
Abstract
Congenital heart disease (CHD) is the most common type of birth defect, affecting 1% of all live births. The recent exponential increase in the knowledge of medical genetics has revolutionized the understanding of CHDs during the past few decades. GATA4, a transcription factor, is involved in heart development. There are many contradictory reports on involvement of single-nucleotide polymorphisms (SNPs) of GATA4 in the manifestation of CHD. In view of this, an attempt has been made to analyze the known SNPs of GATA4 in Mysore patients with CHD. Of the 308 CHD patients recruited, 100 were screened for SNPs of GATA4 by MassARRAY, which identified 11 SNPs, of which 6 were found in both CHD cases and controls. The other 5 SNPs, c.278G>C (G93A), c.1207C>A (L403M), c.1232C>T (A411V), c.1295T>C (L432S), and c.1180C>G (P394A), were found only in CHD patients. Secondary structure analysis revealed that mutant proteins with the SNPs G93A, L403M, and L432S showed structural changes in their helix, sheet, and turn. Thus, these findings suggest the involvement of specific SNPs of GATA4 in the manifestation of CHD, reported for the first time in an Indian scenario. However, screening for a larger number of CHD patients would help us to establish genotype-phenotype correlation.
Introduction
C
The exponential increase in the knowledge of medical genetics has revolutionized the understanding of CHDs during the past few decades (Srivastava, 2001; Smitha et al., 2006). SNPs in GATA4 were identified as a genetic cause of nonsyndromic atrial septal defect (ASD) and VSD without conduction disturbances by studying large pedigrees with familial CHD (Garg et al., 2003; Garg, 2006). Various studies reported the occurrence of a very low frequency of SNPs of GATA4 in CHDs (Garg et al., 2003; Reamon-Buettner et al., 2007; Tomita-Mitchell et al., 2007; Posch et al., 2008; Zhang et al., 2008). Garg et al. (2003) identified the first two families with isolated congenital cardiac septal defects caused by inherited SNPs in the GATA4 (G296S and c.1075delG). Okubo et al. (2004) reported a Japanese family with autosomal dominant ASDII due to a heterozygous GATA4 deletion in an adjacent nucleotide upstream to the frame shift mutation (c.1074delC) reported by Garg et al. (2003). However, a preliminary study from our laboratory revealed that G296S is not responsible for cardiac septal defects in CHD patients of Mysore, South India (Ramegowda et al., 2007b). Further, two other studies have analyzed 16 families each, but only 2 of 16 families with multiple affected members carried SNPs of GATA4 (Hirayama-Yamada et al., 2005; Sarkozy et al., 2005; Reamon-Buettner et al., 2007). There are also reports that SNPs of GATA4 were found in other forms of CHDs (Zhang et al., 2008). However, in 42 patients with nonsyndromic atrio VSDs (AVSDs) (familial as well as sporadic), no SNPs of GATA4 have been detected (Sarkozy et al., 2005; Reamon-Buettner et al., 2007). In view of this, here an attempt has been made to analyze the known SNPs of GATA4, which were repeatedly reported in the literature in the patients with CHD in Mysore, India.
Materials and Methods
Three hundred eight clinically diagnosed CHD patients and 50 unrelated healthy controls were recruited for this investigation. A thorough clinical examination of the patients was carried out by expert doctors, including clinical history, pedigree analysis, physical examination, and an echocardiography. Informed consent was obtained from all participants in accordance with the Institutional Human Ethical Committee, University of Mysore, Cheluvamba Hospital and J.S.S. Hospital, Mysore, India. On the basis of echocardiography reports, 100 patients with severe septal defects, including patients with large defects ranging between 6 and 20 mm of ASD, VSD plus a cyanotic defect, patent ductus arteriosus (PDA), and also other complex forms were analyzed.
