
Abstract
Aims:
Increasing evidence links the abnormal expression of microRNAs and ATP-binding cassette subfamily C member 4 (ABCC4) with tumor development and progression, as well as with chemoresistance. Our aims were to determine the therapeutic potential of targeting both miR-124-3p and ABCC4 in breast cancer cells and to determine if duel targeting increased their sensitivity to chemotherapeutic drugs, in vitro.
Materials and Methods:
The expression of the ABCC4 protein and miR-124-3p were detected, respectively, by immunohistochemical staining and quantitative real-time polymerase chain reaction in breast cancer tumor tissue, MCF-7 and MCF-7-ADR cell lines. Suppression of ABCC4 expression and miR-124-3p overexpression were performed in MCF-7-ADR cell lines. Western blot assays were used to detect expression of ABCC4 and permeability glycoprotein 1/multi-drug resistance protein 1 (P-gp) in cells. Cell Counting Kit-8, flow cytometry, transwell, and scratch assays were conducted to detect cell proliferation, cell cycle, invasion, and migration of cells.
Results:
We found that ABCC4 protein expression was significantly increased, while the miR-124-3p level was significantly decreased in breast cancer tissue and cell lines. Tumor size and clinical tumor node metastasis stage were significantly correlated with elevated expression of ABCC4 and decreased expression of miR-124-3p. Interestingly, ABCC4 expression was significantly increased in MCF-7-ADR cells, while miR-124-3p level was significantly decreased compared with MCF-7 cells. The inhibition of ABCC4 and miR-124-3p overexpression both led to a significant decrease in cell proliferation, invasion, and migration of MCF-7-ADR cells, and combination of suppression of ABCC4 with miR-124-3p overexpression had a synergistic inhibitory effect. Our results further demonstrated that inhibition of ABCC4 expression and overexpression of miR-124-3p significantly enhanced the sensitivity to adriamycin (ADR) in MCF-7-ADR cells, and that simultaneous dual-targeting of miR-124-3p and ABCC4 had a stronger promotive effect on the sensitivity to ADR in MCF-7-ADR cells. Moreover, western blot analysis showed that miR-124-3p overexpression significantly inhibited P-gp expression in MCF-7-ADR cells.
Conclusion:
Our data demonstrate that the combination of downregulation of ABCC4 with overexpression of miR-124-3p significantly increased sensitivity to ADR in MCF-7-ADR cells. This finding suggests that similar dual targeting may serve as a means to enhance therapies for drug-resistant breast cancers.
Introduction
Breast cancer has become the most common malignant cancer and the primary cause of cancer-related mortality in women all over the world (Zhu et al., 2018). Currently, cytotoxic chemotherapy has dramatically improved survival rate of breast cancer patients. However, chemotherapy resistance defined as multidrug resistance (MDR) is the major obstacle to the effective therapy of cancers, including breast cancer, which may possibly result in tumor relapse and the failure of therapy (Deng et al., 2014; Fan et al., 2017). Although adriamycin (ADR) is one of the most common chemotherapeutic agents used for breast cancer therapy, serious side effects and drug resistance lead to its limited clinical application (Deng et al., 2014). Thus, a better understanding of the molecular mechanism underlying drug resistance is urgently needed for providing new appropriate therapeutic approaches to increase chemotherapy efficacy and improve the clinical outcome of breast cancer patients.
Drug transporters are reported to play an important part in drug resistance, and ATP-binding cassette subfamily C member 4 (ABCC4), a MDR-associated protein, is one of the most completely studied transporters concerned with drug resistance (Vogt et al., 2018). Increasing evidence has disclosed that ABCC4 is a transporter decreasing absorption and enhancing drug export, including anticancer drugs, which are correlated with the recurrence, development, and chemotherapeutic susceptibility of multiple cancers (Murray et al., 2017; Tsukamoto et al., 2017). In neuroblastoma patients, high ABCC4 protein levels have a close relationship with decreased overall survival, and knockdown of ABCC4 significantly suppresses the growth of human neuroblastoma cells in vitro, impairs neuroblastoma xenografts growth, and sensitizes to the ABCC4 substrate drug irinotecan (Murray et al., 2017). In human gastric cancer, repression of ABCC4 expression can suppress proliferation of multidrug-resistant gastric cancer cells and enhance gastric cancer cell sensitivity to chemotherapeutic drugs (Zhang et al., 2015).
microRNAs (miRNAs) are a group of endogenous, small noncoding RNAs with ∼22 nucleotides in length that specifically bind to the 3′-untranslated region of mRNA of the target genes, resulting in mRNA degradation or translational inhibition (Hu et al., 2018b). It has been well known that they are involved in various biological and pathological processes, including cell proliferation, metastasis, differentiation, and apoptosis (Fan et al., 2017). Recently, several researches have revealed that the dysregulation of miRNAs is associated with the sensitivity of cancer cells to chemotherapy. For example, miR-221, miR-27a, and miR-199a-3p are usually dysregulated in breast cancer, which contributes to cisplatin resistance (Ye et al., 2016; Zhou et al., 2016; Fan et al., 2017). miR-125b regulates the drug-resistance of breast cancer cells to doxorubicin by targeting HAX-1 (Hu et al., 2018a). Chen et al. have found that increased miR-124 expression reduced DNA strand-break (SB) repair, which may increase sensitivity to SB-inducing chemotherapeutic agents, in breast cancer patients receiving chemotherapy (Chen et al., 2015).
In the present study, our aim is to propose a new approach in targeted therapy of cancer: the simultaneous, dual targeting of two single molecules, miR-124-3p, and ABCC4. Here, we demonstrated that the combination of miR-124-3p overexpression and ABCC4 knockdown can provide high sensitivity of breast cancer cells to ADR, which may serve as a novel ideal therapeutic target for reversing the drug resistance of breast cancer.
