspecies capripneumoniae (Yu et al., 2013) and Escherichia coli (Bai et al., 2016). Currently, there is a lack of global data on the incidence and prevalence of Cyclospora spp. in Tibetan antelopes. This study investigated the prevalence of Cyclospora spp. in Tibetan antelopes, identified its distribution, and assessed the risk of zoonotic transmission. These findings provide crucial epidemiological data to aid in the prevention and control of Cyclospora spp. in Tibetan antelopes.
Methods
Sample collection
From September 2020 and between July 2023 and May 2024, 503 fecal samples were collected from Tibetan antelopes in the Tibet Autonomous Region (Shenza County, n = 82; Nima County, n = 64; Shuanghu County, n = 204), Xinjiang Uygur Autonomous Region (Ruoqiang County, n = 31; Qiemo County, n = 30), and Qinghai Province (Qumarlêb County, n = 92), China. Fresh samples were randomly collected in the field with disposable sterile gloves and then transported in ice boxes to the laboratory for storage at −80°C. This study was approved by Institutional Animal Care and Use Committee of Qingdao Agricultural University (Approval No. QAU-AEW-20200701001).
DNA extraction and polymerase chain reaction amplification
DNA was extracted from each stool sample using the Stool DNA Kit (Omega Bio-Tek Inc., Norcross, GA, USA) according to the manufacturer’s protocol and stored at −20°C. DNA preparations were analyzed for the presence of Cyclospora spp. using nested polymerase chain reaction (PCR) to amplify a ∼500 bp fragment of the small subunit ribosomal RNA (SSU rRNA) gene, as previously described (Li et al., 2007). The first-round PCR protocol included an initial denaturation at 94°C for 7 min, followed by 35 cycles of 95°C denaturation for 45 s, 55°C annealing for 45 s, and 72°C extension for 1 min 30 s. The second-round PCR followed the same thermal cycling conditions as the first, except the extension time was reduced to 1 min. All reactions employed the EXTaq enzyme (Takara, Shiga, Japan). The secondary PCR products were analyzed by 1.5% agarose gel electrophoresis and visualized using GoldView™ stain (Solarbio, Beijing, China).
Restriction fragment length polymorphism analysis
Restriction fragment length polymorphism (RFLP) analysis was carried out following the product manual to differentiate Cyclospora spp. from Eimeria spp. (Chu et al., 2004). A 20 µL reaction mixture was prepared with 2.5 µL of 10× buffer, 1 unit of Kpn2I restriction enzyme (NEB, Ipswich, MA, USA), and 12.5 µL of PCR product. The mixture was incubated at 55°C for 12 h for digestion. Afterward, samples were electrophoresed on a 1.5% agarose gel and visualized under ultraviolet light.
Sequencing and phylogenetic analysis
The positive secondary PCR products were sent to Tongyong Biotech Company (Anhui, China) for bidirectional sequencing. The obtained sequences were edited and analyzed for similarity by comparing them with reference strains on the BLAST server and in the GenBank database. A phylogenetic tree was constructed using MEGA 6.0 (http://www.megasoftware.net/) through the Neighbor–Joining method, applying the Kimura 2-parameter model, and supported by a bootstrap analysis with 1000 iterations. Finally, the phylogenetic tree was beautified through the ChiPlot website (Xie et al., 2023).
Statistical analysis
The chi-square test was performed in SPSS 22.0 (IBM Corp., Armonk, NY, USA) to assess the association between the molecular prevalence of Cyclospora spp., and various risk factors. Odds ratios and 95% confidence intervals were also calculated, with a p value of <0.05 considered statistically significant.
Results
Detection of Cyclospora spp. by nested PCR
Nested PCR detected a specific band of approximately 500 bp in 192 fecal samples from Tibetan antelopes. The PCR amplicon of Cyclospora spp. SSU rRNA gene can be digested into 130 bp and 370 bp fragments with the restriction endonuclease Kpn2I, but the PCR product amplified from Eimeria spp. cannot be digested (Fig. 1). Digestion yielded fragments of the expected sizes (130 bp and 370 bp) in 6 samples (Table 1). The detection rate of Cyclospora spp. in Tibetan antelopes was 1.2% (6/503).