Peripheral whole-blood samples were collected in an EDTA vacutainer and stored at −80°C until further use. Genomic DNA was isolated using a Wizard genomic DNA purification kit (Promega). From the literature, 29 nonsynonymous SNPs (nsSNPs) of GATA4 were obtained, which were reported in other studies (Hirayama-Yamada et al., 2005; Reamon-Buettner and Borlak, 2005; Rajagopal et al., 2007; Reamon-Buettner et al., 2007; Schluterman et al., 2007; Tomita-Mitchell et al., 2007; Posch et al., 2008; Zhang et al., 2008; Hamanoue et al., 2009), using MassARRAY analysis, which was carried out by using a Sequenom-iPLEX® Gold SNP genotyping platform with Spectro CHIP® and MALDI-Time of Flight (TOF) Mass spectrometer (Sladek et al., 2007; Wilke et al., 2009). The sequence analysis and the secondary structures were predicted for the proteins derived from these 11 nsSNPs with the Chou Fasman secondary structure algorithm by using Accelrys gene software. The χ2 test was applied between cases and controls to ascertain the SNP differences.
Results
Human GATA4 located on chromosome 8p23.1-p22 consists of seven exons. GATA4 (MIM No. 600576) spans 50 kbp and is composed of 442 amino acids with two transactivation domains and two class IV zinc finger domains (N-terminal Zinc finger and a C-terminal zinc finger) along with a nuclear localization signal (Fig. 1). Table 1 presents the SNPs of GATA4 selected for the present study. Of the 50 known nsSNPs in GATA4, the 29 SNPs present in the coding exons of 2, 3, 4, 5, 6, and 7 were screened.
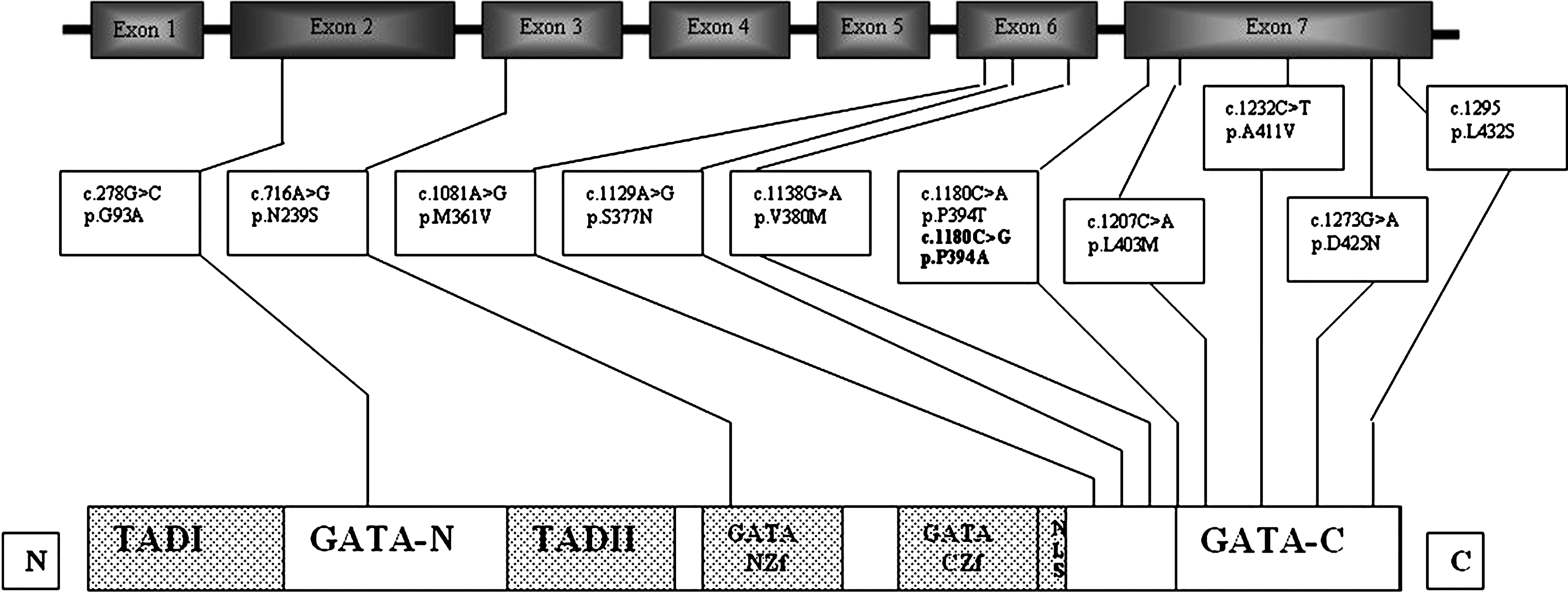
Genomic organization of GATA4 structure and single-nucleotide polymorphisms identified in the congenital heart disease patients of Mysore, India. Functional protein: domains 5; Transactivation domain I (TADI), Transactivation domain II (TADII), N-terminal zinc finger domain (NZf), C-terminal zinc finger domain (CZf) and nuclear localization signal region (NLS). GATA-N terminal region (GATA-N) and GATA-C terminal region (GATA-C).