Materials and Methods
Samples
Surgically resected paired breast cancer and normal adjacent tissues were obtained from 40 patients who were all diagnosed with invasive ductal breast carcinoma and underwent breast-conserving surgery from May 2014 to May 2016. The surgically removed tissues were quickly frozen in liquid nitrogen until further analysis. All protocols concerning the use of patient samples in this study were approved by the Research Ethics Committee of the First Affiliated Hospital of Zhengzhou University. A signed informed consent was obtained from all participants.
Cell lines and culture
The MCF-10A mammary epithelial cells, MCF-7 breast cancer cells, and HEK 293T cells were obtained from Beijing Beina Chuanglian Biotechnology Research Institute (Beijing, China). Multidrug-resistant MCF-7-ADR human breast cancer cells were constructed in our laboratory. The cells were grown in RPMI 1640 medium supplemented with 10% fetal bovine serum (FBS; Invitrogen, Carlsbad, CA) in a humidified 5% CO2 atmosphere at 37°C. To maintain the MDR phenotype, 0.5 μg/mL of ADR was added to the culture of MCF-7-ADR cells and removed 7 days before the experiment. HEK 293T cells were cultured in Dulbecco's modified Eagle's medium (DMEM; HyClone, Logan, UT) with 10% FBS, and 1% penicillin-streptomycin at 37°C in 5% CO2.
Immunohistochemical analysis
Immunohistochemical (IHC) analysis was performed on 4% paraformaldehyde-fixed, paraffin-embedded 3 μm sections of breast cancer tissue collected. Sections were deparaffinized in xylene for 15 min, rehydrated in graded ethanol, and boiled in antigen retrieval solution for 10 min, at 95°C, followed by incubation with 3% hydrogen peroxide to block endogenous peroxidase for 5 min. After blocked with 30 μL of 5% bovine serum albumin (Abcam, Cambridge, UK), sections were incubated with rabbit anti-ABCC4 (1:1200; Cell Signaling Technology, Inc., Hitchin, UK), overnight at 4°C, and then were incubated with a horseradish peroxidase (HRP)-conjugated anti-rabbit secondary antibody (1:500; Cell Signaling Technology, Inc.) at room temperature for 25 min. Sections were incubated with diaminobenzidine (DAB; Sigma-Aldrich, St. Louis, MO), counterstained with hematoxylin (Richard Allan Scientific, Kalamazoo, MI), and examined and photographed with a fluorescence microscope (Olympus BX41; Olympus Corporation, Japan). Staining intensities were classified into four grades of intensity as follows: (−) negative; (+) weakly positive, (++) moderately positive, and (+++) strongly positive.
Quantitative real-time polymerase chain reaction analysis
Total RNA was extracted from cultured cells tissues using the TRIzol reagents (Invitrogen) in accordance with the manufacturer's protocol. For detection of mRNA and miRNA expression, reverse transcription was performed using the PrimeScript RT Reagent Kit (Takara Bio, Inc., Otsu, Japan). The relative expressions of ABCC4 and miR-124-3p were determined using the SYBR-Green PCR Master Mix Kit (Takara) and TaqMan MicroRNA Assay Kit (Applied Biosystems, Foster City, CA) on an ABI7500 system (Applied Biosystems), respectively. The primer sequences used in this study are as follows: ABCC4: sense: 5′-GGATCCAAGAACTGATGAGTTAAT-3′, antisense: 5′-TCACAGTGCTGTCTCGAAAATAG-3′; miRNA-124-3p: sense: 5′-GCGTAAGGCACGCGGTGAATGCC-3′, antisense: universal primers (in the kit); GAPDH: sense: 5′-CCTCAAGATTGTCAGCAAT-3′, antisense: 5′-CCATCCACAGTCTTCTGAGT-3′; U6: 5′-CTCGCTTCGGCAGCACATATACT-3′, antisense: 5′-ACGCTTCACGAATTTGCGTGTC-3′. The relative ABCC4 and miR-124-3p levels were normalized to GAPDH and U6 snRNA, respectively. The relative expression level was calculated using 2−ΔΔCt method.
Western blot assay
Cultured cells were collected, washed twice with phosphate-buffered saline (PBS), and lysed in RIPA buffer. Protein concentration was assessed using the Bradford method. Thirty micrograms of proteins were separated by 10% SDS-PAGE and then electrophoretically transferred onto a polyvinylidene difluoride membrane (Millipore, Bedford, MA). The blots were incubated in 1 × PBS containing 5% nonfat dry milk for 1 h. Following washing, blots were incubated with primary antibodies overnight at 4°C. Blots were exposed to HRP-conjugated secondary antibody for 2 h and were visualized using the enhanced chemiluminescence detection kit (Sigma). The following primary antibodies in this study were used: rabbit anti-ABCC4, rabbit anti-P-gp (1:200; Santa Cruz), and the second antibody.
Construction of the recombinant lentiviral vectors and lentivirus packaging
DNA fragment containing the miR-124-3p sequence was amplified using the extracted genome DNA of HEK 293T cells as template and cloned into lentiviral vector (pCDH-CMV-MCS-EF1-copGFP-T2A-Puro; System Biosciences Inc, Mountain View, CA) to generate the recombinant plasmid (pLenti-miR-124-3p). ABCC4 shRNA (inhibiting ABCC4 expression) was synthesized by Shanghai Shenggong Biological Engineering Co., Ltd. and cloned into lentiviral vector pLenti-hU6-GFP-Puro (Shanghai Shenggong Biological Engineering Co., Ltd.) to obtain the recombinant plasmid (pLenti-sh-ABCC4). The pLenti-miR-124-3p plasmid was digested by Not I enzyme to produce the sticky ends, respectively, and sh-ABCC4 fragment was amplified using the pLenti-sh-ABCC4 plasmid as template. Then, the two plasmids were connected by seamless junctions using Ready-to-Use Seamless Cloning Kit (Shanghai Shenggong Biological Engineering Co., Ltd.) to gain the recombinant plasmid pLenti-miR-124-3p+sh-ABCC4.