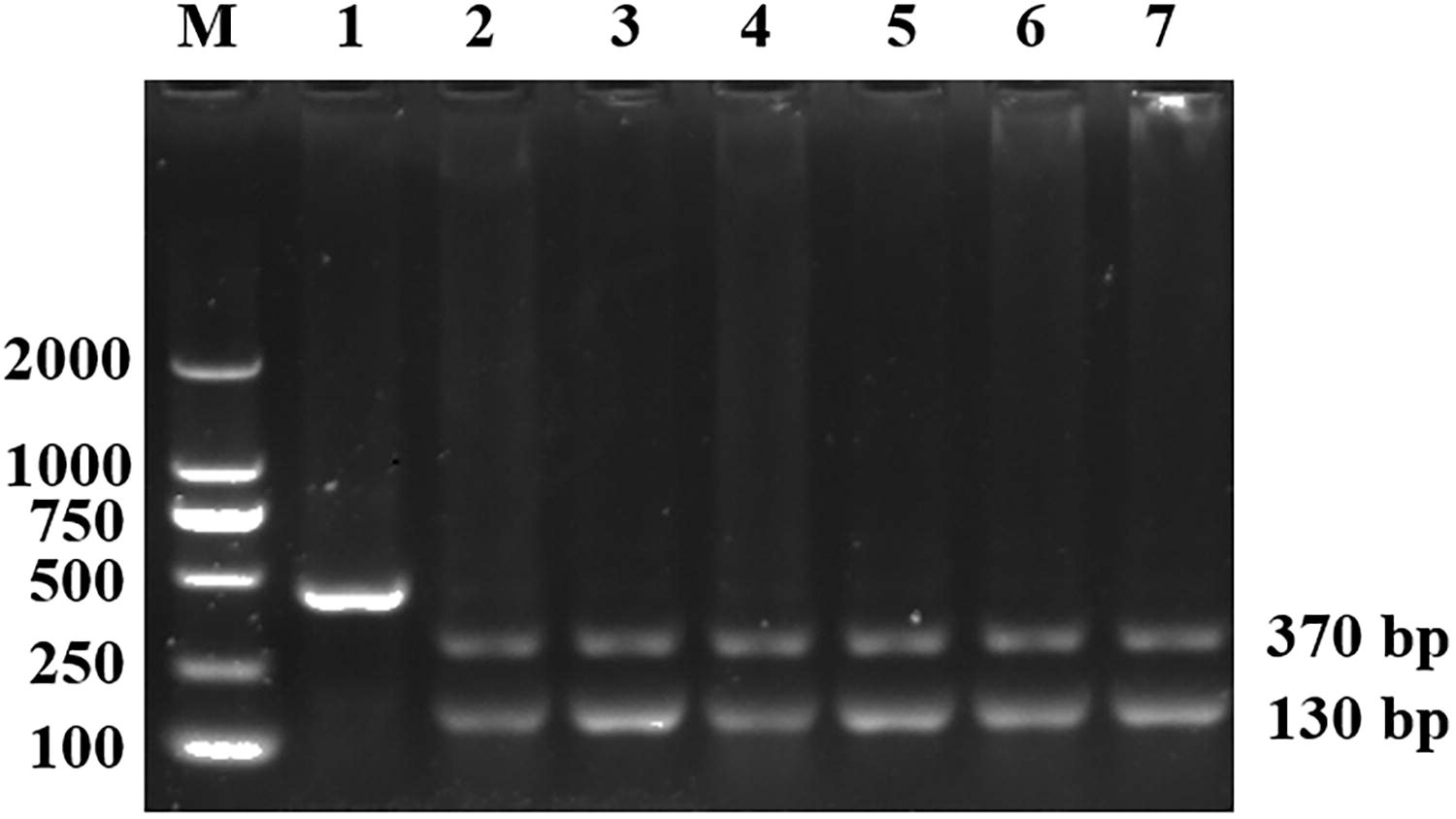
Partial SSU rRNA gene amplicons were digested using the restriction endonuclease Kpn2I. Lane M: DL 2000 DNA Marker; Lanes 1–7: Kpn2I digested products from nested PCR. SSU rRNA, small subunit ribosomal RNA.
Factors Associated with the Prevalence of Cyclospora spp. in Tibetan Antelope in China
CI, confidence interval; OR, odds ratio.