SNP, single-nucleotide polymorphism.
Of the 308 CHD patients analyzed, 100 CHD patients were recruited for the analysis of SNPs of GATA4, which includes 33 patients with VSD, 12 patients with ASD, 3 patients with ASD-VSD, 4 patients with TOF, and 6 patients with PDA, and the remaining with other complex forms (Table 2). MassARRAY screening identified 11 nsSNPs (Fig. 1), of which 5, namely, c.1129A>G (S377G), c.1180C>A (P394T), c.1081A>G (M361V), c.1138G>A (V380M), and c.1273G>A (D425N) SNPs, were found in both CHD cases and controls (Table 2). One of the SNPs, c.716A>G (N239S), was not found in CHD cases but was present in controls. Another SNP, c.1081A>G (M361V), was found only in one patient with VSD in a heterozygous condition and also in one of the controls in homozygous condition.
p=0.002.
ASD, atrial septal defect; VSD, ventricular septal defect; PDA, patent ductus arteriosus; COA, coarctation of aorta; TOF, Tetralogy of Fallot; AVSD, atrioventricular septal defect; PHT, pulmonary hypertension; CCHD, complex CHD; SI, situs inversus; MS, mitral stenosis; TGA, transposition of gtreat arteries; TAPVC, total abnormal pulmonary venous connection; PAH, pulmonary artery hypertension; DORV, double outlet right ventricle; RV, right ventricle; SS, subaortic stenosis; PS, pulmonary stenosis; TA, truncus arteriosus, PA, pulmonary atresia.
Interestingly, χ2 analysis revealed that SNP c.1138G>A (V380M) was found significantly more frequently in CHD cases than controls. The remaining five SNPs were found only in CHD cases (Table 2). The SNP c.278G>C (G93A) was found only in one of the complex CHD (CCHD) patients with TOF associated with VSD, but not in controls. A novel SNP, c.1180 C>G (P394A), was observed only in cases with ASD, VSD, and ASD-VSD and not in any of the controls. The SNP c.1207C>A (L403M) was found only in one patient with ASD-VSD. The SNP c.1232C>T (D425N) was also found only in one CHD patient with AVSD. Only one patient with PDA showed c.1295T>C (L432S) SNP (Table 2).
Secondary structure analysis of proteins derived from these 11 nsSNPs revealed that mutant proteins of G93A showed structural changes in its helix, L403M showed structural changes in its helix and sheet, and L432S showed structural changes in its helix and turn (Fig. 2).