Lentiviral packing was performed as previously described (Song et al., 2017). In brief, the 293T cells were cultured for 48 h and then cotransfected with the recombinant lentiviral vector and two auxiliary packaging plasmids (pCMV-Δ8.2 and pCMV-VSV-G). Lentivirus particles carrying miR-124-3p, sh-ABCC4, or miR-124-3p+sh-ABCC4 were harvested by centrifugation at 3000 rpm for 5 min at 4°C after 48 h. All lentiviral constructs also contained a puromycin resistance sequence for drug screening. The high-concentration lentiviral concentrate was used to infect the MCF-7-ADR cells.
Lentiviral transfection of the MCF-7-ADR
To obtain MCF-7-ADR cells stably expressing miR-124-3p, inhibiting ABCC4, or expressing miR-124-3p and inhibiting ABCC4, MCF-7-ADR cells were infected with three lentivirus particles (LV-miR-124-3p, LV-sh-ABCC4, or LV-miR-124-3p+sh-ABCC4, respectively) in the presence of 8 μg/mL polybrene (GeneCopoeia, Guangzhou, China) for 12 h. The culture medium was removed during 10 min at 1000 rpm in the horizontal centrifuge. Then, the fresh culture medium containing 2% FBS was added each well, and cells were cultured for another 72 h. The cells were then cultured in medium with 10 μg/mL puromycin (Sigma-Aldrich) to gain the stable cells. Quantitative real-time polymerase chain reaction (qRT-PCR) analysis and western blot analysis were used to determine the expression miR-124-3p and ABCC4 in the stable cells.
Cell Counting Kit-8 assay
For cell proliferation assay, MCF-7-ADR cells (4 × 103/well) were seeded in 96-well plates. The number of viable cells was detected using a Cell Counting Kit-8 (CCK-8; Dojindo, Tokyo, Japan) according to the manufacturer's instructions. Ten microliters of CCK-8 assay solution was added to each well after incubation for 1, 2, 3, 4, or 5 days. Afterward, absorbance was detected using enzyme immunoassay analyzer (Thermo Fisher Scientific, Inc.) at 450 nm.
For cytotoxicity assessment by means of the CCK-8 assay, MCF-7-ADR cells (4 × 103/well) were seeded in 96-well plates. After cells were cultured for 24 h, cells were treated with different doses of ADR (0.25, 0.5, 1, 2, 4, 8, 16, and 32 μg/mL). Cells were cultured for 72 h, and then the number of viable cells was detected using a CCK-8.
Cell cycle distribution analysis
For cell cycle distribution analysis, MCF-7-ADR cells (2 × 105/well) were seeded in six-well plates. Cells were cultured for 48 h. Cells were trypsinized, harvested, and then washed twice in cold PBS. Cells were fixed in 1 mL of cold 70% ethanol (in PBS) overnight at 4°C. Cells were washed once in PBS and then cells were resuspended in 500 μL of 50 mg/mL propidium iodide staining solution containing 20 μL RNase A solution and incubated for 30 min in the dark.
Transwell assay
Invasion assay was performed as described previously (Ni et al., 2015). In brief, before the start of the experiment, BioCoat Matrigel Invasion Chambers (BD Biosciences) with 8-μm pore polycarbonate membranes were pretreated with serum-free RPMI-1640 medium at 37°C for 2 h. Two hundred microliters of cell suspension of MCF-7-ADR cells (4 × 104/well) was plated into the upper chamber and 500 μL of RPMI-1640 medium with 10% FBS as chemoattractant to each lower chamber. Furthermore, cells were cultured at 37°C for 36 h, and a cotton swab was used to remove the noninvasive cells on the upper surface. The invasive cells on the lower surface of the membrane were then fixed in 100% methanol for 10-15 min, air-dried, and stained with 1% crystal violet. Cells from five microscopic fields were photographed and counted.
Scratch assays
For migration assay, 200 μL of cell suspension of MCF-7-ADR cells (4 × 104/well) were plated into 24-well plates until they reached 70-80% confluence. At the initial time (T0), a cross-shaped scratch was made on the cell monolayer using a sterile 10 μL micropipette tip and photographed and measured by taking pictures at the beginning. After 48 h (T48), cells were photographed at the end of the experiment at the times indicated using 40 × magnification. The scratched area was quantified using ImageJ 1.44p software (NIH) to draw the cell-free region limits in each case.
Statistical analysis
All analyses were performed using SPSS 19.0. Statistical analysis was conducted in SPSS 19.0 (SPSS, Inc., Chicago, IL). All data were presented as the mean ± standard deviation. Statistical significance was detected by t-test for two groups and one-way analysis of variance for more than two groups. p < 0.05 was considered significant.
Results
ABCC4 expression was increased and miR-124-3p level was decreased in breast cancer tissue and cells
The expression of ABCC4 and miR-124-3p was detected in the tumor tissues of 40 patients with breast cancer and cells using IHC staining and RT-qPCR. As shown in Figure 1A and B, the ABCC4 expression was significantly increased in breast cancer tissues compared with normal adjacent tissues, while the miR-124-3p levels were significantly downregulated. Furthermore, we analyzed the relationship between ABCC4 and miR-124-3p expression as well as clinical features of breast cancer patients. The results showed that tumor size and clinical tumor node metastasis (TNM) stage were significantly related with the expression of high ABCC4 and low miR-124-3p, whereas there was not any obvious correlations between the ABCC4 and miR-124-3p levels and clinical characteristics in ages, HER-2 expression, and histological subtype (Table 1). Interestingly, protein and mRNA levels of ABCC4 expression were significantly higher in MCF-7 cells than that in MCF-10A cells, and ABCC4 expression was significantly increased in multiresistant MCF-7-ADR cells compared with MCF-7 cells (Fig. 1C). Moreover, miR-124-3p levels were obviously reduced in MCF-7 cells than those in MCF-10A cells, and miR-124-3p expression was markedly decreased in MCF-7-ADR cells compared with MCF-7 cells (Fig. 1D). These results suggested that both ABCC4 and miR-124-3p may be involved in the regulation of ADR sensitivity in breast cancer.