Risk factors of Cyclospora spp. infection
Qumarlêb County showed the highest prevalence of Cyclospora spp. infection at 4.4% (4/92), followed by Qiemo County at 3.3% (1/30) and Shuanghu County at 0.5% (1/204). No Cyclospora spp. infections were detected in Shenza, Nima, or Ruoqiang counties. Significant differences were observed in the Cyclospora spp. infection rates among Tibetan antelopes across different regions (p = 0.0358). The prevalence of Cyclospora spp. was 3.3% (1/30) in summer and 1.2% (5/423) in autumn, with no Cyclospora spp. detected in spring (0/50) (p = 0.4216). The prevalence of Cyclospora spp. was higher in 2024 (3.3%, 5/153) compared with 2020 (0.3%, 1/300), with no Cyclospora spp. detected in 2023 (0/50). Significant differences were found in the prevalence of Cyclospora spp. across different years (p = 0.0177) (Table 1).
Sequence alignment and phylogenetic analysis
In this study, the PCR products successfully digested by Kpn2I were identified by Sanger sequencing analysis, yielding six different sequences. The sequences were uploaded to the GenBank database, with accession numbers PV053000–PV053005. Two sequences (PV053000 and PV053001) share 99.4–99.8% homology with Cyclospora spp. isolated from Holstein cattle in Yunnan, China (OP268225), showing 1–3 base differences. Three sequences (PV053002–PV053004) share 97.8–99.4% homology with Cyclospora spp. isolated from Holstein cattle in Yunnan, China (OP268233), with 3–10 base differences. The remaining sequence (PV053005) shares 99.2% homology with Cyclospora spp. isolated from cattle in Shanxi, China (PP930929), exhibiting four base differences. All six samples isolated from Tibetan antelopes were classified as Cyclospora spp. (Fig. 2).
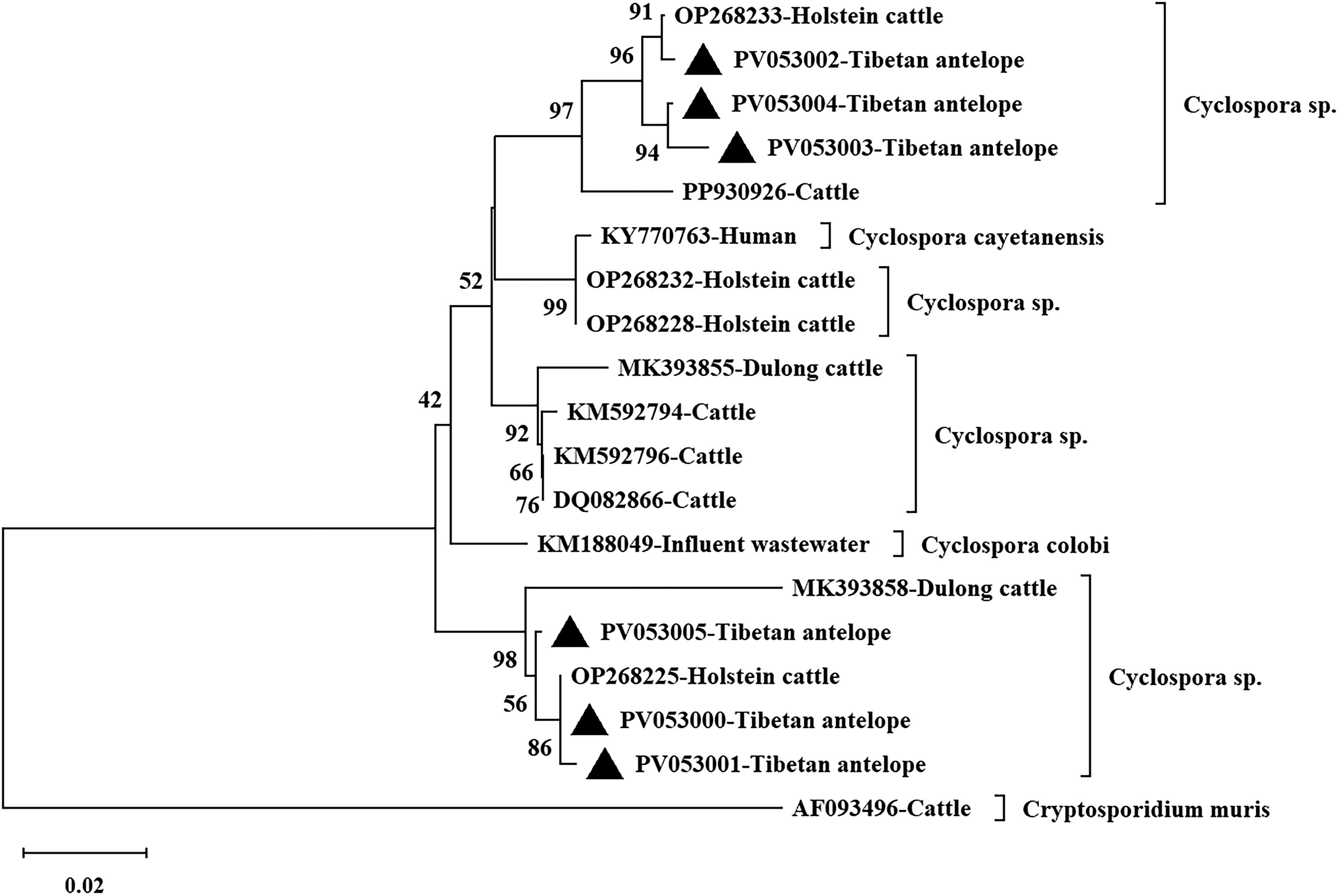
The phylogenetic relationship of Cyclospora spp. found in Tibetan antelope in China was analyzed using the Neighbor–Joining (NJ) method in MEGA 6.0, with sequences from this study marked by solid triangles in the resulting phylogenetic tree.
Discussion