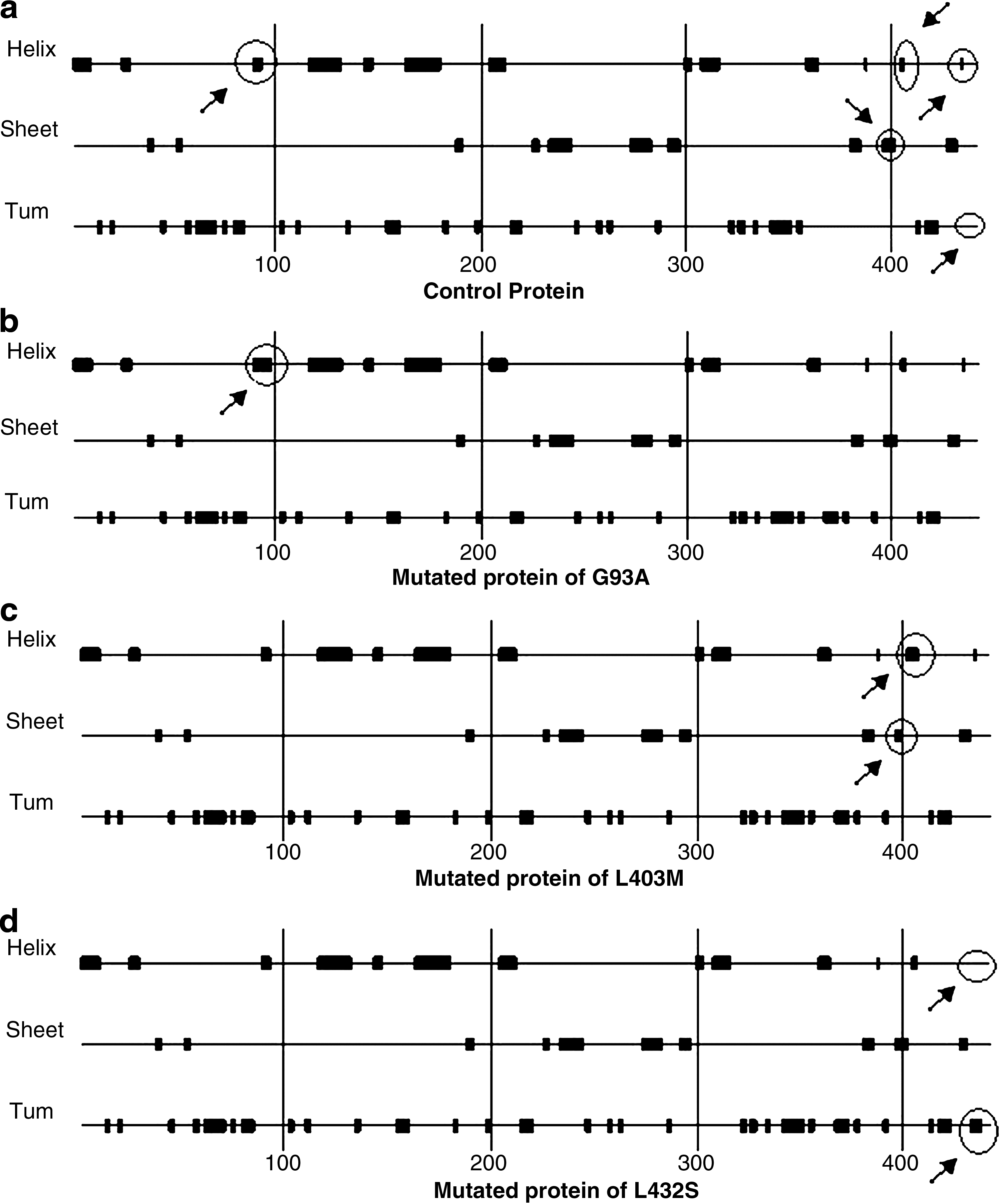
Chou Fasman secondary structure of GATA4 protein:
Discussion
Several GATA4 missense mutations were identified, particularly in patients with inherited ASDII (Hirayama-Yamada et al., 2005; Rajagopal et al., 2007; Tomita-Mitchell et al., 2007; Posch et al., 2008) and also in consanguineous patients with TOF (Nemer et al., 2006). Zhang et al. (2008) found many SNPs of GATA4 in different Chinese cohorts. In the present study, 11 nsSNPs of GATA4 were recorded, 5 of which are found only in CHD patients. Reamon-Buettner et al. (2007) reported the c.1081 A>G (M361V) SNP in 1 VSD case of the 80 CHD patients screened. In the present study, this SNP in heterozygous condition was found in one VSD case as well as in one control sample. Tomita-Mitchell et al. (2007) reported c.1273 G>A (D425N) in one ASD and one TOF case. This SNP was found in 2% of CHD as well as 6% of the controls. Another SNP, c.716 A>G (N239S), found only in one of the controls of this study was reported by Reamon-Buettner and Borlak (2005) in 1 VSD case of 68 CHD patients screened. These three SNPs were found in both controls and cases, with no impact on CHD, indicating that these are randomly distributed changes in the genome.