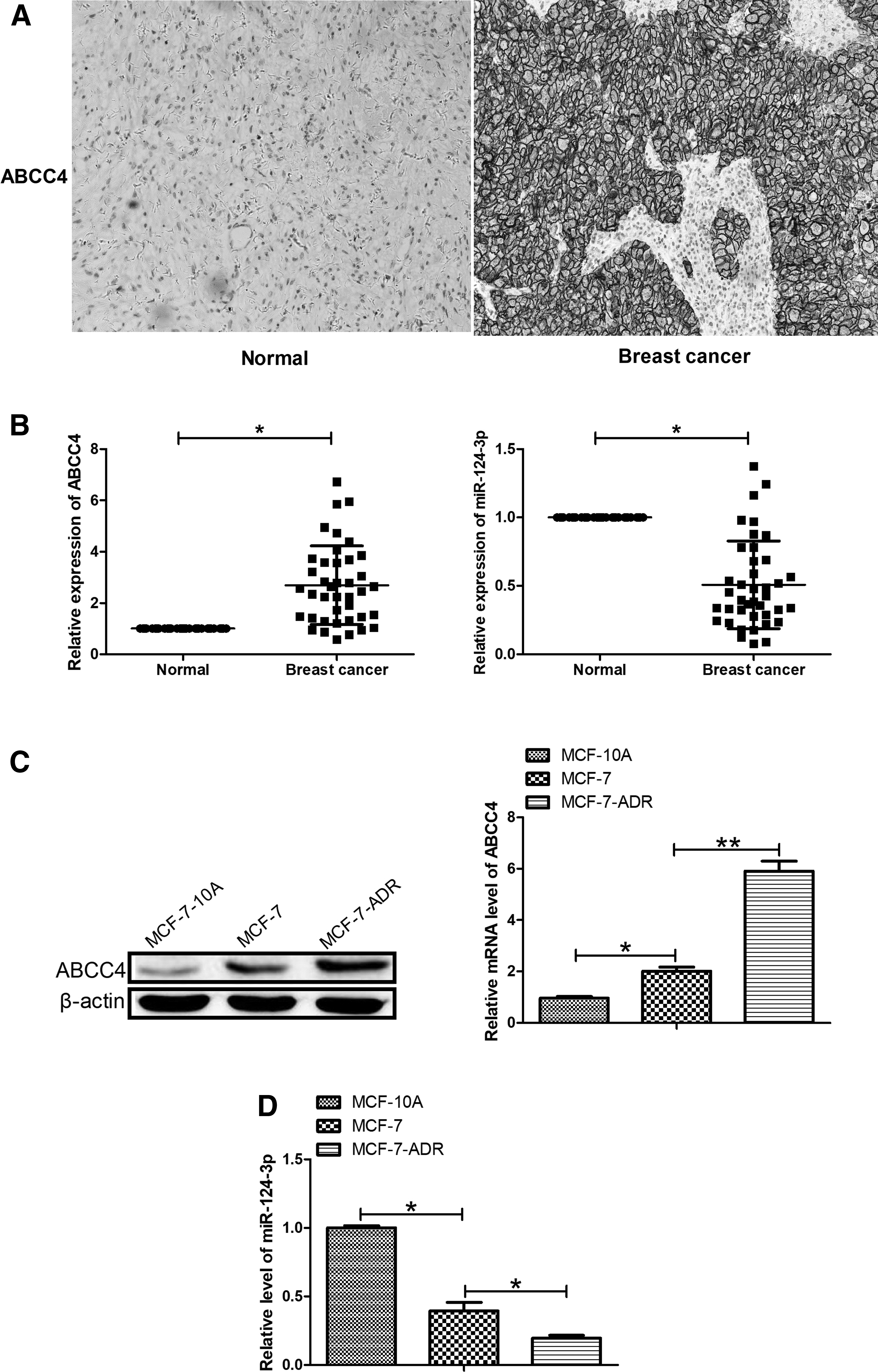
ABCC4 and miR-124-3p levels in breast cancer tissue and cells.
Correlation Between miR-124-3p and ATP-Binding Cassette Subfamily C Member 4 Expression as Well as Clinicopathological Features of Breast Cancer Patients
ABCC4, ATP-binding cassette subfamily C member 4; TNM, tumor node metastasis.
The construction of stable miR-124-3p, sh-ABCC4, and miR-124-3p+sh-ABCC4 MCF-7-ADR cells
MCF-7-ADR cells were transfected with lentivirus particles (LV-miR-124-3p, LV-sh-ABCC4, or LV-miR-124-3p+sh-ABCC4, respectively), and then, GFP protein expression was observed under a fluorescence microscope, and the result indicated a successful transfection (Fig. 2A). qRT-PCR and western blot analyses showed that the mRNA and protein levels of ABCC4 were both significantly lower in MCF-7-ADR-miR-124-3p and MCF-7-ADR-sh-ABCC4 cells than those in the MCF-7-ADR cells, while ABCC4 levels were significantly decreased in MCF-7-ADR-miR-124-3p+sh-ABCC4 compared with the MCF-7-ADR-miR-124-3p and MCF-7-ADR-sh-ABCC4 cells (Fig. 2B, C). Moreover, results from qRT-PCR demonstrated that miR-124-3p levels were significantly increased in MCF-7-ADR-miR-124-3p cells compared with MCF-7-ADR cells, and miR-124-3p levels in MCF-7-ADR-miR-124-3p+sh-ABCC4 cells were higher than those in MCF-7-ADR and MCF-7-ADR-sh-ABCC4 cells (Fig. 2B). These data suggested that we successfully constructed stable miR-124-3p, sh-ABCC4, and miR-124-3p+sh-ABCC4 MCF-7-ADR cells.