Detection methods for Cyclospora spp. primarily include morphological examination and molecular biology techniques. However, due to the morphological similarity between Cyclospora spp. and Eimeria spp., microscopy cannot reliably distinguish them. In contrast, molecular detection methods offer higher specificity and sensitivity for diagnosing Cyclospora spp. infection. The SSU rRNA gene is highly conserved, making it an ideal genetic marker to differentiate Cyclospora spp. from other apicomplexan protozoa (Relman et al., 1996; Sulaiman et al., 2014). In 1996, scientists developed a nested PCR method targeting the SSU rRNA gene fragments of Cyclospora spp. (Relman et al., 1996). Subsequently, researchers developed an RFLP analysis method for PCR products, based on nucleotide differences in the amplification region, to distinguish Cyclospora spp. from Eimeria spp. (Jinneman et al., 1998). Currently, restriction enzymes used in RFLP analysis include MnlI, KpnI, and Kpn2I (synonym BspEI) (Li et al., 2007; Ortega and Sanchez, 2010; Yang et al., 2023). In this study, we selected Kpn2I to digest SSU rDNA PCR products amplified from Tibetan antelope fecal samples.
There is no available epidemiological data on Cyclospora spp. infection in Tibetan antelopes. In this study, Cyclospora spp. was detected in Tibetan antelopes with an infection rate of 1.2% (6/503). This rate is lower compared to the infection rates of Blastocystis (4.8%, 30/627) (Geng et al., 2021) and Cryptosporidium (3.0%, 19/627) (Qin et al., 2021) in Tibetan antelopes. This difference may be due to differences in susceptibility of Tibetan antelope to different parasites. The prevalence of Cyclospora spp. in Tibetan antelopes (1.2%) was lower than in goats (1.9%) and sheep (3.1%) in Tamil Nadu, India (Basnett et al., 2018), which may be due to differences in susceptibility to Cyclospora spp. due to species differences. It was also lower than cattle in Shanxi (2.1%) (Wu et al., 2024) and Holstein cattle in Yunnan (2.5%) (Yang et al., 2023). Furthermore, this study identified statistically significant regional differences in the infection rate of Cyclospora spp. in Tibetan antelopes, possibly influenced by factors such as geographical location, local ecological and health conditions, and sample size.