The SNP c.1180 C>A (P394T) was also observed in 8% of the cases including PDA, VSD, coarctation of aorta (COA), TOF, AVSD, pulmonary hypertension, and CCHD+TOF+VSD and also in about 10% of the controls. The database SNP (dbSNP) c.1129 A>G (S377G) (rs3729856) was observed in one CHD patient with situs inversus and in 8% of the controls. These two SNPs were also reported earlier in CHD cases as well as in controls (Zhang et al., 2006; Schluterman et al., 2007). Schluterman et al. (2007) reported c.1138 G>A (V380M) in 1 CHD patient of 157 patients screened and it was also reported in one of the controls. In this study, this SNP was found in 10% of the CHD patients including isolated ASD, VSD, COA, and pulmonary hypertension as well as in 4% of controls. Even though this SNP did not show any structural changes in protein, it was observed to be significantly greater in patients than controls, which is an interesting observation. However, in the same region, a homozygous SNP c.1180 C>G (P394A) was found in five CHD patients, interestingly all with septal defects and not in any of the controls. This is a unique observation, because it has not been reported in any of the previous studies; hence, it is a novel SNP. This SNP codes for the amino acid alanine instead of proline and is found in the C-terminal domain of the GATA4, which may disturb the transcriptional activation.
The heterozygous SNP c.1207 C>A (L403M) occurred only in a proband with osteum secundum ASD and perimembranous VSD (7 and 15 mm, respectively) and this SNP was not found in any controls. Rajagopal et al. (2007) found this SNP in one of the patients with ASD and hypoplastic right ventricle, of 107 patients screened. This SNP occurs in the C-terminal domain that is required for transcription activation. The SNP c.278 G>C (G93A) was found in the transactivation domain of the GATA-N terminal end of the GATA4 and this was observed only in one CCHD patient with malalignment of subaortic VSD (15 mm defect size) and TOF. Tomita-Mitchell et al. (2007) reported this SNP in 1 ASD case of the total 628 patients screened. They also identified another SNP, c.1232 C>T (A411V), in one VSD case. This SNP was found in one AVSD case under study, which was located near the C-terminal end of GATA4, a moderately conserved region for transactivation. The c.1295 T>C (L432S) SNP was reported by Wei-min et al. (2009) in two ASD, one VSD, and one TOF cases. However, we observed it only in one patient with PDA. The protein derived from this SNP showed secondary structural changes in the helix and turn, indicating significance in the manifestation of CHD (Fig. 2).
Thus, this study indicates that five SNPs were found only in CHD patients and another six were present in both cases and controls. The occurrence of SNPs of GATA4 in CHD patients of Mysore is more with severe septal defects as well as cyanotic PDA. Most of the SNPs of GATA4 were found in the medium- and large-sized septal defects. These SNPs may affect the normal pathway of interaction between GATA4 and other genes involved in heart development. Thus, these findings suggest the involvement of specific SNPs of GATA4 in the manifestation of CHD, reported for the first time in an Indian scenario. However, screening for large number of CHD patients would help us to better establish a genotype-phenotype correlation.
Footnotes
Acknowledgments
The authors thank the Council for Scientific and Industrial Research, New Delhi, Government of India [No. 27 (0156)/06/EMR-II dated 19.10.2006] for the financial support. The authors are grateful to all the patients and families who participated in this investigation, to doctors and PG students for their kind support, to Professor and Chairman, DOS in Zoology, University of Mysore, and also to Prof. H.A. Ranganath for his encouragement.
Disclosure Statement
No competing financial interests exist.