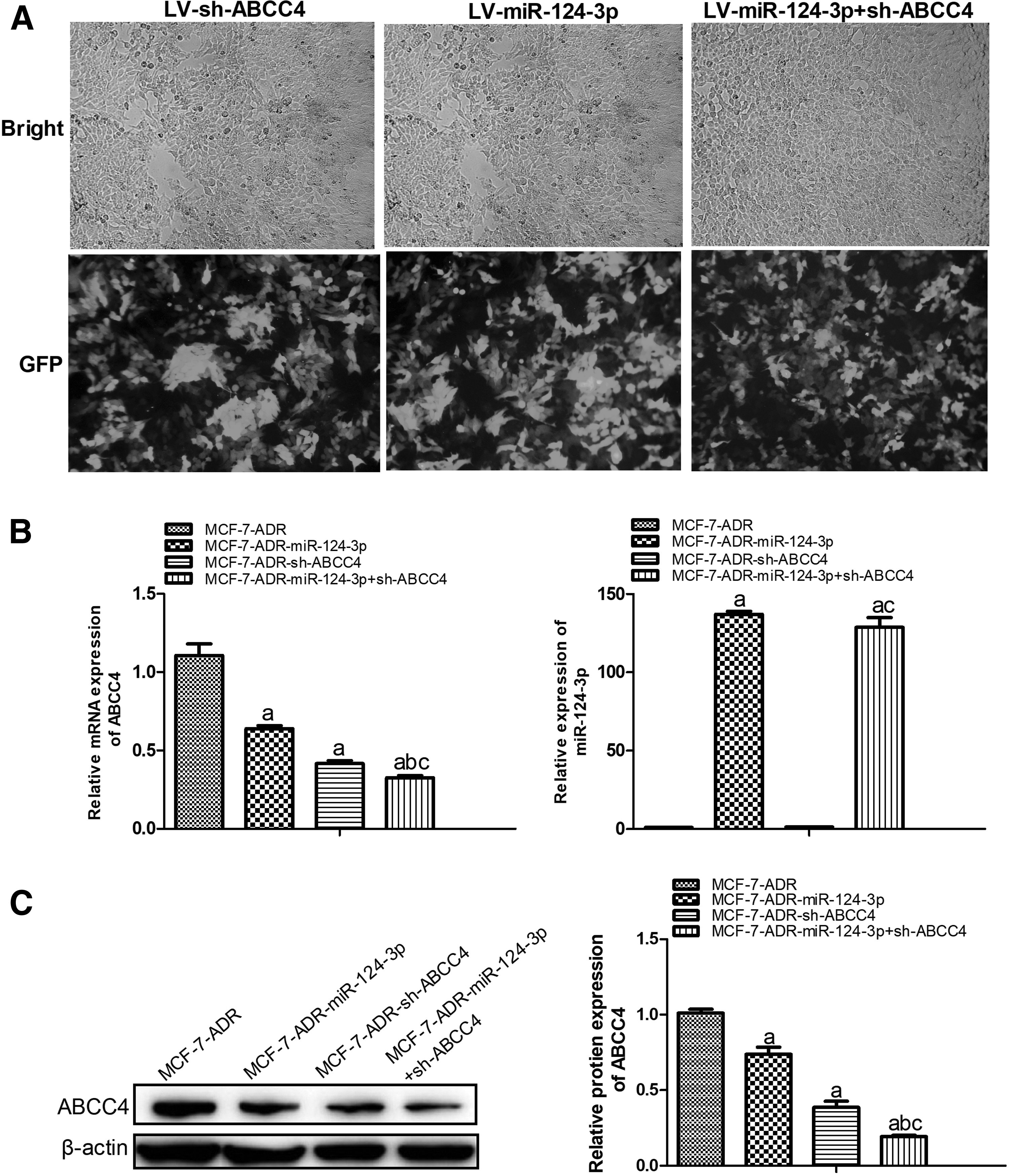
Construction of stable MCF-7-ADR cells transfected with miR-124-3p, sh-ABCC4, or miR-124-3p+sh-ABCC4.
Combination of miR-124-3p overexpression and ABCC4 knockdown inhibited proliferation of MCF-7-ADR cells
To confirm the effects, miR-124-3p overexpression or ABCC4 knockdown alone or combinedly on MCF-7-ADR cell growth, CCK-8 assay and cell cycle distribution analysis were performed. As presented in Figure 3A and B, both miR-124-3p overexpression and ABCC4 knockdown significantly inhibited cell proliferation and hindered the normal cell cycle process of MCF-7-ADR cells. Especially, the combination of miR-124-3p overexpression and ABCC4 knockdown had more obvious inhibitory effects on MCF-7-ADR cell proliferation.