Across seasons, the highest infection rate of Cyclospora spp. in Tibetan antelopes was observed in summer (3.3%), followed by autumn (1.2%), with no infections detected in spring. This pattern is linked to the life cycle of Cyclospora. Previous studies have shown that cyclosporiasis is closely associated with seasonal changes, local climate, and other environmental factors (Chacín-Bonilla, 2008; Frickmann et al., 2021; Helmy et al., 2006; Li et al., 2020). During the rainy season or summer, warm and humid conditions promote the formation of infectious Cyclospora oocysts and sporozoites, significantly increasing infection rates.
In 2020, the infection rate of Cyclospora spp. in Tibetan antelopes was 0.3%. No Cyclospora infections were detected in 2023, but in 2024, the rate rose to 3.3%. This rise in infection rates highlights the need for greater attention to Cyclospora spp. infections and underscores the importance of enhancing surveillance and testing efforts. The study has some limitations, including relatively small sample sizes for certain years, regions, and seasons. To address this, we plan to expand our sample collection in the future to validate our findings more effectively.
Currently, no molecular biological data on Cyclospora spp. from sheep or goats have been reported, and this study provides the first data on Cyclospora spp. isolates from Tibetan antelope. An analysis of the six Cyclospora spp. sequences identified in this study revealed that two of them share 99.4–99.8% homology with Cyclospora spp. (OP268225) from Holstein cattle in Yunnan, China, differing by 1–3 nucleotides. Three sequences share 97.8–99.4% homology with Cyclospora spp. (OP268233) from Holstein cattle in Yunnan, China, differing by 3–10 nucleotides. The remaining sequence exhibits 99.2% homology with Cyclospora spp. (PP930929) from cattle in Shanxi, China, with a difference of 4 nucleotides. These results indicate that the SSU rRNA gene of the Cyclospora spp. isolates from Tibetan antelope shows a certain degree of sequence variation. No C. cayetanensis was detected in this study, suggesting that Tibetan antelopes may not be hosts for C. cayetanensis.
Previous reports have identified the presence of C. cayetanensis in fecal samples from cattle, dogs, and cats (Chu et al., 2004; Qin et al., 2024; Wu et al., 2024; Yang et al., 2023). However, the detection of Cyclospora spp. in animal feces does not necessarily indicate that these animals are true hosts, as oocysts may pass through the digestive tract without causing infection (Wu et al., 2024). Although Cyclospora spp. was detected in fecal samples from Tibetan antelopes, it remains unclear whether Tibetan antelopes are the natural hosts. Further investigation, including small bowel biopsy, is needed to determine whether Tibetan antelopes serve as natural or paratenic hosts for Cyclospora spp.
Conclusion
This study is the first to report that the molecular prevalence of Cyclospora spp. in Tibetan antelopes in China is 1.2% (6/503), with significant correlations between Cyclospora spp. infection and both region and sampling year. Six sequences were obtained, showing homology ranging from 97.8 to 99.8%, all identified as belonging to Cyclospora spp. These findings provide basic data for assessing the transmission of Cyclospora spp. in Tibetan antelopes and provide new perspectives on its role and risk in zoonotic diseases.
Footnotes
Acknowledgments
The authors would like to extend their deepest gratitude to Chair Professor Hany Elsheikha, from University of Nottingham, for his invaluable assistance in revising and improving the article. His insightful feedback, expert advice, and meticulous attention to detail have significantly enhanced the quality of this work.
Authors’ Contributions
Conceptualization: J.J. and X.-X.Z. Funding acquisition: X.-X.Z., J.J., and H.-T.S. Investigation: W.-L.Y. and H.-L.G. Software: N.-Y.X. Methodology: S.-Y.Q. and N.-Y.X. Visualization: N.-Y.X. and Y.T. Resources: S.-Y.Q., J.-H.L., Z.-J.W., T.M., and J.-H.Z. Formal analysis: N.-Y.X. and S.L. Writing—original draft: N.-Y.X. and S.-Y.Q. Writing—review and editing: J.-H.L., Z.-J.W., T.M., J.-H.Z., W.-L.Y., H.-L.G., S.L., Y.T., H.-T.S., J.J., and X.-X.Z. All authors contributed to the article editing and approved the final article.
Data Availability
The generated sequences have been uploaded to the GenBank database with accession numbers PV053000–PV053005.
Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This work was supported by the National Key Research and Development Program of China (2021YFC2300903) and the Shandong Province Higher Education Institutions “Youth Innovation Team Plan” (2022KJ169).