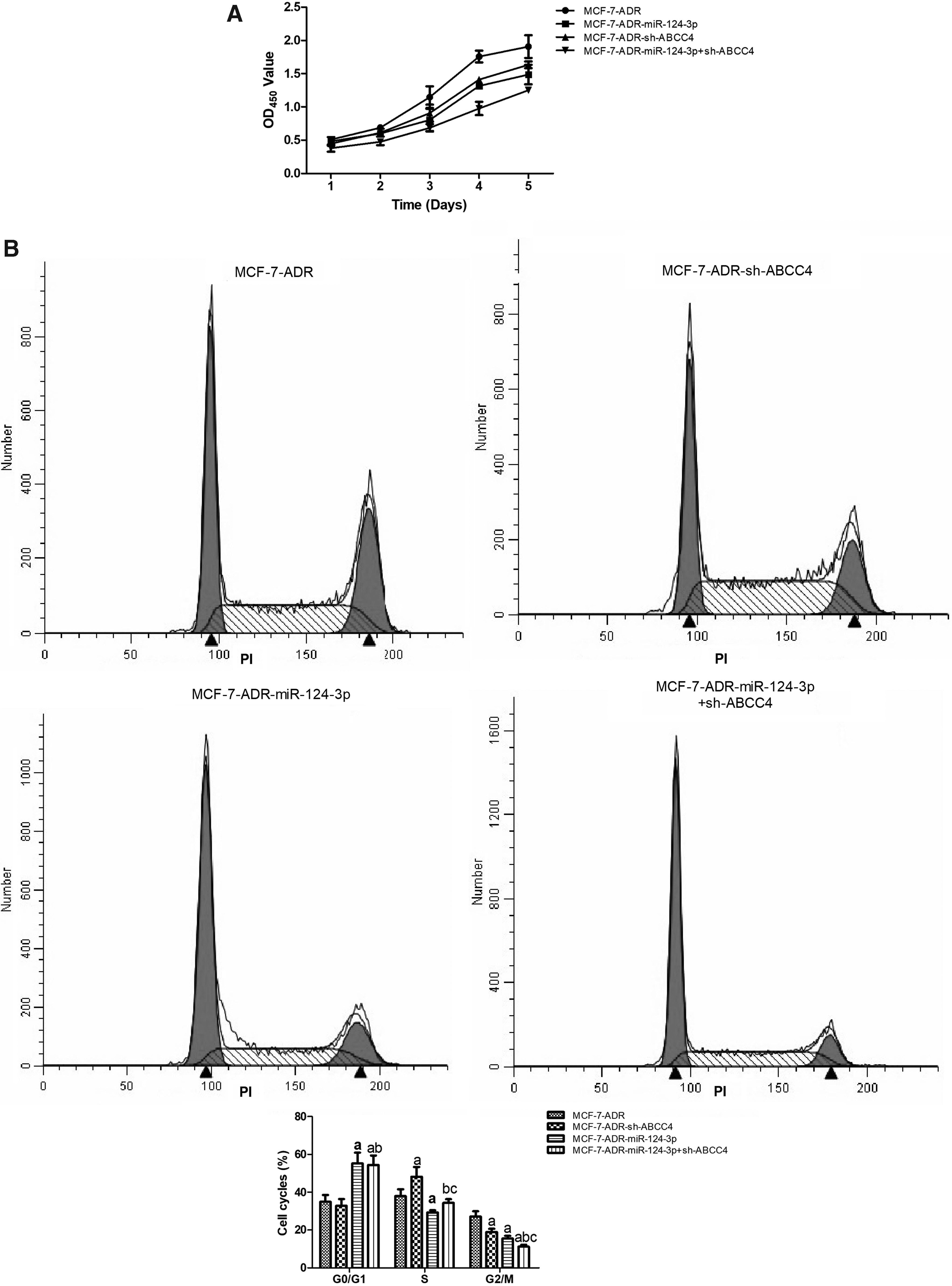
Combination of miR-124-3p overexpression and ABCC4 knockdown inhibited proliferation of MCF-7-ADR cells.
Combination of miR-124-3p overexpression and ABCC4 knockdown inhibited invasion and migration of MCF-7-ADR cells
To investigate the role of miR-124-3p overexpression or ABCC4 knockdown alone or combinedly in invasion and migration of MCF-7-ADR cells, transwell and scratch assays were conducted. As Figure 4A and B show, miR-124-3p overexpression or ABCC4 knockdown can suppress invasion and migration of MCF-7-ADR cells. In addition, combination of miR-124-3p overexpression and ABCC4 knockdown had more obvious inhibitory effects on MCF-7-ADR cell invasion and migration.
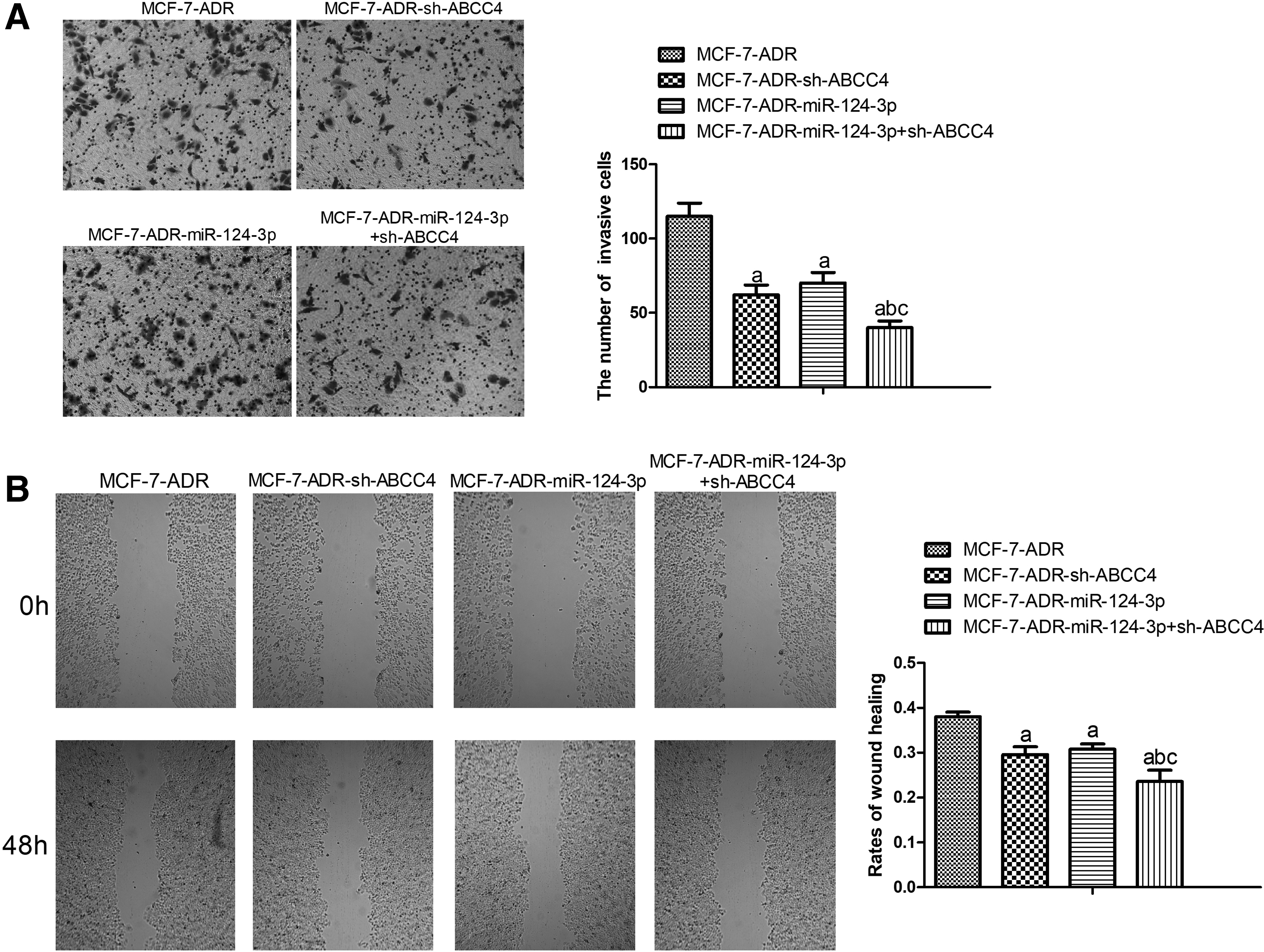
Combination of miR-124-3p overexpression and ABCC4 knockdown inhibited invasion and migration of MCF-7-ADR cells.
Combination of miR-124-3p overexpression and ABCC4 knockdown enhanced sensitivity to ADR in MCF-7-ADR cells
Next, we explored whether miR-124-3p overexpression or ABCC4 knockdown alone or combinedly contributes to sensitivity to ADR of MCF-7-ADR cells. CCK-8 assays exhibited that miR-124-3p overexpression or ABCC4 knockdown significantly promoted the sensitivity to ADR in MCF-7-ADR, and combination of miR-124-3p overexpression and ABCC4 knockdown had more obvious promotive effect on drug sensitivity in MCF-7-ADR cells, compared with miR-124-3p overexpression and ABCC4 knockdown MCF-7-ADR cells (Fig. 5A). In addition, the expression of drug resistance-related protein P-gp was detected by western blot assay. The results showed that miR-124-3p overexpression significantly inhibited protein expression of P-gp in MCF-7-ADR cells, while ABCC4 downregulation had no impact on P-gp expression (Fig. 5B).
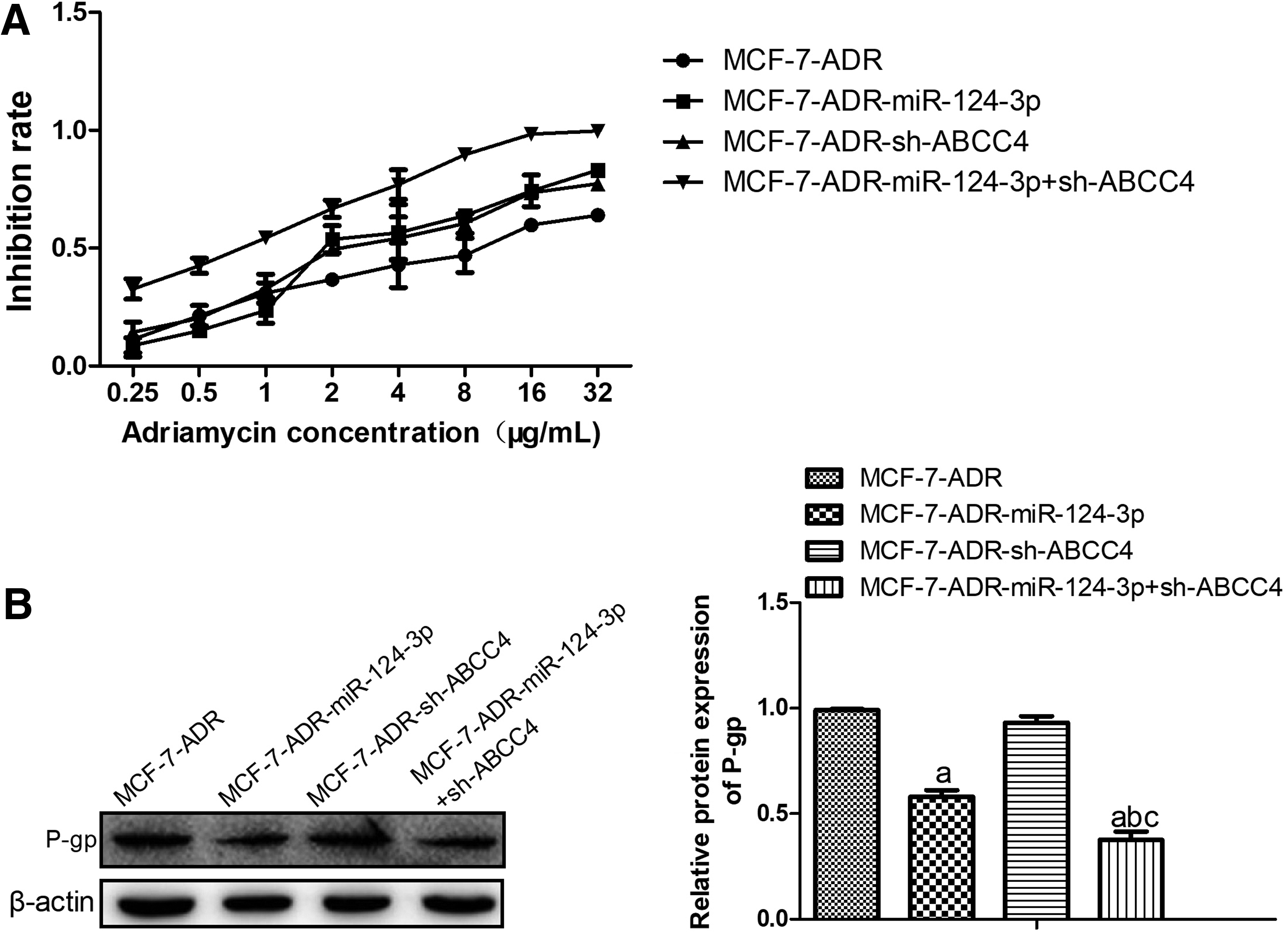
The combination of miR-124-3p overexpression and ABCC4 knockdown enhanced sensitivity of resistant breast cancer cells (MCF-7-ADR) to ADR drug.
Discussion
Despite chemotherapeutic agents and combined chemotherapy regimens have been applied to significantly improve survival outcome of patients with breast cancer, drug resistance is still one of the main reasons of therapy failure (Liu et al., 2016; Zhang et al., 2016). ABC transporters efflux drugs outside of tumor cells, which causes the occurrence of chemoresistance and thus weakening the therapeutic effect (Pan et al., 2017). Several studies previously identified that ABCC4 as an alluring therapeutic target in cancers, including breast cancer (Kochel et al., 2013; Murray et al., 2017). Murray et al. have reported that ABCC4 knockdown slows the progression of neuroblastoma xenografts and sensitizes neuroblastom tumor to irinotecan in vivo (Murray et al., 2017). In human pancreatic cancer, ABCC4 has been reported to be overexpressed and affect the sensitivity of pancreatic cancer cells to 5-fluorouracil (5-FU) and is expressed specifically in 5-FU-resistant cells (He et al., 2015). ADR is an antimitotic drug applied to many malignant tumors, including breast cancer (He et al., 2015). In our study, we found that ABCC4 expression was significantly increased in breast cancer tissues and cells. Tumor size and clinical TNM stage were significantly related with the expression of high ABCC4. Interestingly, we also found that ABCC4 expression was significantly increased in multiresistant MCF-7-ADR cells. Furthermore, our data suggested that ABCC4 downregulation suppressed cell proliferation, invasion, and migration, hindered the normal cell cycle process, and promoted the sensitivity to ADR of MCF-7-ADR.
Also, accumulating evidence shows that miRNAs have a critical role in the initiation and progression of types of human cancer, including breast cancer (Ye et al., 2016; Ding et al., 2018; Zhong et al., 2018). Importantly, recent research has indicated that miRNAs are also related with the chemotherapy on cancers. For example, it has been reported that miRNA-17-5p is increased in chemoresistant patients and its overexpression promotes chemotherapeutic drug resistance and tumor metastasis of colorectal cancer by repressing PTEN expression (Fang et al., 2014). It has been presented by Niu et al. that suppression of miR-181a expression enhances chemotherapeutic resistance and metastasis in a triple-negative breast cancer (TNBC) orthotopic model (Niu et al., 2016). miR-1291 has been demonstrated to modulate cellular drug disposition and chemosensitivity through regulation of ABCC1 expression in PANC-1 cells (Pan et al., 2013). In breast cancer cells, miR-621 has been reported to enhance chemosensitivity of breast cancer cells to PTX/CBP chemotherapy by suppressing FBXO11-depedent inhibition of p53 (Xue et al., 2016).
miR-124-3p was determined as a potential tumor-suppressive miRNA and downregulated in many cancers, including breast cancer. Recently, Zhang et al. have demonstrated that miR-124-3p is decreased in breast cancer tissues and breast cancer cells lines and reduced miR-124-3p prompted breast cancer progression by targeting Beclin-1 (Zhang et al., 2016). Consistent with that, our study also disclosed that miR-124-3p level was decreased in breast cancer tissue and cells. Tumor size and clinical TNM stage were significantly related with the expression of low miR-124-3p. Moreover, miR-124-3p expression was markedly decreased in MCF-7-ADR cells compared with MCF-7 cells, which implied that miR-124-3p might be involved in drug resistance in breast cancer. Increasing evidence has showed that miRNAs play vital roles in regulating cancer cell proliferation, survival, and sensitivity to chemotherapy (Xue et al., 2016). In our present investigation, we found that miR-124-3p overexpression inhibited cell proliferation, invasion, and migration, hindered the normal cell cycle process, and enhanced the sensitivity to ADR in MCF-7-ADR.
Several studies have shown that in the gene therapy of pancreatic cancer, the combination of multigene therapy tends to be more effective than the single-gene therapy strategy (Xu et al., 2010). Flores-Perez et al. have found that inhibition of both ANGPT1 and TGFBR2 genes inhibits angiogenesis in breast cancer, which may provide a promising treatment in breast cancer (Flores-Perez et al., 2016). Holler et al. have shown that dual targeting of mTORC1 and Akt1 inhibits repair of DNA-double-strand break, leading to radiosensitization of solid tumor cells (Holler et al., 2016). Rhodes L et al. have pointed out that dual regulation by miRNA-200b-3p and miRNA-200b-5p inhibits epithelial-to-mesenchymal transition in TNBC (Rhodes et al., 2015). Given that both miR-124-3p overexpression and ABCC4 knockdown inhibited cell proliferation, invasion, and migration, hindered the normal cell cycle process, and enhanced the sensitivity to ADR in MCF-7-ADR, we further investigated the effects of the combination of miR-124-3p overexpression and ABCC4 knockdown on the biologic function and sensitivity to ADR of MCF-7-ADR. As expected, what we found was that the combination of miR-124-3p overexpression and ABCC4 knockdown had a stronger promotive effect on drug sensitivity to ADR in MCF-7-ADR cells, compared with miR-124-3p overexpression and ABCC4 knockdown MCF-7-ADR cells. Moreover, miR-124-3p overexpression significantly inhibited protein expression of P-gp, a multidrug resistance-related protein in MCF-7-ADR cells. Our data demonstrated that combination of miR-124-3p overexpression and ABCC4 knockdown enhanced sensitivity to ADR in MCF-7-ADR cells.
In conclusion, we determined that both ABCC4 and miR-124-3p were aberrantly expressed in breast cancer tissue and MCF-7-ADR cells. Moreover, inhibition of ABCC4 and miR-124-3p overexpression both significantly enhanced the sensitivity to ADR in MCF-7-ADR cells, and dual-targeting miR-124-3p and ABCC4 had a stronger promotive effect on the sensitivity to ADR in MCF-7-ADR cells. Taken together, our findings confirmed that concentrating on the regulation of multigene therapy approach may provide a novel and effective treatment against resistant breast cancer.
Footnotes
Author Disclosure Statement
No competing financial interests exist.